
Dynamics of Pyricularia oryzae Population Race Structures from 2003 to 2017 in Jiangsu Province, China


Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Monoconidial Isolates
2.2. Plant Infection Assays and Disease Assessments
2.3. Race Identification
2.4. Data Analysis
3. Results
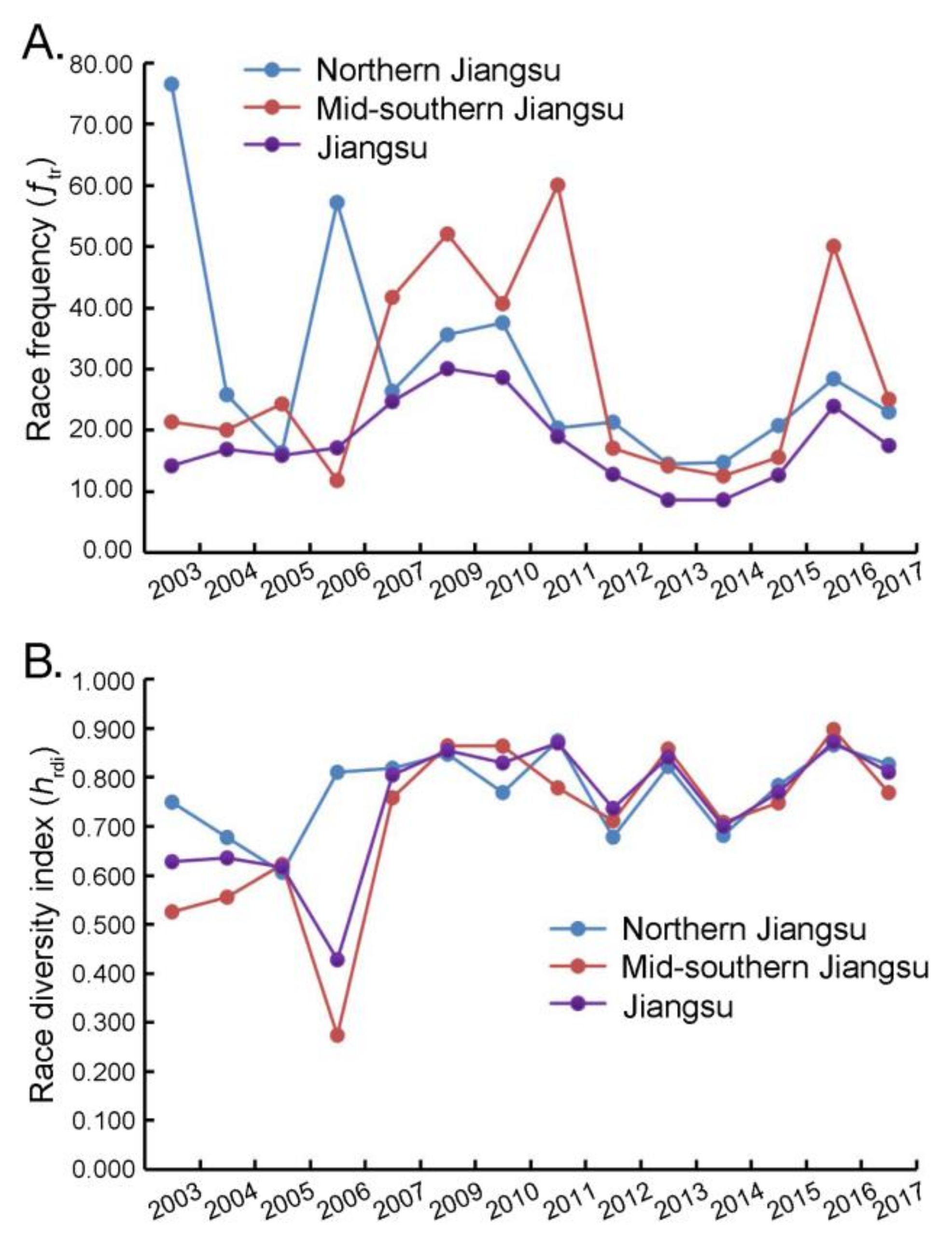
3.1. Race Diversity of P. oryzae in Jiangsu Province
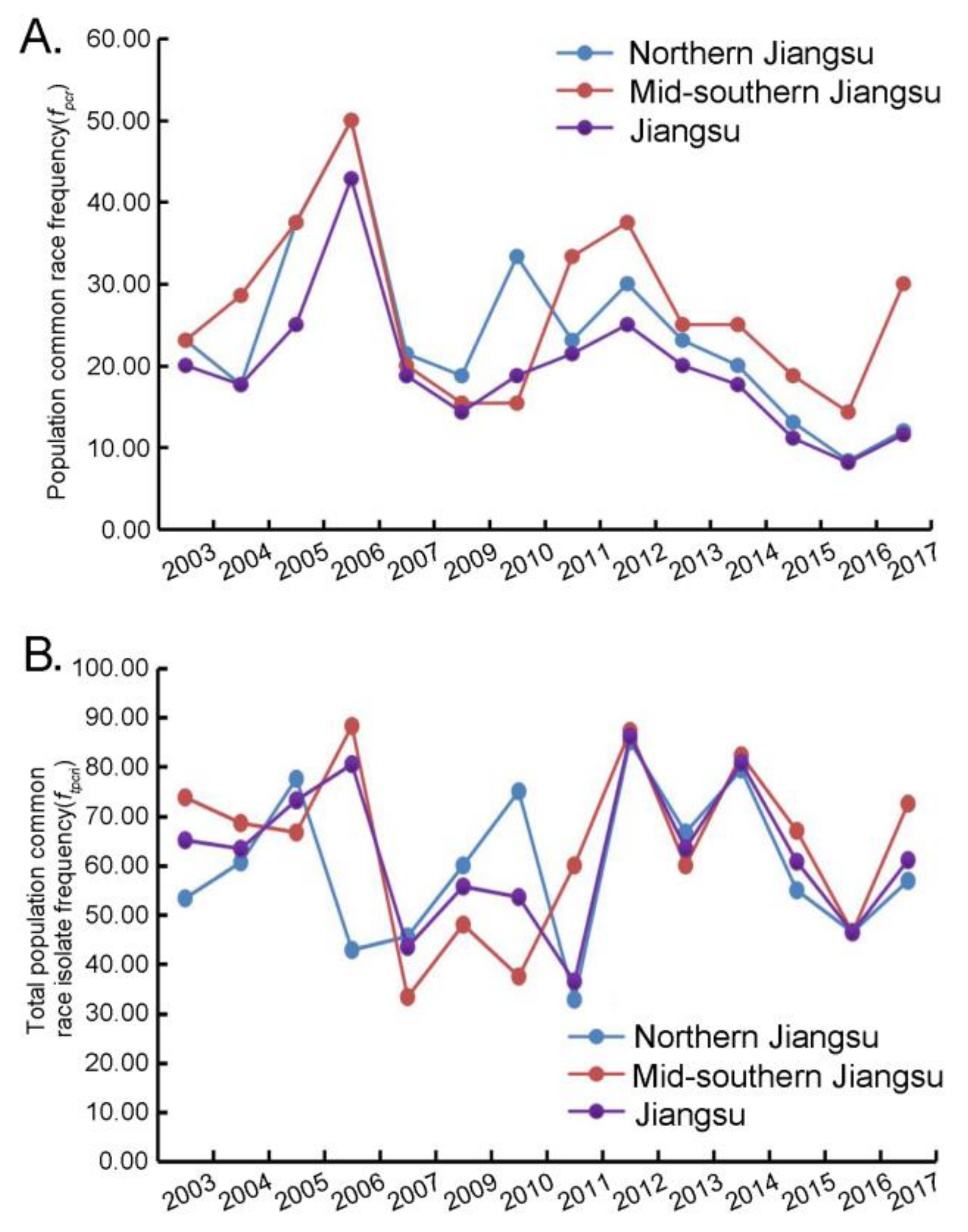
3.2. Common Race Structure in Jiangsu Province
3.3. Dominant Race Structure in Jiangsu Province
3.4. Rice Resistance Structure in Jiangsu Province
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Couch, B.C.; Kohn, L.M. A multilocus gene geneology concordant with host preference indicates segregation of a new species, Magnaporthe oryzae, from Magnaporthe grisea. Mycologia 2002, 94, 683–693. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Zhai, K.; Xie, Z.; Yang, D.; Zhu, X.; Liu, J.; Wang, X.; Qin, P.; Yang, Y.; Zhang, G.; et al. Epigenetic regulation of antagonistic receptors confers rice blast resistance with yield balance. Science 2017, 355, 962–965. [Google Scholar] [CrossRef] [PubMed]
- Singh, W.H.; Kapila, R.K.; Sharma, T.R.; Rathour, R. Genetic and physical mapping of a new allele of Pik locus from japonica rice ‘Lijiangxintuanheigu’. Euphytica 2015, 205, 889–901. [Google Scholar] [CrossRef]
- Wilson, R.A.; Talbot, N.J. Under pressure, investigating the biology of plant infection by Magnaporthe oryzae. Nat. Rev. Microbiol. 2009, 7, 185–195. [Google Scholar] [CrossRef]
- Zhai, C.; Zhang, Y.; Yao, N.; Lin, F.; Liu, Z.; Dong, Z.; Wang, L.; Pan, Q. Function and interaction of the coupled genes responsible for Pik-h encoded rice blast resistance. PLoS ONE 2014, 9, e98067. [Google Scholar] [CrossRef]
- Mekwatanakarn, P.; Kositratana, W.; Levy, M.; Zeigler, R.S. Pathotype and avirulence gene diversity of Pyricularia grisea in Yhailand as determined by rice lines near-isogenic for major resistance gene. Plant Dis. 2000, 84, 60–70. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Jia, Y.; Wamishe, Y.; Jia, M.H.; Valent, B. Dynamic changes in the rice blast population in the USA over six decades. Mol. Plant-Microbe Interac. 2017, 30, 803–812. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.; Wang, L.; Zhang, S.; Li, Z.; Zhang, Y.; Lin, F.; Pan, Q. Stepwise arms race between AvrPik and Pik alleles in the rice blast pathosystem. Mol. Plant-Microbe Interac. 2014, 27, 759–769. [Google Scholar] [CrossRef] [Green Version]
- Zeigler, R. The relationship between lineage and virulence in Pyricularia grisea in the Philippines. Phytopathology 1995, 85, 443–451. [Google Scholar] [CrossRef]
- Flor, H.H. Current status of the gene for gene concept. Annu. Rev. Phytopathol. 1971, 9, 275–296. [Google Scholar] [CrossRef]
- Silue, D.; Notteghem, J.L.; Tharreau, D. Evidence for a gene-for-gene relationship in the Oryza sativa-Magnaporthe grisea pathosystem. Phytopathology 1992, 82, 569–578. [Google Scholar] [CrossRef]
- Hu, Z.; Chen, J.; Wang, L.; Qiu, Q.; Pan, Q. Molecular genetic studies on the rice blast fungus population. II. Temporal and spatial characterization of genetic structure of the fungus population in 2000 in Guangdong province. Mycosystema 2002, 21, 203–209, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Kawasaki-Tanaka, A.; Hayashi, N.; Yanagihara, S.; Fukuta, Y. Diversity and distribution of rice blast (Pyricularia oryzae Cavara) races in Japan. Plant Dis. 2016, 100, 816–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolmer, J.A.; Hughes, M.E. Physiologic specialization of Puccinia triticina on wheat in the United States in 2016. Plant Dis. 2018, 102, 1066–1071. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Wan, A.; Liu, D.; Chen, X. Changes of races and virulence genes in Puccinia striiformis f. sp. tritici, the wheat stripe rust pathogen, in the United States from 1968 to 2009. Plant Dis. 2017, 101, 1522–1532. [Google Scholar] [CrossRef] [Green Version]
- All China Corporation of Research on Physiological Races of Pyricularia oryzae. Research on physiological races of rice blast fungus in China. Acta Phytopathol. Sin. 1980, 10, 71–82, (In Chinese with English Abstract). [Google Scholar]
- Xiao, D.; Zhang, P.; Wang, L.; Huang, S. Research progress on populations and physiological race distribution of rice blast pathogen (Magnaporthe grisea) in China. Chin. J. Rice Sci. 2013, 27, 312–320, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Liu, Y.; Chen, Z.; Hu, M.; Li, L.; Liu, Y. Distribution of Magnaporthe grisea populations and virulence of predominant races in Jiangsu province. Chin. J. Rice Sci. 2004, 18, 351–356, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Qi, Z.; Pan, X.; Du, Y.; Shen, L.; Yu, M.; Cao, H.; Song, T.; Yu, J.; Zhang, R.; Yong, M.; et al. Pathogenicity and population structure analysis of Pyricularia oryzae in different districts of Jiangsu province, China. Plant Pathol. 2021, 70, 449–458. [Google Scholar] [CrossRef]
- Qi, Z.; Yu, J.; Shen, L.; Yu, Z.; Yu, M.; Du, Y.; Zhang, R.; Song, T.; Yin, X.; Zhou, Y.; et al. Enhanced resistance to rice blast and sheath blight in rice (Oryza sativa L.) by expressing the oxalate decarboxylase protein Bacisubin from Bacillus subtilis. Plant Sci. 2017, 265, 51–60. [Google Scholar] [CrossRef]
- Pan, Q.; Wang, L.; Ikehashi, H.; Tanisaka, T. Identification of a new blast resistance gene in the indica rice cultivar kasalath using Japanese differential cultivars and isozyme markers. Phythopathology 1996, 86, 1071–1075. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, J.; Yao, Y.; Jin, X.; Correll, J.; Wang, L.; Pan, Q. Dynamics of race structures of the rice blast pathogen population in Heilongjiang province, China from 2006 through 2015. Plant Dis. 2019, 103, 2759–2763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Xiao, N.; Yu, L.; Pan, C.; Li, Y.; Zhang, X.; Liu, G.; Dai, Z.; Pan, X.; Li, A. Combination patterns of major R genes determine the level of resistance to the M. oryzae in rice (Oryza sativa L.). PLoS ONE 2015, 10, e0126130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiyosawa, S.; Ikehashi, H.; Kato, H.; Zhong, Z. Pathogenicity test of Philippine isolates of blast fungus sing two sets of rice varieties. Japan. J. Breed. 1981, 31, 367–376. [Google Scholar] [CrossRef] [Green Version]
- Tsunematsu, H.; Yanoria, M.; Ebron, L.; Nagao, H.; Ikuo, A.; Hiroshi, K.; Tokio, I.; Khush, G. Development of monogenic lines for rice blast resistance. Breed. Sci. 2000, 50, 229–234. [Google Scholar] [CrossRef] [Green Version]
- Telebanco-Yanoria, M.; Koide, Y.; Fukuta, Y.; Imbe, T.; Kato, H.; Tsunematsu, H.; Kobayashi, N. Development of near-isogenic lines of Japonica-type rice variety Lijiangxintuanheigu as differentials for blast resistance. Breed. Sci. 2010, 60, 629–638. [Google Scholar] [CrossRef] [Green Version]
- Wang, C. Current situation and development trend of rice breeding and production in Jiangsu province. Jiangsu Agr. Sci. 2005, 2, 1–6, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- McDonald, B.A.; Linde, C. Pathogen population genetics, evolutionary potential, and durable resistance. Annu. Rev. Phytopathol. 2002, 40, 349–379. [Google Scholar] [CrossRef] [Green Version]
- Saleh, D.; Milazzo, J.; Adreit, H.; Fournier, E.; Tharreau, D. South-East Asia is the center of origin, diversity and dispersion of the rice blast fungus, Magnaporthe oryzae. New Phytol. 2014, 201, 1440–1456. [Google Scholar] [CrossRef] [Green Version]
- Dong, L.; Wang, Q.; Liu, S.; Zheng, F.; Li, X.; Yang, Q. Pathogenicity analysis of Magnaporthe oryzae populations of Yunnan on monogenic lines for resistance to rice blast. Southwest China J. Agric. Sci. 2012, 25, 467–473, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Zhang, Y.; Zhu, Q.; Yao, Y.; Zhao, Z.; Correll, J.; Wang, L.; Pan, Q. The race structure of the rice blast pathogen across southern and northeastern China. Rice 2017, 10, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Parameter | 2003 A (106) | 2004 (101) | 2005 (82) | 2006 (41) | 2007 (69) | 2009 (70) | 2010 (56) | 2011 (74) | 2012 (94) | 2013 (175) | 2014 (198) | 2015 (214) | 2016 (155) | 2017 (149) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| No. of race group | 7 | 6 | 6 | 4 | 6 | 7 | 7 | 6 | 5 | 6 | 6 | 7 | 7 | 7 |
| No. of race | 15 | 17 | 13 | 7 | 16 | 21 | 16 | 14 | 12 | 15 | 17 | 27 | 37 | 26 |
| Race frequency (ƒtr) | 14.15 a–d | 16.83 b–e | 15.85 a–e | 17.07 a–f | 23.19 c–f | 30.00 f | 28.57 e,f | 18.92 b–f | 12.77 a–c | 8.57 a | 8.59 a | 12.62 a,b | 23.87 d–f | 17.45 b–e |
| Race diversity index (hrdi) | 0.627 | 0.635 | 0.616 | 0.427 | 0.803 | 0.854 | 0.829 | 0.869 | 0.736 | 0.841 | 0.699 | 0.769 | 0.871 | 0.810 |
| χ2 for ftr | 45.55 B | |||||||||||||
| Race/Parameter | ZB15 | ZC15 | ZG1 | ƒpcr | ∑/ƒtpcri | χ2 for ftpcri | |
|---|---|---|---|---|---|---|---|
| 2003 (106) A | No. | 3 | 2 | 64 | 20.00 | 65.09 b–e | 113.99 B |
| ƒpcri | 2.83 | 1.89 | 60.38 | ||||
| 2004 (101) | No. | 3 | 1 | 60 | 17.65 | 63.37 b–d | |
| ƒpcri | 2.97 | 0.99 | 59.41 | ||||
| 2005 (82) | No. | 3 | 7 | 50 | 25.00 | 73.17 c–e | |
| ƒpcri | 3.66 | 8.54 | 60.98 | ||||
| 2006 (41) | No. | 1 | 1 | 31 | 42.86 | 80.49 c–e | |
| ƒpcri | 2.44 | 2.44 | 75.61 | ||||
| 2007 (69) | No. | 3 | 5 | 22 | 18.75 | 43.48 a,b | |
| ƒpcri | 4.35 | 7.25 | 31.88 | ||||
| 2009 (70) | No. | 9 | 6 | 24 | 14.29 | 55.71 a–c | |
| ƒpcri | 12.86 | 8.57 | 34.29 | ||||
| 2010 (56) | No. | 3 | 5 | 22 | 18.75 | 53.57 a–c | |
| ƒpcri | 5.36 | 8.93 | 39.29 | ||||
| 2011 (74) | No. | 1 | 10 | 16 | 21.43 | 36.49 a | |
| ƒpcri | 1.35 | 13.51 | 21.62 | ||||
| 2012 (94) | No. | 16 | 30 | 35 | 25.00 | 86.17 e | |
| ƒpcri | 17.02 | 31.91 | 37.23 | ||||
| 2013 (175) | No. | 31 | 33 | 47 | 20.00 | 63.43 b–d | |
| ƒpcri | 17.71 | 18.86 | 26.86 | ||||
| 2014 (198) | No. | 8 | 68 | 84 | 17.65 | 80.81 d,e | |
| ƒpcri | 4.04 | 34.34 | 42.42 | ||||
| 2015 (214) | No. | 4 | 31 | 95 | 11.11 | 60.75 b,c | |
| ƒpcri | 1.87 | 14.49 | 44.39 | ||||
| 2016 (155) | No. | 2 | 20 | 50 | 8.11 | 46.45 a,b | |
| ƒpcri | 1.29 | 12.90 | 32.26 | ||||
| 2017 (149) | No. | 11 | 20 | 60 | 11.54 | 61.07 b,c | |
| ƒpcri | 7.38 | 13.42 | 40.27 | ||||
| Population | Jiangsu | Northern Jiangsu | Mid-Southern Jiangsu | |||
|---|---|---|---|---|---|---|
| Top Races (No. of Isolates) | ftri | Top Races (No. of Isolates) | ftri | Top Races (No. of Isolates) | ftri | |
| 2003 (106) A | ZG1 (64) | 60.38 b,d | ZG1 (22) A | 48.89 a,b | ZG1 (42) A | 68.85 c,d |
| 2004 (101) | ZG1 (60) | 59.41 b,d | ZG1 (37) | 56.06 b | ZG1 (23) | 65.71 b–d |
| 2005 (82) | ZG1 (50) | 60.98 b,d | ZG1 (30) | 61.22 b | ZG1 (20) | 60.61 b–d |
| 2006 (41) | ZG1 (31) | 75.61 d | ZB13 (3) | 42.86 a,b | ZG1 (29) | 85.29 d |
| 2007 (69) | ZG1 (22) | 31.88 a,b | ZG1 (18) | 31.58 a,b | ZF1 (5) | 41.67 a–c |
| 2009 (70) | ZG1 (24) | 34.29 a–c | ZG1 (15) | 33.33 a,b | ZG1 (9) | 36.00 a–c |
| 2010 (56) | ZG1 (22) | 39.29 a–c | ZG1 (11) | 45.83 a,b | ZG1 (11) | 34.38 a–c |
| 2011 (74) | ZD1 (16)/ZG1 (16) | 21.62 a | ZD1 (15) | 23.44 a | ZG1 (5) | 50.00 b–d |
| 2012 (94) | ZG1 (35) | 37.23 a–c | ZG1 (25) | 53.19 a,b | ZC15 (22) | 46.81 b,c |
| 2013 (175) | ZG1 (47) | 26.86 a | ZG1 (30) | 33.33 a,b | ZG1 (17)/ZB15 (17)/ZC15 (17) | 20.00 a |
| 2014 (198) | ZG1 (84) | 42.42 a–c | ZG1 (50) | 49.02 a,b | ZC15 (39) | 40.63 a–c |
| 2015 (214) | ZG1 (95) | 44.39 b,c | ZG1 (48) | 43.24 a,b | ZG1 (47) | 45.63 b,c |
| 2016 (155) | ZG1 (50) | 32.26 a–c | ZG1 (43) | 33.86 a,b | ZG1 (7) | 25.00 a,b |
| 2017 (149) | ZG1 (60) | 40.27 a–c | ZG1 (42) | 38.53 a,b | ZG1 (18) | 45.00 a–c |
| χ2 for ftri | 100.02 B | 38.09 B | 74.45 B | |||
| Differential Cultivar | Tetep | Zhenlong 13 | Sifeng 43 | Dongnong 363 | Kando 51 | Hejiang 18 | Lijiangxintuanheigu | |
|---|---|---|---|---|---|---|---|---|
| Race code | A64 | B32 | C16 | D8 | E4 | F2 | G1 | |
| Resistance gene | Pi1, Pi4, Pi54, Pid3, Pia, Pita, Pi5 | Pik, Pia, Pi9, Pb1, β, ε | Pib, Pia, Pi9, Pb1, α | Pik, Pia, Piz, Pish, Pi54, β | Pik, Pish, γ | Pii, Pia, Pi9, Pik, Pish, δ | Pik-l | |
| Resistance gene (ƒCR) | 2003 (106) | 99.06 * | 77.36 | 77.36 | 85.85 * | 95.28 * | 72.64 | 0.00 |
| 2004 (101) | 100.00 * | 80.20 | 84.16 | 94.06 * | 85.15 * | 76.24 | 0.00 | |
| 2005 (82) | 100.00 * | 85.37 * | 80.49 | 95.12 * | 92.68 * | 87.80 * | 0.00 | |
| 2006 (41) | 100.00 * | 87.80 * | 80.49 | 97.56 * | 95.12 * | 85.37 * | 0.00 | |
| 2007 (69) | 94.20 * | 81.16 | 73.91 | 85.51 * | 100.00 * | 46.38 | 0.00 | |
| 2009 (70) | 97.14 * | 64.29 | 64.29 | 80.00 | 85.71 * | 85.71 * | 0.00 | |
| 2010 (56) | 98.21 * | 76.79 | 73.21 | 76.79 | 60.71 | 73.21 | 0.00 | |
| 2011 (74) | 97.30 * | 93.24 * | 70.27 | 51.35 | 41.89 | 58.11 | 0.00 | |
| 2012 (94) | 100.00 * | 76.60 | 40.43 | 93.62 * | 94.68 * | 90.43 * | 0.00 | |
| 2013 (175) | 99.43 * | 53.14 | 32.57 | 78.29 | 97.71 * | 82.29 | 0.00 | |
| 2014 (198) | 97.98 * | 89.90 * | 47.47 | 91.92 * | 97.47 * | 90.91 * | 0.00 | |
| 2015 (214) | 95.79 * | 84.11 | 70.09 | 84.11 | 82.71 | 90.65 * | 0.00 | |
| 2016 (155) | 85.81 * | 81.29 | 58.06 | 76.13 | 65.16 | 76.77 | 0.00 | |
| 2017 (149) | 95.97 * | 80.54 | 55.70 | 83.22 | 83.22 | 80.54 | 0.00 | |
| Total resistance frequencies (ƒTCR) | 96.72 | 78.98 | 61.05 | 83.90 | 85.29 | 80.74 | 0.00 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qi, Z.; Du, Y.; Liu, M.; Yu, J.; Zhang, R.; Yu, M.; Cao, H.; Song, T.; Pan, X.; Liang, D.; et al. Dynamics of Pyricularia oryzae Population Race Structures from 2003 to 2017 in Jiangsu Province, China. Agronomy 2022, 12, 956. https://doi.org/10.3390/agronomy12040956
Qi Z, Du Y, Liu M, Yu J, Zhang R, Yu M, Cao H, Song T, Pan X, Liang D, et al. Dynamics of Pyricularia oryzae Population Race Structures from 2003 to 2017 in Jiangsu Province, China. Agronomy. 2022; 12(4):956. https://doi.org/10.3390/agronomy12040956
Chicago/Turabian StyleQi, Zhongqiang, Yan Du, Muxing Liu, Junjie Yu, Rongsheng Zhang, Mingna Yu, Huijuan Cao, Tianqiao Song, Xiayan Pan, Dong Liang, and et al. 2022. "Dynamics of Pyricularia oryzae Population Race Structures from 2003 to 2017 in Jiangsu Province, China" Agronomy 12, no. 4: 956. https://doi.org/10.3390/agronomy12040956