
Identification of Aluminum Tolerance in Ethiopian Chickpea (Cicer arietinum L.) Germplasm


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experiment I: Determination of Optimum Aluminum Concentration
2.1.1. Plant Materials
2.1.2. Sterilization and Germination
2.1.3. Nutrient Solution Preparation and Growth Conditions
2.1.4. Experimental Layout and Statistical Analysis
2.2. Experiment II: Evaluation of Improved Cultivars and Advanced Chickpea Genotypes for Tolerance to Aluminum
2.2.1. Plant Materials
2.2.2. Experimental Design and Statistical Analysis
3. Results
3.1. Determination of Optimum Al Concentration
3.1.1. Analysis of Variance (ANOVA) for Seedling Traits
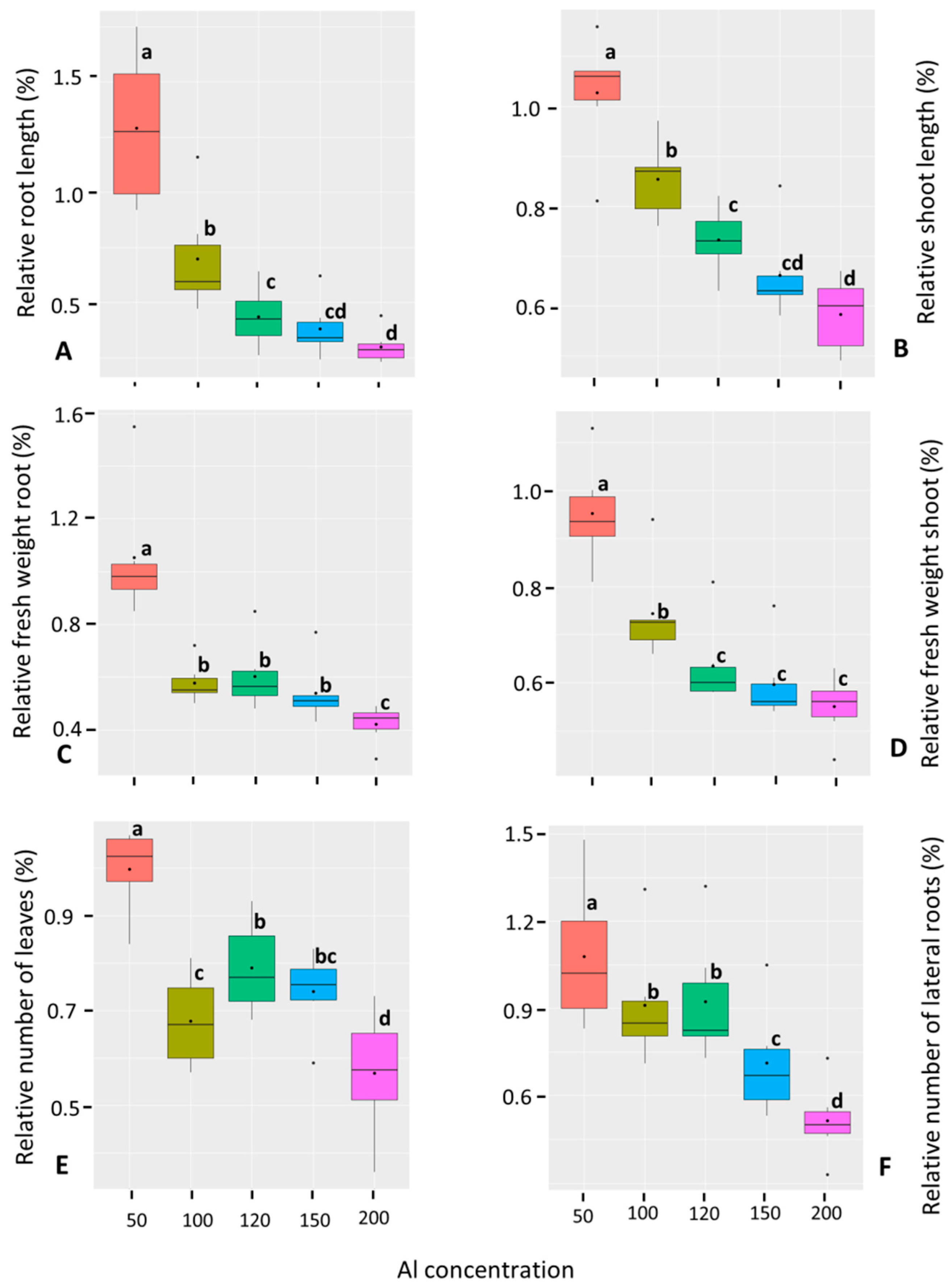
3.1.2. Response of Chickpea Genotypes to Increasing Al Concentrations
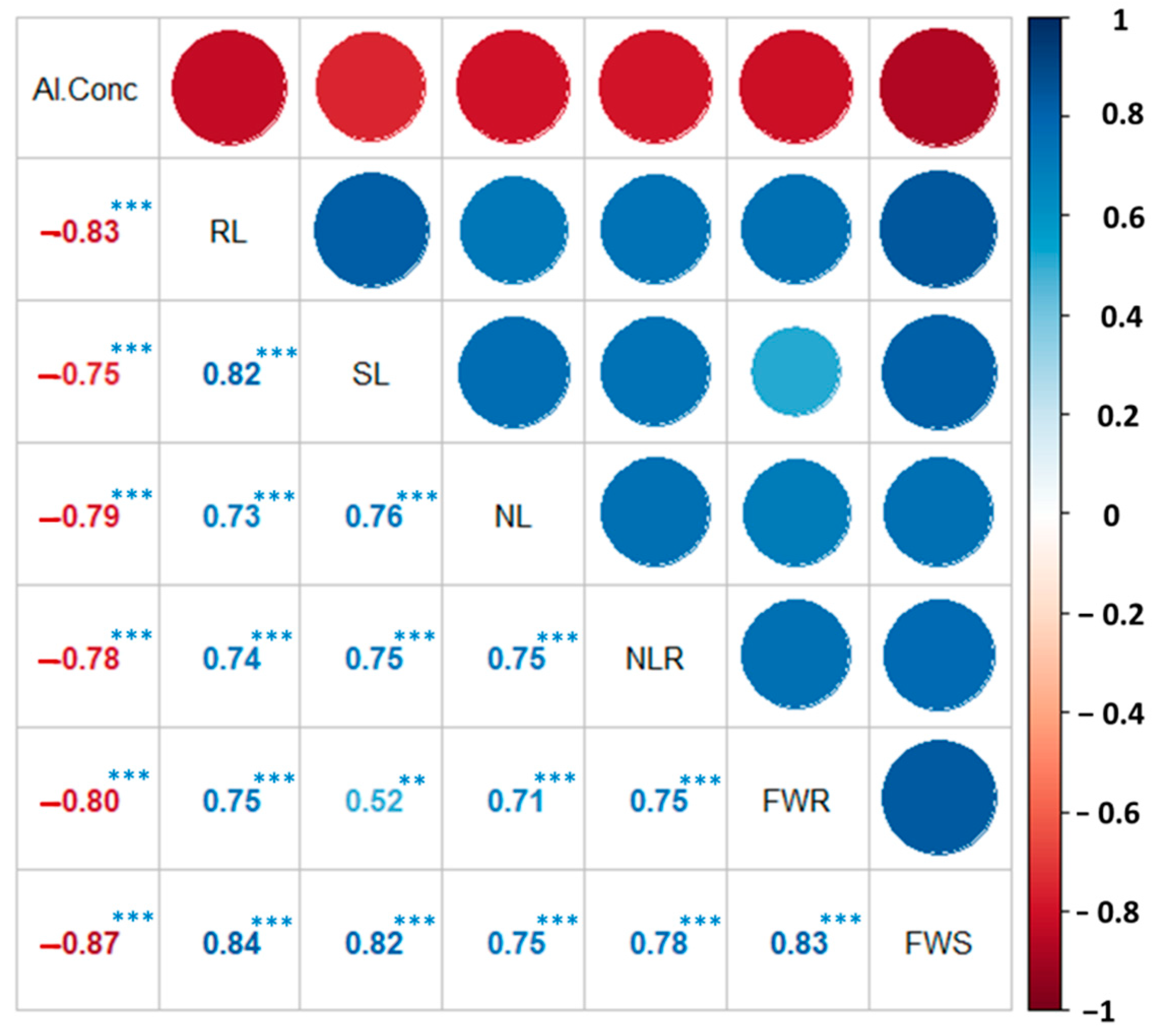
3.1.3. Correlation Analysis among Phenotypic Traits of Chickpea Genotypes
3.2. Screening of Improved Cultivars and Advanced Genotypes of Chickpea for Aluminum Tolerance
3.2.1. Analysis of Variance (ANOVA) for Seedling Phenotypic Traits
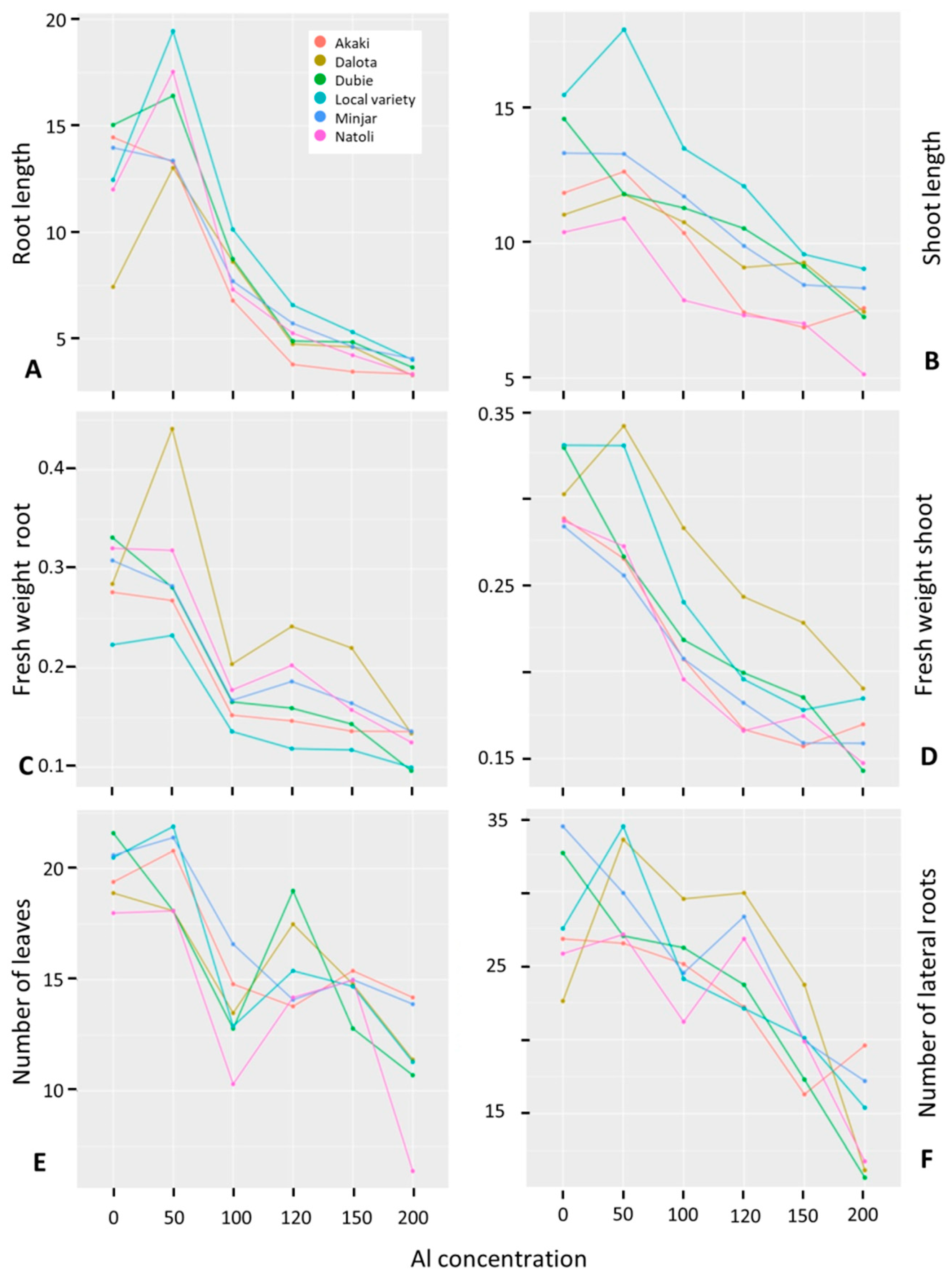
3.2.2. Differential Response of Ethiopian Chickpea Germplasm to Al Treatment
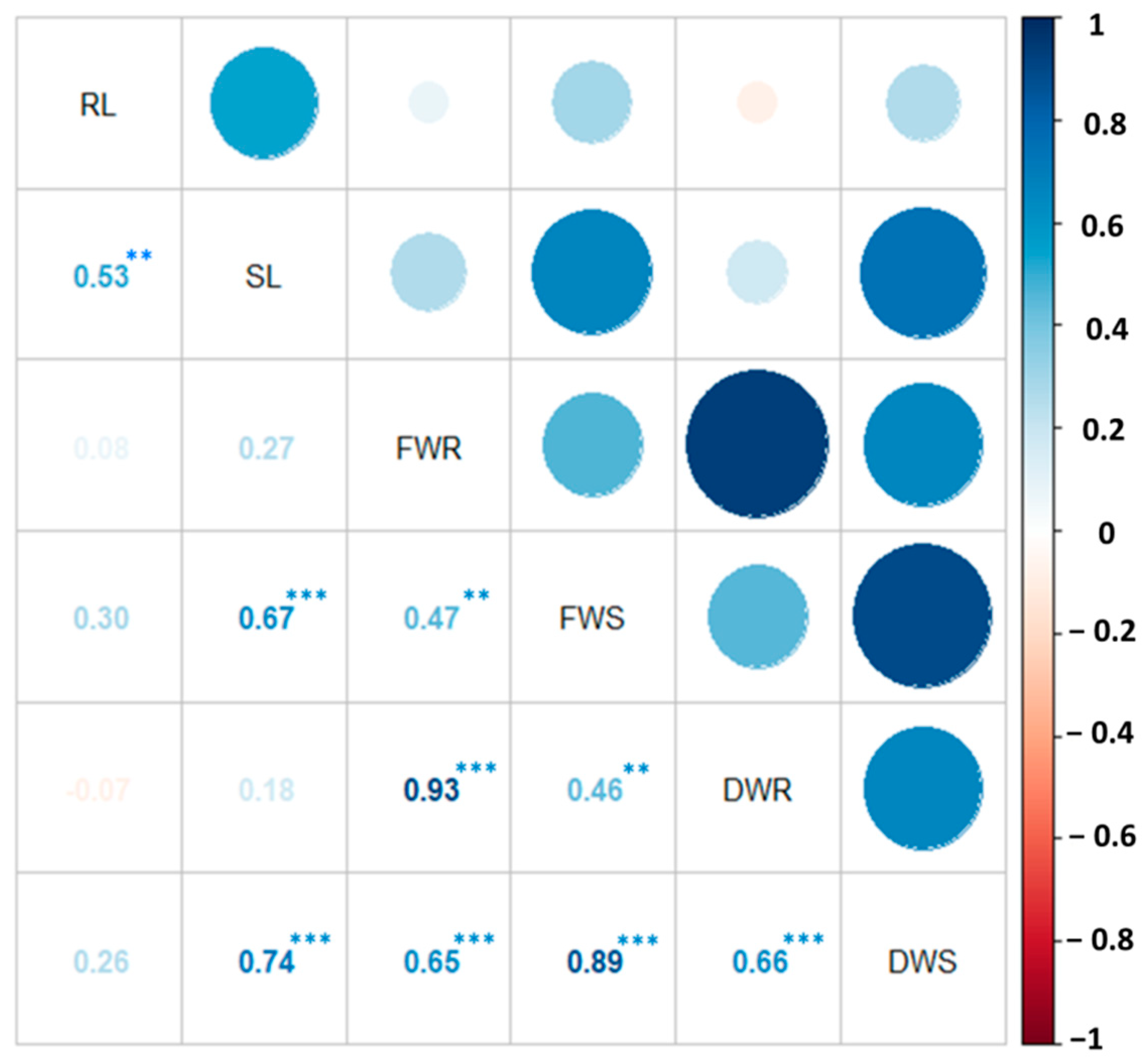
3.2.3. Relationship among Phenotypic Traits of Improved and Advanced Chickpea Genotypes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Damte, T.; Ojiewo, C.O. Current status of wilt/root rot diseases in major chickpea growing areas of Ethiopia. Arch. Phytopathol. Plant Prot. 2016, 49, 222–238. [Google Scholar] [CrossRef]
- FAOSTAT. FAO (Food and Agriculture Organization of the United Nations). Available online: https://www.fao.org/faostat/en/ (accessed on 18 June 2020).
- CSA (Central Statistical Agency). The Federal Democratic Republic of Ethiopia Central Statistical Agency Report on Area and Production of Major Crops Private Peasant Holdings; Meher Season: Addis Ababa, Ethiopia, 2018; pp. 1–58. [Google Scholar]
- Dadi, L.; Regassa, S.; Fikre, A.; Mitiku, D.; Gaur, P.; Gowda, C.; Bantilan, M. Adoption Studies on Improved Chickpea Varieties in Ethiopia; International Crops Research Institute for the Semi-Arid Tropics (ICRISAT): Patancheru, India, 2005; pp. 1–32. [Google Scholar]
- Shiferaw, B.; Teklewold, H. Structure and Functioning of Chickpea Markets in Ethiopia: Evidence Based on Analyses of Value Chains Linking Smallholders and Markets; Improving Productivity and Market Success (IPMS) of Ethiopian Farmers Project Working Paper 6; ILRI (International Livestock Research Institute): Nairobi, Kenya, 2007; pp. 1–63. [Google Scholar]
- Ferede, S.; Fikre, A.; Ahmed, S. Assessing the Competitiveness of Smallholders Chickpea Production in the Central Highlands of Ethiopia. Ethiop. J. Crop Sci. 2018, 6, 51–65. [Google Scholar]
- Korbu, L.; Tafes, B.; Kassa, G.; Mola, T.; Fikre, A. Unlocking the genetic potential of chickpea through improved crop management practices in Ethiopia. A review. Agron. Sustain. Dev. 2020, 40, 13. [Google Scholar] [CrossRef]
- Ferede, S.; Aliy, S.; Girma, N. Validating Product Concepts for Optimizing Chickpea Breeding; Ethiopian Institute of Agricultural Research (EIAR): Addis Ababa, Ethiopia, 2019; pp. 1–55.
- USDA (United States Department of Agriculture Foreign Agricultural Service). Pulse Crops Market Update, Assessments of Commodity and Trade Issues; Report Number: ET1819; Global Agricultural Information Network (GAIN): Addis Ababa, Ethiopia, 2018.
- Fikre, A.; Funga, A.; Korbu, L.; Eshete, M.; Girma, N. Stability Analysis in Chickpea Genotype Sets as Tool for Breeding Germplasm Structuring Strategy and Adaptability Scoping. Ethiop. J. Crop Sci. 2018, 6, 19–37. [Google Scholar]
- Hawando, T. Soil and Climatic Stresses on Chickpea Production in Ethiopia. In Adaptation of Chickpea and Pigeonpea to Abiotic Stresses, Proceedings of the Consultants’ Workshop, ICRISAT Center, Patancheru, India, 19–21 December 1984; Gopalachari, N.C., Garver, C.L., Eds.; International Crops Research Institute for the Semi-Arid Tropics (ICRISAT): Patancheru, India, 1987; pp. 159–167. [Google Scholar]
- Alemu, B.; Lule, D. Yield and agronomic performances of desi type chickpea genotypes against acidic soil of Western Ethiopia. J. Agric. Biotechnol. Sustain. Dev. 2018, 10, 116–121. [Google Scholar] [CrossRef] [Green Version]
- Delhaize, E.; Ryan, P.R. Aluminum toxicity and tolerance in plants. Plant Physiol. 1995, 107, 315–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kochian, L.V.; Piñeros, M.A.; Hoekenga, O.A. The physiology, genetics and molecular biology of plant aluminum resistance and toxicity. Plant Soil 2005, 274, 175–195. [Google Scholar] [CrossRef]
- Roy, B.; Bhadra, S. Effects of toxic levels of aluminium on seedling parameters of rice under hydroponic culture. Rice Sci. 2014, 21, 217–223. [Google Scholar] [CrossRef]
- Delhaize, E.; Ryan, P.R.; Randall, P.J. Aluminum Tolerance in Wheat (Triticum aestivum L.) II. Aluminum-Stimulated Excretion of Malic Acid from Root Apices. Plant Physiol. 1993, 695–702. [Google Scholar] [CrossRef] [Green Version]
- Kochian, L.V. Cellular mechanisms of aluminum toxicity and resistance in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1995, 46, 237–260. [Google Scholar] [CrossRef]
- Hede, A.R.; Skovmand, B.; López-Cesati, J. Acid Soils and Aluminum Toxicity. In Application of Physiology in Wheat Breeding; Reynolds, M.P., Ortiz-Monasterio, J.I., McNab, A., Eds.; International Maize and Wheat Improvement Center (CIMMYT): Texcoco, Mexico, 2001; pp. 172–182. [Google Scholar]
- Blancaflor, E.B.; Jones, D.L.; Gilroy, S. Alterations in the Cytoskeleton Accompany Aluminum Induced Growth Inhibition and Morphological Changes in Primary Roots of Maize. Plant Physiol. 1998, 118, 159–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Legesse, H.; Nigussie-Dechassa, R.; Gebeyehu, S.; Bultosa, G.; Mekbib, F. Response to Soil Acidity of Common Bean Genotypes (Phaseolus vulgaris L.) under Field Conditions at Nedjo, Western Ethiopia Hirpa. Sci. Technol. Arts Res. J. 2013, 2, 3–15. [Google Scholar] [CrossRef] [Green Version]
- Kassahun, B. Soil Fertility Mapping and Fertilizer Blending; Ethiopian Agricultural Transformation Agency (Ethiopian ATA): Addis Ababa, Ethiopia, 2015.
- Mosissa, F. Progress of Soil Acidity Management Research in Ethiopia. Adv. Crop Sci. Technol. 2018, 6, 1–8. [Google Scholar] [CrossRef]
- Kassie, M.; Shiferaw, B.; Asfaw, S.; Abate, T.; Muricho, G.; Ferede, S.; Eshete, M.; Assefa, K. Current Situation and Future Outlooks of the Chickpea Sub—Sector in Ethiopia; International Crops Research Institute for the Semi-Arid Tropics (ICRISAT): Nairobi, Kenya, 2009; pp. 1–39. [Google Scholar]
- Agegnehu, G.; Yirga, C.; Erkossa, T. Soil Acidity Management; Ethiopian Institute of Agricultural Research (EIAR): Addis Ababa, Ethiopia, 2019; pp. 1–56.
- Fageria, N.K.; Wright, R.J.; Baligar, V.C. Rice cultivar response to aluminum in nutrient solution. Commun. Soil Sci. Plant Anal. 1988, 19, 1133–1142. [Google Scholar] [CrossRef]
- Ma, J.F.; Zheng, S.J.; Li, X.F.; Takeda, K.; Matsumoto, H. A rapid hydroponic screening for aluminium tolerance in barley. Plant Soil 1997, 191, 133–137. [Google Scholar] [CrossRef]
- Rao, I.M.; Zeigler, R.S.; Vera, R.; Sarkarung, S. Selection and Breeding for Acid-Soil Tolerance in Crops Upland rice and tropical forages as case studies. Bioscience 1993, 43, 454–465. [Google Scholar] [CrossRef]
- Furlani, P.R.; Clark, R.B. Screening Sorghum for Aluminum Tolerance in Nutrient Solutions 1. Agron. J. 1981, 73, 587–594. [Google Scholar] [CrossRef]
- Choudhary, A.K.; Singh, D. Screening of pigeonpea genotypes for nutrient uptake efficiency under aluminium toxicity. Physiol. Mol. Biol. Plants 2011, 17, 145–152. [Google Scholar] [CrossRef] [Green Version]
- Hede, A.R.; Skovmand, B.; Ribaut, J.M.; González-De-León, D.; Stølen, O. Evaluation of aluminium tolerance in a spring rye collection by hydroponic screening. Plant Breed. 2002, 121, 241–248. [Google Scholar] [CrossRef]
- Dos Santos Neto, J.; Delfini, J.; Silva, T.W.; Hirose, A.A.; Novais, J.M.; Azeredo Gonçalves, L.S.; Moda-Cirino, V. Response of common bean cultivars and lines to aluminum toxicity. Agronomy 2020, 10, 296. [Google Scholar] [CrossRef] [Green Version]
- Shavrukov, Y.; Genc, Y.; Hayes, J. The Use of Hydroponics in Abiotic Stress Tolerance Research. In Hydroponics—A Standard Methodology for Plant Biological Researches; Asao, T., Ed.; InTech: Rijeka, Croatia, 2012; pp. 39–66. [Google Scholar]
- Giaveno, C.D.; Miranda Filho, J.B. Rapid screening for aluminum tolerance in maize (Zea mays L.). Genet. Mol. Biol. 2000, 23, 847–850. [Google Scholar] [CrossRef] [Green Version]
- Foy, C.D. Tolerance of durum wheat lines to an acid, aluminum-toxic subsoil. J. Plant Nutr. 1996, 19, 1381–1394. [Google Scholar] [CrossRef]
- Wayima, E.F.; Ligaba-Osena, A.; Dagne, K.; Tesfaye, K.; Machuka, E.M.; Mutiga, S.K.; Delhaize, E. Screening of diverse Ethiopian durum wheat accessions for aluminum tolerance. Agronomy 2019, 9, 440. [Google Scholar] [CrossRef] [Green Version]
- Echart, C.L.; Fernandes Barbosa-Neto, J.; Garvin, D.F.; Cavalli-Molina, S. Aluminum tolerance in barley: Methods for screening and genetic analysis. Euphytica 2002, 126, 309–313. [Google Scholar] [CrossRef]
- Abate, E.; Hussein, S.; Laing, M.; Mengistu, F. Quantitative responses of tef [Eragrostis tef ( Zucc.) Trotter] and weeping love grass [Eragrostis curvula (Schrad.) Nees] varieties to acid soil. Aust. J. Crop Sci. 2013, 7, 1854–1860. [Google Scholar]
- Campbell, K.A.G.; Carter, T.E. Aluminum Tolerance in Soybean: I. Genotypic Correlation and Repeatability of Solution Culture and Greenhouse Screening Methods. Crop Sci. 1990, 30, 1049. [Google Scholar] [CrossRef]
- Villagarcia, M.R.; Carter, T.E.; Rufty, T.W.; Niewoehner, A.S.; Jennette, M.W.; Arrellano, C. Genotypic rankings for aluminum tolerance of soybean roots grown in hydroponics and sand culture. Crop Sci. 2001, 41, 1499–1507. [Google Scholar] [CrossRef] [Green Version]
- Belachew, K.Y.; Stoddard, F.L. Screening of faba bean (Vicia faba L.) accessions to acidity and aluminium stresses. PeerJ 2017, 5, e2963. [Google Scholar] [CrossRef] [Green Version]
- Blair, M.W.; López-Marín, H.D.; Rao, I.M. Identification of aluminum resistant Andean common bean (Phaseolus vulgaris L.) genotypes. Braz. J. Plant Physiol. 2009, 21, 291–300. [Google Scholar] [CrossRef]
- Singh, D.; Dikshit, H.K.; Singh, R. Variation of aluminium tolerance in lentil (Lens culinaris Medik.). Plant Breed. 2012, 131, 751–761. [Google Scholar] [CrossRef]
- Froese, P.S.; Carter, A.H.; Pumphrey, M.O. Recommended Crop Species and Wheat Varieties for Acidic Soil; Washington State University: Washington, DC, USA, 2015. [Google Scholar]
- Singh, D.; Raje, R.S. Genetics of aluminium tolerance in chickpea (Cicer arietinum). Plant Breed. 2011, 130, 563–568. [Google Scholar] [CrossRef]
- Rai, R. Effects of soil acidity factors on interaction of chickpea (Cicer arietinum L.) genotypes and Rhizobium strains: Symbiotic N-fixation, grain quality and grain yield in acid soils. In Plant-Soil Interactions at Low pH, Proceedings of the Second International Symposium on Plant-Soil Interactions at Low pH, Beckley West Virginia, USA, 24–29 June 1990; Wright, R.J., Baligar, V.C., Murrmann, R.P., Eds.; Springer: Dordrecht, The Netherlands, 1991; pp. 619–631. [Google Scholar]
- Rodrigues, C.S.; Laranjo, M.; Oliveira, S. Effect of Heat and pH Stress in the Growth of Chickpea Mesorhizobia. Curr. Microbiol. 2006, 53, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Vargas, A.A.T.; Graham, P.H. Cultivar and pH effects on competition for nodule sites between isolates of Rhizobium in beans. Plant Soil 1989, 117, 195–200. [Google Scholar] [CrossRef]
- Vance, W.; Pradeep, K.; Strachan, S.R.; Diffey, S.; Bell, R.W. Novel Sources of Tolerance to Aluminium Toxicity in Wild Cicer (Cicer reticulatum and Cicer echinospermum) Collections. Front. Plant Sci. 2021, 12, 678211. [Google Scholar] [CrossRef] [PubMed]
- Simon, L.; Smalley, T.J.; Benton Jones, J., Jr.; Lasseigne, F.T. Aluminum toxicity in tomato. Part 1. growth and mineral nutrition. J. Plant Nutr. 1994, 17, 293–306. [Google Scholar] [CrossRef]
- Delhaize, E.; Ryan, P.R.; Hebb, D.M.; Yamamoto, Y.; Sasaki, T.; Matsumoto, H. Engineering high-level aluminum tolerance in barley with the ALMT1 gene. Proc. Natl. Acad. Sci. USA 2004, 101, 15249–15254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinheiro, J.; Bates, D.; DebRoy, S.S.; Sarkar, D.; R Core Development Team. Nlme: Linear and Nonlinear Mixed Effects Models. R Package Version 3.1-155. 2022. Available online: https://CRAN.R-project.org/package=nlme (accessed on 13 December 2021).
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 48. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. 2021. Available online: https://www.R-project.org/ (accessed on 20 September 2021).
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis. Springer-Verlag New York. 2016. Available online: https://ggplot2.tidyverse.org (accessed on 20 September 2021).
- Wickham, H.; François, R.; Henry, L.; Müller, K. Dplyr: A Grammar of Data Manipulation. R Package Version 0.7.6. 2018. Available online: https://CRAN.R-project.org/package=dplyr (accessed on 13 December 2021).
- Wei, T.; Simko, V. R Package “Corrplot”: Visualization of a Correlation Matrix (Version 0.92). 2021. Available online: https://github.com/taiyun/corrplot (accessed on 20 September 2021).
- Baier, A.C.; Somers, D.J.; Gusiafson, J.P. Aluminium tolerance in wheat: Correlating hydroponic evaluations with field and soil performances. Plant Breed. 1995, 114, 291–296. [Google Scholar] [CrossRef]
- Choudhury, S.; Sharma, P. Aluminum stress inhibits root growth and alters physiological and metabolic responses in chickpea (Cicer arietinum L.). Plant Physiol. Biochem. 2014, 85, 63–70. [Google Scholar] [CrossRef]
- Fageria, N.K.; Ballgar, V.C.; Wright, R.J. Aluminum toxicity in crop plants. J. Plant Nutr. 1988, 11, 37–41. [Google Scholar] [CrossRef]
- Paul, K.I.; Black, A.S.; Conyers, M. Development of Acidic Subsurface Layers of Soil under Various Management Systems. Adv. Agron. 2003, 78, 187–214. [Google Scholar]
- Foy, C.D.; Chaney, R.L.; White, M.C. The Physiology of Metal Toxicity in Plants. Annu. Rev. Plant Physiol. 1978, 29, 511–566. [Google Scholar] [CrossRef]
- Kushwaha, J.K.; Pandey, A.K.; Dubey, R.K.; Singh, V.; Mailappa, A.S.; Singh, S. Screening of cowpea [Vigna unguiculata (L.) walp.] for aluminium tolerance in relation to growth, yield and related traits. Legum. Res. 2017, 40, 434–438. [Google Scholar] [CrossRef] [Green Version]
- Belachew, K.Y. Root and Shoot Traits Associated with Acidity and Drought Tolerance in Vicia faba L. Plants. Ph.D. Thesis, University of Helsinki, Helsinki, Finland, 25 October 2019. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | Type | Origin | Classification | Year of Release | Seed Source |
|---|---|---|---|---|---|
| Akaki | Desi | ICRISAT | Variety | 1995 | DZARC |
| Dalota | Desi | ICRISAT | Variety | 2013 | DZARC |
| Dubie | Desi | Ethiopia | Variety | 1978 | DZARC |
| Minjar | Desi | ICRISAT | Variety | 2010 | DZARC |
| Natoli | Desi | ICRISAT | Variety | 2007 | DZARC |
| Wollega LV | Desi | Ethiopia | Landrace | Unknown | FARMERS |
| Mean Square | |||||||
|---|---|---|---|---|---|---|---|
| Source of Variation | df | RL | SL | FWR | FWS | NLS | NLRS |
| Genotypes | 5 | 11.17 *** | 31.67 *** | 0.013 * | 0.007 * | 15.170 ** | 24.00 NS |
| Replication (Block) | 1 | 0.10 | 2.86 | 0.005 | 0.000 | 34.376 | 44.10 |
| Main plot error | 5 | 0.84 | 0.86 | 0.003 | 0.001 | 1.028 | 13.35 |
| Al. Conc. | 5 | 279.39 *** | 64.46 *** | 0.069 *** | 0.040 *** | 140.712 *** | 403.29 *** |
| Genotype: Al. Conc. | 25 | 4.87 *** | 1.86 NS | 0.002 * | 0.001 NS | 6.085 NS | 23.18 *** |
| Subplot error | 30 | 1.33 | 3.062 | 0.001 | 0.001 | 7.569 | 5.85 |
| CV(A) | 11.1% | 9% | 25.2% | 14.9% | 6.4% | 15.4% | |
| CV(B) | 14% | 16.9% | 13.3% | 10.9% | 17.4% | 10.2% | |
| R2 | 0.98 | 0.85 | 0.96 | 0.93 | 0.81 | 0.94 | |
| Relative Root Length | ||||||
|---|---|---|---|---|---|---|
| Al Concentration | ||||||
| Variety | 0 | 50 | 100 | 120 | 150 | 200 |
| Akaki | - | 0.92 b | 0.47 c | 0.26 e | 0.24 d | 0.23 b |
| Dalota | - | 1.77 a | 1.16 a | 0.64 a | 0.62 a | 0.44 a |
| Dubie | - | 1.09 b | 0.58 bc | 0.33 de | 0.32 cd | 0.24 b |
| Wollega LV | - | 1.56 ab | 0.81 b | 0.53 b | 0.43 b | 0.32 ab |
| Minjar | - | 0.96 b | 0.55 bc | 0.41 cd | 0.33 cd | 0.29 ab |
| Natoli | - | 1.46 ab | 0.61 bc | 0.44 bc | 0.35 bc | 0.28 ab |
| Range | - | 0.90–2.15 | 0.32–1.24 | 0.24–0.67 | 0.23–0.67 | 0.21–0.54 |
| G.M | - | 1.29 | 0.70 | 0.44 | 0.38 | 0.30 |
| SEM | - | 0.11 | 0.08 | 0.04 | 0.04 | 0.03 |
| LSD | - | 0.65 | 0.32 | 0.10 | 0.09 | 0.18 |
| CV % | - | 19.68 | 17.79 | 8.58 | 9.31 | 22.39 |
| Mean Square | ||||||||
|---|---|---|---|---|---|---|---|---|
| Source of Variation | df | RL | SL | FWR | FWS | DWR | DWS | |
| Control | Replication | 1 | 0.187 | 1.750 | 0.001 | 0.001 | 0.000 | 0.001 |
| (0 µM Al) | Genotype | 30 | 24.643 *** | 18.865 *** | 0.015 *** | 0.008 *** | 0.002 *** | 0.001 *** |
| Error | 30 | 0.572 | 0.832 | 0.002 | 0.001 | 0.0003 | 0.0003 | |
| CV | 5.54 | 9.92 | 11.48 | 11.66 | 15.04 | 16.70 | ||
| Al treatment | Replication | 1 | 0.525 | 0.817 | 0.0001 | 0.0001 | 0.0001 | 0.0002 |
| (120 µM Al) | Genotype | 30 | 2.565 *** | 16.790 *** | 0.007 *** | 0.0061 *** | 0.0012 *** | 0.0011 *** |
| Error | 30 | 0.424 | 0.313 | 0.001 | 0.0004 | 0.0001 | 0.0001 | |
| CV | 13.83 | 7.88 | 11.58 | 10.18 | 15.18 | 11.76 | ||
| Combined | Genotype | 30 | 16.67 *** | 33.71 *** | 0.016 *** | 0.013 *** | 0.002 *** | 0.002 *** |
| Al. Conc. | 1 | 2481.89 *** | 136.64 *** | 0.766 *** | 0.081 *** | 0.053 *** | 0.013 *** | |
| Replication | 1 | 0.04 | 2.48 * | 0.001 | 0.0005 | 0.0001 | 0.001 * | |
| Genotype: Al. Conc. | 30 | 10.54 *** | 1.94 *** | 0.005 *** | 0.001 ** | 0.0005 ** | 0.0002 NS | |
| Error | 61 | 0.50 | 0.56 | 0.001 | 0.001 | 0.0002 | 0.0002 | |
| CV | 7.71 | 9.22 | 11.89 | 11.12 | 15.32 | 15.01 | ||
| Genotypes | Root Length | Tolerance Index | |
|---|---|---|---|
| Control (pH 4.5 and 0 µM) | Al Treatment (pH 4.5 and 120 µM) | RTI (%) | |
| DZ-2012-CK-0032 | 15.79 c–h | 3.87 cd | 25 d–f |
| DZ-2012-CK-0034 | 15.03 d–j | 5.05 cd | 34 d–f |
| DZ-2012-CK-20113-2-0042 | 18.13 bc | 5.89 bc | 33 d–f |
| DZ-2012-CK-0233 | 13.44 g–o | 5.58 bcd | 42 b–f |
| DZ-2012-CK-0237 | 20.02 ab | 8.3 a | 42 b–f |
| DZ-2012-CK-0313 | 12.47 i–p | 4.72 cd | 38 c–f |
| Akaki | 16.19 c–g | 3.48 d | 22 f |
| Arerti | 15.06 d–i | 3.96 cd | 26 d–f |
| Chefe | 9.78 pq | 3.5 d | 37 c–f |
| Dalota | 7.31 r | 4.94 cd | 68 a |
| Dhera | 11.99 k–p | 4.64 cd | 39 c–f |
| Dimtu | 15.95 c–g | 5.19 cd | 33 d–f |
| Dubie | 14.83 d–j | 5.24 bcd | 35 d–f |
| Dz-10-11 | 11.45 m–p | 3.92 cd | 34 d–f |
| Dz-10-4 | 14.17 e–l | 4.83 cd | 35 d–f |
| Ejere | 10.65 n–q | 3.99 cd | 38 c–f |
| Fetenech | 22.19 a | 5.07 cd | 23 f |
| Habru | 14.87 d–j | 4.33 cd | 30 d–f |
| Hora | 10.44 o–q | 4.49 cd | 43 b–e |
| Kasech | 12.02 j–p | 4.07 cd | 34 d–f |
| Kobo | 10.79 n–p | 4.81 cd | 45 b–d |
| Mariye | 16.52 cde | 4.01 cd | 24 ef |
| Mastewal | 17.19 b–d | 4.23 cd | 25 ef |
| Minjar | 13.79 f–m | 5.53 bcd | 40 c–f |
| Natoli | 12.92 h–o | 5.49 bcd | 43 b–f |
| Shasho | 11.58 l–p | 3.63 cd | 32 d–f |
| Teji | 6.68 r | 4.03 cd | 60 ab |
| Teketay | 14.74 d–k | 4.11 cd | 28 d–f |
| Wollega LV | 13.44 g–n | 7.82 ab | 58 a–c |
| Worku | 16.33 c–f | 4.30 cd | 26 d–f |
| Yelebe | 7.67 qr | 3.03 d | 40 c–f |
| Range | 6.68–22.19 | 3.03–8.30 | 22–68 |
| G.M | 13.66 | 4.71 | 36 |
| SEM | 0.45 | 0.15 | 0.01 |
| CV% | 5.54 | 13.83 | 14.88 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Negusse, H.; Cook, D.R.; Haileselassie, T.; Tesfaye, K. Identification of Aluminum Tolerance in Ethiopian Chickpea (Cicer arietinum L.) Germplasm. Agronomy 2022, 12, 948. https://doi.org/10.3390/agronomy12040948
Negusse H, Cook DR, Haileselassie T, Tesfaye K. Identification of Aluminum Tolerance in Ethiopian Chickpea (Cicer arietinum L.) Germplasm. Agronomy. 2022; 12(4):948. https://doi.org/10.3390/agronomy12040948
Chicago/Turabian StyleNegusse, Hawi, Douglas R. Cook, Teklehaimanot Haileselassie, and Kassahun Tesfaye. 2022. "Identification of Aluminum Tolerance in Ethiopian Chickpea (Cicer arietinum L.) Germplasm" Agronomy 12, no. 4: 948. https://doi.org/10.3390/agronomy12040948