
Effect of Organic and Conventional Cereal Production Methods on Fusarium Head Blight and Mycotoxin Contamination Levels



Abstract
:1. Introduction
2. Materials and Methods
2.1. Literature Search and Review
2.2. Estimates of Mean Mycotoxin Concentration
3. Results and Discussion
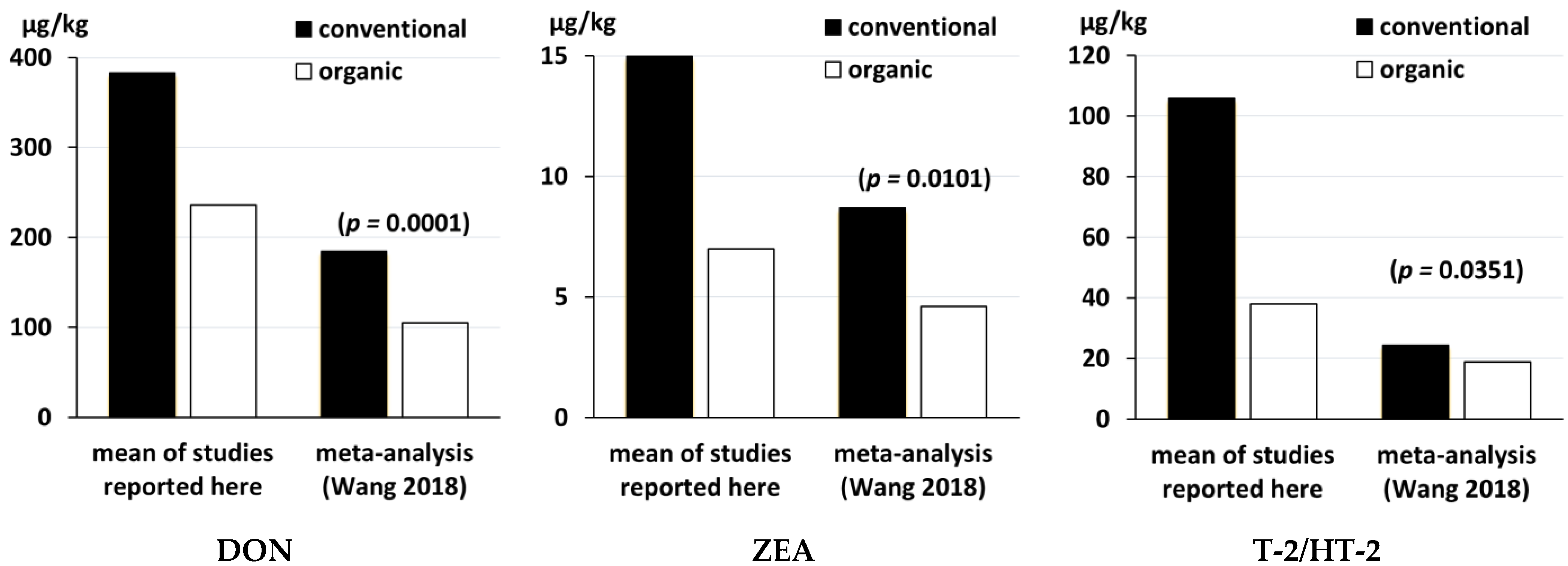
3.1. Studies That Compared Mycotoxin Levels in Organic and Conventional Cereal Grains
3.1.1. Field Experiments
3.1.2. Farm Surveys
3.2. Studies That Investigated Effects of Specific Agronomic Practices on Mycotoxin Levels
3.2.1. Crop Rotation
3.2.2. Mineral Nitrogen Fertiliser
3.2.3. Fungicides and Herbicides
3.2.4. Tillage
3.2.5. Regenerative Agricultural Practices and Soil Organic Matter
3.2.6. Genetic Resistance
3.2.7. Soil and Plant Microbiota
3.2.8. Biological Control Agents and Botanical Fungicides
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- EFSA; Knutsen, H.K.; Alexander, J.; Barregård, L.; Bignami, M.; Brüschweiler, B.; Ceccatelli, S.; Cottrill, B.; Dinovi, M.; Grasl-Kraupp, B.; et al. Risks to human and animal health related to the presence of deoxynivalenol and its acetylated and modified forms in food and feed. EFSA J. 2017, 15, e04718. [Google Scholar] [CrossRef] [PubMed]
- EFSA; Knutsen, H.K.; Alexander, J.; Barregård, L.; Bignami, M.; Brüschweiler, B.; Ceccatelli, S.; Cottrill, B.; Dinovi, M.; Edler, L.; et al. Risks for animal health related to the presence of zearalenone and its modified forms in feed. EFSA J. 2017, 15, e04851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- EFSA; Arcella, D.; Gergelova, P.; Innocenti, M.L.; Steinkellner, H. Human and animal dietary exposure to T-2 and HT-2 toxin. EFSA J. 2017, 15, e04972. [Google Scholar] [CrossRef] [PubMed]
- VKM; Bernhoft, A.; Eriksen, G.S.; Sundheim, L.; Berntssen, M.; Brantsæter, A.L.; Brodal, G.; Fæste, C.K.; Hofgaard, I.S.; Rafoss, T.; et al. Risk assessment of mycotoxins in cereal grain in Norway. In Opinion of the Scientific Steering Committee of the Norwegian Scientific Committee for Food Safety (VKM); VKM: Oslo, Norway, 2013; p. 287. [Google Scholar]
- Langseth, W.; Elen, O. The occurrence of deoxynivalenol in Norwegian cereals—differences between years and districts, 1988–1996. Acta Agric. Scand. Sect. B Soil Plant Sci. 1997, 47, 176–184. [Google Scholar] [CrossRef]
- Bottalico, A.; Perrone, G. Toxigenic Fusarium species and mycotoxins associated with head blight in small-grain cereals in Europe. Eur. J. Plant Pathol. 2002, 108, 611–624. [Google Scholar] [CrossRef]
- Bernhoft, A.; Torp, M.; Clasen, P.E.; Løes, A.K.; Kristoffersen, A.B. Influence of agronomic and climatic factors on Fusarium infestation and mycotoxin contamination of cereals in Norway. Food Addit. Contam. Part A 2012, 29, 1129–1140. [Google Scholar] [CrossRef] [Green Version]
- Van Der Fels-Klerx, H.J.; Klemsdal, S.; Hietaniemi, V.; Lindblad, M.; Ioannou-Kakouri, E.; Van Asselt, E.D. Mycotoxin contamination of cereal grain commodities in relation to climate in North West Europe. Food Addit. Contam. Part A 2012, 29, 1581–1592. [Google Scholar] [CrossRef]
- Xu, X.M.; Madden, L.V.; Edwards, S.G. Modeling the effects of environmental conditions on HT2 and T2 toxin accumulation in field oat grains. Phytopathology 2014, 104, 57–66. [Google Scholar] [CrossRef]
- Ferrigo, D.; Raiola, A.; Causin, R. Fusarium toxins in cereals: Occurrence, legislation, factors promoting the appearance and their management. Molecules 2016, 21, 627. [Google Scholar] [CrossRef] [Green Version]
- Persson, T.; Eckersten, H.; Elen, O.; Roer Hjelkrem, A.-G.; Markgren, J.; Söderström, M.; Börjesson, T. Predicting deoxynivalenol in oats under conditions representing Scandinavian production regions. Food Addit. Contam. Part A 2017, 34, 1026–1038. [Google Scholar] [CrossRef]
- Edwards, S.G. Influence of agricultural practices on Fusarium infection of cereals and subsequent contamination of grain by trichothecene mycotoxins. Toxicol. Lett. 2004, 153, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Schöneberg, T.; Martin, C.; Wettstein, F.E.; Bucheli, T.D.; Mascher, F.; Bertossa, M.; Musa, T.; Keller, B.; Vogelgsang, S. Fusarium and mycotoxin spectra in Swiss barley are affected by various cropping techniques. Food Addit. Contam. Part A 2016, 33, 1608–1619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schöneberg, T.; Jenny, E.; Wettstein, F.E.; Bucheli, T.D.; Mascher, F.; Bertossa, M.; Musa, T.; Seifert, K.A.; Gräfenhan, T.; Keller, B.; et al. Occurrence of Fusarium species and mycotoxins in Swiss oats—Impact of cropping factors. Eur. J. Agron. 2018, 92, 123–132. [Google Scholar] [CrossRef]
- Shah, L.; Ali, A.; Yahya, M.; Zhu, Y.; Wang, S.; Si, H.; Rahman, H.; Ma, C. Integrated control of Fusarium head blight and deoxynivalenol mycotoxin in wheat. Plant Pathol. 2018, 67, 532–548. [Google Scholar] [CrossRef]
- Newton, A.C.; Begg, G.S.; Swanston, J.S. Deployment of diversity for enhanced crop function. Ann. Appl. Biol. 2009, 154, 309–322. [Google Scholar] [CrossRef]
- Grettenberger, I.M.; Tooker, J.F. Moving beyond resistance management toward an expanded role for seed mixtures in agriculture. Agric. Ecosyst. Environ. 2015, 208, 29–36. [Google Scholar] [CrossRef]
- Commission implementation regulation (EU). Commission Implementation Regulation (EU) 2016/673 of 29 April 2016 Amending Regulation (EC) No 889/2008 Laying down Detailed Rules for the Implementation of Council Regulation (EC) No 834/2007 on Organic Production and Labelling of Organic Products with Regard to Organic Production, Labelling and Control; Commission implementation regulation (EU): Brussels, Belgium, 2016. [Google Scholar]
- Baker, B.P.; Benbrook, C.M.; Iii, E.G.; Benbrook, K.L. Pesticide residues in conventional, integrated pest management (IPM)-grown and organic foods: Insights from three US data sets. Food Addit. Contam. 2002, 19, 427–446. [Google Scholar] [CrossRef] [PubMed]
- Hansen, A.L. The Organic Farming Manual: A Comprehensive Guide to Starting and Running a Certified Organic Farm; Storey Publishing: North Adams, MA, USA, 2010. [Google Scholar]
- Murphy, K.M.; Campbell, K.G.; Lyon, S.R.; Jones, S.S. Evidence of varietal adaptation to organic farming systems. Field Crops Res. 2007, 102, 172–177. [Google Scholar] [CrossRef] [Green Version]
- Rempelos, L.; Almuayrifi, M.S.B.; Baranski, M.; Tetard-Jones, C.; Barkla, B.; Cakmak, I.; Ozturk, L.; Cooper, J.; Volakakis, N.; Hall, G.; et al. The effect of agronomic factors on crop health and performance of winter wheat varieties bred for the conventional and the low input farming sector. Field Crops Res. 2020, 254, 107822. [Google Scholar] [CrossRef]
- Brodal, G.; Hofgaard, I.S.; Eriksen, G.S.; Bernhoft, A.; Sundheim, L. Mycotoxins in organically versus conventionally produced cereal grains and some other crops in temperate regions. World Mycotoxin J. 2016, 9, 755–770. [Google Scholar] [CrossRef]
- Pussemier, L.; Piérard, J.Y.; Anselme, M.; Tangni, E.K.; Motte, J.C.; Larondelle, Y. Development and application of analytical methods for the determination of mycotoxins in organic and conventional wheat. Food Addit. Contam. 2006, 23, 1208–1218. [Google Scholar] [CrossRef] [PubMed]
- Vogelgsang, S.; Musa, T.; Bänziger, I.; Kägi, A.; Bucheli, T.D.; Wettstein, F.E.; Pasquali, M.; Forrer, H.-R. Fusarium mycotoxins in Swiss wheat: A survey of growers’ samples between 2007 and 2014 shows strong year and minor geographic effects. Toxins 2017, 9, 246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaarschmidt, S.; Fauhl-Hassek, C. The Fate of Mycotoxins During the Processing of Wheat for Human Consumption. Compr. Rev. Food Sci. Food Saf. 2018, 17, 556–593. [Google Scholar] [CrossRef] [Green Version]
- Góral, T.; Łukanowski, A.; Małuszyńska, E.; Stuper-Szablewska, K.; Buśko, M.; Perkowski, J. Performance of winter wheat cultivars grown organically and conventionally with focus on Fusarium head blight and Fusarium trichothecene toxins. Microorganisms 2019, 7, 439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munger, H.; Vanasse, A.; Rioux, S.; Légère, A. Bread wheat performance, Fusarium head blight incidence and weed infestation response to low-input conservation tillage systems in eastern Canada. Can. J. Plant Sci. 2014, 94, 193–201. [Google Scholar] [CrossRef]
- Lacko-Bartošová, M.; Kobida, L. Deoxynivalenol and zearalenone in winter wheat grown in ecological and integrated systems. Res. J. Agric. Sci. 2011, 43, 68–72. [Google Scholar]
- Quaranta, F.; Amoriello, T.; Aureli, G.; Belocchi, A.; D’Egidio, M.G.; Fornara, M.; Melloni, S.; Desiderio, E. Grain yield, quality and deoxynivalenol (DON) contamination of durum wheat (Triticum durum Desf.): Results of national networks in organic and conventional cropping systems. Ital. J. Agron. 2010, 5, 353–366. [Google Scholar] [CrossRef]
- Vanova, M.; Klem, K.; Misa, P.; Matusinsky, P.; Hajslova, J.; Lancová, K. The content of Fusarium mycotoxins, grain yield and quality of winter wheat cultivars under organic and conventional cropping systems. Plant Soil Environ. 2008, 54, 395–402. [Google Scholar] [CrossRef] [Green Version]
- Champeil, A.; Fourbet, J.F.; Doré, T.; Rossignol, L. Influence of cropping system on Fusarium head blight and mycotoxin levels in winter wheat. Crop Prot. 2004, 23, 531–537. [Google Scholar] [CrossRef]
- Schneweis, I.; Meyer, K.; Ritzmann, M.; Hoffmann, P.; Dempfle, L.; Bauer, J. Influence of organically or conventionally produced wheat on health, performance and mycotoxin residues in tissues and bile of growing pigs. Arch. Anim. Nutr. 2005, 59, 155–163. [Google Scholar] [CrossRef]
- Mäder, P.; Hahn, D.; Dubois, D.; Gunst, L.; Alföldi, T.; Bergmann, H.; Oehme, M.; Amadò, R.; Schneider, H.; Graf, U.; et al. Wheat quality in organic and conventional farming: Results of a 21 year field experiment. J. Sci. Food Agric. 2007, 87, 1826–1835. [Google Scholar] [CrossRef]
- Griesshaber, D.; Kuhn, F.; Berger, U.; Oehme, M. Comparison of trichothecene contaminations in wheat cultivated by three different farming systems in Switzerland: Biodynamic, bioorganic and conventional. Mitt. Lebensm. Hyg. 2004, 95, 251–260. [Google Scholar]
- Birzele, B.; Meier, A.; Hindorf, H.; Krämer, J.; Dehne, H.W. Epidemiology of Fusarium infection and deoxynivalenol content in winter wheat in the Rhineland, Germany. Eur. J. Plant Pathol. 2002, 108, 667–673. [Google Scholar] [CrossRef]
- Hietaniemi, V.; Kontturi, M.; Rämö, S.; Eurola, M.; Kangas, A.; Niskanen, M.; Saastamoinen, M. Contents of trichothecenes in oats during official variety, organic cultivation and nitrogen fertilization trials in Finland. Agric. Food Sci. 2004, 13, 54–67. [Google Scholar] [CrossRef]
- Polišenská, I.; Jirsa, O.; Salava, J.; Sedláčková, I.; Frydrych, J. Fusarium mycotoxin content and Fusarium species presence in Czech organic and conventional wheat. World Mycotoxin J. 2021, 14, 201–211. [Google Scholar] [CrossRef]
- Giménez, I.; Escobar, J.; Ferruz, E.; Lorán, S.; Herrera, M.; Juan, T.; Herrera, A.; Ariño, A. The effect of weather and agronomic practice on deoxynivalenol mycotoxin in durum wheat. J. Life Sci. 2012, 49, 513–517. [Google Scholar]
- Bernhoft, A.; Clasen, P.-E.; Kristoffersen, A.B.; Torp, M. Less Fusarium infestation and mycotoxin contamination in organic than in conventional cereals. Food Addit. Contam. Part A 2010, 27, 842–852. [Google Scholar] [CrossRef]
- Edwards, S.G. Fusarium mycotoxin content of UK organic and conventional wheat. Food Addit. Contam. Part A 2009, 26, 496–506. [Google Scholar] [CrossRef] [Green Version]
- Meister, U. Fusarium toxins in cereals of integrated and organic cultivation from the Federal State of Brandenburg (Germany) harvested in the years 2000–2007. Mycotoxin Res. 2009, 25, 133–139. [Google Scholar] [CrossRef]
- Döll, S.; Valenta, H.; Dänicke, S.; Flachowsky, G. Fusarium mycotoxins in conventionally and organically grown grain from Thuringia/Germany. Landbauforsch. Völkenrode 2002, 2, 91–96. [Google Scholar] [CrossRef]
- Marx, H.; Gedek, B.; Kollarczik, B. Vergleichende Untersuchungen zum mykotoxikologischen Status von ökologisch und konventionell angebautem Getreide. Z. Lebensm.-Unters. Forsch. 1995, 201, 83–86. [Google Scholar] [CrossRef] [PubMed]
- Edwards, S.G. Fusarium mycotoxin content of UK organic and conventional barley. Food Addit. Contam. Part A 2009, 26, 1185–1190. [Google Scholar] [CrossRef] [Green Version]
- Wang, J. Effect of Organic and Conventional Agronomic Practices and Variety Choice on Nutritional Quality, the Contents of Undesirable Compounds and Yield of Cereals. Ph.D. Thesis, Newcastle University, Newcastle, UK, 2018. [Google Scholar]
- Kolawole, O.; De Ruyck, K.; Greer, B.; Meneely, J.; Doohan, F.M.; Danaher, M.; Elliott, C. Agronomic factors influencing the scale of Fusarium mycotoxin contamination of oats. J. Fungi 2021, 7, 965. [Google Scholar] [CrossRef] [PubMed]
- Edwards, S.G. Fusarium mycotoxin content of UK organic and conventional oats. Food Addit. Contam. Part A 2009, 26, 1063–1069. [Google Scholar] [CrossRef] [Green Version]
- Gottschalk, C.; Barthel, J.; Engelhardt, G.; Bauer, J.; Meyer, K. Occurrence of type A trichothecenes in conventionally and organically produced oats and oat products. Mol. Nutr. Food Res. 2007, 51, 1547–1553. [Google Scholar] [CrossRef]
- Wang, J.; Hasanalieva, G.; Wood, L.; Markellou, E.; Iversen, P.O.; Bernhoft, A.; Seal, C.; Baranski, M.; Vigar, V.; Ernst, L.; et al. Effect of wheat species (Triticum aestivum vs T. spelta), farming system (organic vs conventional) and flour type (wholegrain vs white) on composition of wheat flour; results of a retail survey in the UK and Germany—1. Mycotoxin content. Food Chem. 2020, 327, 127011. [Google Scholar] [CrossRef]
- Dänicke, S.; Gareis, M.; Bauer, J. Orientation values for critical concentrations of deoxynivalenol and zearalenone in diets for pigs, ruminants and gallinaceous poultry. Proc. Soc. Nutr. Physiol. USA 2001, 10, 171–174. [Google Scholar]
- Beyer, M.; Klix, M.B.; Klink, H.; Verreet, J.A. Quantifying the effects of previous crop, tillage, cultivar and triazole fungicides on the deoxynivalenol content of wheat grain—A review. J. Plant Dis. Prot. 2006, 113, 241–246. [Google Scholar] [CrossRef]
- Schaafsma, A.W.; Tamburic-Ilincic, L.; Miller, J.; Hooker, D.C. Agronomic considerations for reducing deoxynivalenol in wheat grain. Can. J. Plant Pathol. 2001, 23, 279–285. [Google Scholar] [CrossRef]
- Pageau, D.; Lafond, J.; Lajeunesse, J.; Savard, M.E. Impact du précédent cultural et de la fertilisation azotée sur la teneur en désoxynivalénol chez l’orge. Can. J. Plant Pathol. 2008, 30, 397–403. [Google Scholar] [CrossRef]
- Dill-Macky, R.; Jones, R.K. The effect of previous crop residues and tillage on Fusarium head blight of wheat. Plant Dis. 2000, 84, 71–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Champeil, A.; Doré, T.; Fourbet, J.F. Fusarium head blight: Epidemiological origin of the effects of cultural practices on head blight attacks and the production of mycotoxins by Fusarium in wheat grains. Plant Sci. 2004, 166, 1389–1415. [Google Scholar] [CrossRef]
- Koch, H.-J.; Pringas, C.; Maerlaender, B. Evaluation of environmental and management effects on Fusarium head blight infection and deoxynivalenol concentration in the grain of winter wheat. Eur. J. Agron. 2006, 24, 357–366. [Google Scholar] [CrossRef]
- Köpke, U.; Thiel, B.; Elmholt, S. Strategies to reduce mycotoxin and fungal alkaloid contamination in organic and conventional cereal production systems. In Handbook of Organic Food Safety and Quality; Cooper, J., Niggli, U., Leifert, C., Eds.; Woodhead Publishing: Cambridge, UK, 2007; pp. 353–391. [Google Scholar]
- Wegulo, S.N.; Baenziger, P.S.; Hernandez Nopsa, J.; Bockus, W.W.; Hallen-Adams, H. Management of Fusarium head blight of wheat and barley. Crop Prot. 2015, 73, 100–107. [Google Scholar] [CrossRef]
- Edwards, S.G. Impact of agronomic and climatic factors on the mycotoxin content of harvested oats in the United Kingdom. Food Addit. Contam. Part A 2017, 34, 2230–2241. [Google Scholar] [CrossRef]
- Schwartau, V.; Zozulia, O.; Mykhalska, L.; Sanin, O. Strategies of decreasing harmfulness of fusariosis agents in agrophytocenoses. Agric. Sci. Pract. 2018, 5, 60–70. [Google Scholar] [CrossRef] [Green Version]
- Vogelgsang, S.; Beyer, M.; Pasquali, M.; Jenny, E.; Musa, T.; Bucheli, T.D.; Wettstein, F.E.; Forrer, H.-R. An eight-year survey of wheat shows distinctive effects of cropping factors on different Fusarium species and associated mycotoxins. Eur. J. Agron. 2019, 105, 62–77. [Google Scholar] [CrossRef]
- Xia, R.; Schaafsma, A.W.; Wu, F.; Hooker, D.C. Impact of the improvements in Fusarium head blight and agronomic management on economics of winter wheat. World Mycotoxin J. 2020, 13, 423–439. [Google Scholar] [CrossRef]
- Drakopoulos, D.; Kägi, A.; Six, J.; Zorn, A.; Wettstein, F.E.; Bucheli, T.D.; Forrer, H.-R.; Vogelgsang, S. The agronomic and economic viability of innovative cropping systems to reduce Fusarium head blight and related mycotoxins in wheat. Agric. Syst. 2021, 192, 103198. [Google Scholar] [CrossRef]
- Fagard, M.; Launay, A.; Clément, G.; Courtial, J.; Dellagi, A.; Farjad, M.; Krapp, A.; Soulié, M.-C.; Masclaux-Daubresse, C. Nitrogen metabolism meets phytopathology. J. Exp. Bot. 2014, 65, 5643–5656. [Google Scholar] [CrossRef]
- Lemmens, M.; Haim, K.; Lew, H.; Ruckenbauer, P. The effect of nitrogen fertilization on Fusarium head blight development and deoxynivalenol contamination in wheat. J. Phytopathol. 2004, 152, 1–8. [Google Scholar] [CrossRef]
- Heier, T.; Jain, S.K.; Kogel, K.H.; Pons-Kühnemann, J. Influence of N-fertilization and fungicide strategies on Fusarium head blight severity and mycotoxin content in winter wheat. J. Phytopathol. 2005, 153, 551–557. [Google Scholar] [CrossRef]
- Martin, R.; Macleod, J.; Caldwell, C. Influences of production inputs on incidence of infection by Fusarium species on cereal seed. Plant Dis. 1991, 75, 784–788. [Google Scholar] [CrossRef]
- Yi, C.; Kaul, H.-P.; Kübler, E.; Schwadorf, K.; Aufhammer, W. Head blight (Fusarium graminearum) and deoxynivalenol concentration in winter wheat as affected by pre-crop, soil tillage and nitrogen fertilization. Z. Pflanzenkrankh. Pflanzenschutz 2001, 108, 217–230. [Google Scholar]
- Teich, A.H. Less wheat scab with urea than with ammonium nitrate fertilisers. Cereal Res. Commun. 1987, 15, 35–38. [Google Scholar]
- Van Der Burgt, G.J.H.M.; Timmermans, B.G.H.; Scholberg, J.M.S.; Osman, A.M. Fusarium head blight and deoxynivalenol contamination in wheat as affected by nitrogen fertilization. NJAS Wagening. J. Life Sci. 2011, 58, 123–129. [Google Scholar] [CrossRef] [Green Version]
- Subedi, K.D.; Ma, B.L.; Xue, A.G. Planting date and nitrogen effects on Fusarium head blight and leaf spotting diseases in spring wheat. Agron. J. 2007, 99, 113–121. [Google Scholar] [CrossRef]
- Aufhammer, W.; Kubler, E.; Kaul, H.-P.; Hermann, W.; Hohn, D.; Yi, C. Infection with head blight (F. graminearum, F. culmorum) and deoxynivalenol concentration in winter wheat as influenced by N fertilization. Pflanzenbauwissenschaften 2000, 4, 72–78. [Google Scholar]
- Fauzi, M.; Paulitz, T. The effect of plant growth regulators and nitrogen on Fusarium head blight of the spring wheat cultivar Max. Plant Dis. 1994, 78, 289–292. [Google Scholar] [CrossRef]
- Hofer, K.; Barmeier, G.; Schmidhalter, U.; Habler, K.; Rychlik, M.; Hückelhoven, R.; Hess, M. Effect of nitrogen fertilization on Fusarium head blight in spring barley. Crop Prot. 2016, 88, 18–27. [Google Scholar] [CrossRef]
- Yang, F.; Jensen, J.D.; Spliid, N.H.; Svensson, B.; Jacobsen, S.; Jørgensen, L.N.; Jørgensen, H.J.L.; Collinge, D.B.; Finnie, C. Investigation of the effect of nitrogen on severity of Fusarium head blight in barley. J. Proteom. 2010, 73, 743–752. [Google Scholar] [CrossRef] [PubMed]
- Fauci, M.F.; Dick, R.P. Soil microbial dynamics: Short-and long-term effects of inorganic and organic nitrogen. Soil Sci. Soc. Am. J. 1994, 58, 801–806. [Google Scholar] [CrossRef]
- Yu, H.; Ding, W.; Luo, J.R.G.; Cai, Z. Long-term application of compost and mineral fertilizers on aggregation and aggregate-associated carbon in a sandy loam soil. Soil Tillage Res. 2012, 124, 170–177. [Google Scholar] [CrossRef]
- Gryndler, M.; Larsen, J.; Hrselová, H.; Řezáčová, V.; Gryndlerová, H.; Kubát, J. Organic and mineral fertilization, respectively, increase and decrease the development of external mycelium of arbuscular mycorrhizal fungi in a long-term field experiment. Mycorrhiza 2006, 16, 159–166. [Google Scholar] [CrossRef]
- Van Arendonk, J.J.; Niemann, G.; Boon, J.; Lambers, H. Effects of nitrogen supply on the anatomy and chemical composition of leaves of four grass species belonging to the genus Poa, as determined by image processing analysis and pyrolysis-mass spectrometry. Plant Cell Environ. 1997, 20, 881–897. [Google Scholar] [CrossRef]
- Paul, P.A.; Lipps, P.E.; Hershman, D.E.; McMullen, M.P.; Draper, M.A.; Madden, L.V. Efficacy of triazole-based fungicides for Fusarium head blight and deoxynivalenol control in wheat: A multivariate meta-analysis. Phytopathology 2008, 98, 999–1011. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, J.; Haber, S. Overview of some recent research developments in Fusarium head blight of wheat. Can. J. Plant Pathol. 2013, 35, 149–174. [Google Scholar] [CrossRef]
- Haidukowski, M.; Visconti, A.; Perrone, G.; Vanadia, S.; Pancaldi, D.; Covarelli, L.; Balestrazzi, R.; Pascale, M. Effect of prothioconazole-based fungicides on Fusarium head blight, grain yield and deoxynivalenol accumulation in wheat under field conditions. Phytopathol. Mediterr. 2012, 51, 236–246. [Google Scholar] [CrossRef]
- Kharbikar, L.L.; Dickin, E.T.; Edwards, S.G. Impact of post-anthesis rainfall, fungicide and harvesting time on the concentration of deoxynivalenol and zearalenone in wheat. Food Addit. Contam. Part A 2015, 32, 2075–2085. [Google Scholar] [CrossRef]
- Hudec, K.; Kičinová, J.; Mihók, M. Changes of species spectrum associated with Fusarium head blight caused by fungicides. J. Cent. Eur. Agric. 2019, 20, 376–388. [Google Scholar] [CrossRef]
- Spolti, P.; Del Ponte, E.M.; Dong, Y.; Cummings, J.A.; Bergstrom, G.C. Triazole sensitivity in a contemporary population of Fusarium graminearum from New York wheat and competitiveness of a tebuconazole-resistant isolate. Plant Dis. 2014, 98, 607–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simpson, D.R.; Weston, G.E.; Turner, J.A.; Jennings, P.; Nicholson, P. Differential control of head blight pathogens of wheat by fungicides and consequences for mycotoxin contamination of grain. Eur. J. Plant Pathol. 2001, 107, 421–431. [Google Scholar] [CrossRef]
- Magan, N.; Hope, R.; Colleate, A.; Baxter, E.S. Relationship between growth and mycotoxin production by Fusarium species, biocides and environment. Eur. J. Plant Pathol. 2002, 108, 685–690. [Google Scholar] [CrossRef]
- Ioos, R.; Belhadj, A.; Menez, M.; Faure, A. The effects of fungicides on Fusarium spp. and Microdochium nivale and their associated trichothecene mycotoxins in French naturally-infected cereal grains. Crop Prot. 2005, 24, 894–902. [Google Scholar] [CrossRef]
- Edwards, S.G.; Anderson, E. Impact of agronomy on HT-2 and T-2 toxin content of oats. Plant Breed. Seed Sci. 2011, 63, 49–57. [Google Scholar] [CrossRef]
- Karron, E.; Runno-Paurson, E.; Lõiveke, H.; Islamov, B.; Kütt, M.-L.; Talve, T.; Lauringson, E.; Hõrak, H.; Edesi, L.; Niinemets, Ü. Application of widely used fungicides does not necessarily affect grain yield, and incidence of Fusarium spp. and mycotoxins DON, HT-2 and T-2 in spring barley in northern climates. Kvas. Prum. 2020, 66, 215–223. [Google Scholar] [CrossRef] [Green Version]
- Altman, J.; Rovira, A.D. Herbicide-pathogen interactions in soil-borne root diseases. Can. J. Plant Pathol. 1989, 11, 166–172. [Google Scholar] [CrossRef]
- Fernandez, M.R.; Zentner, R.P.; Basnyat, P.; Gehl, D.; Selles, F.; Huber, D. Glyphosate associations with cereal diseases caused by Fusarium spp. in the Canadian Prairies. Eur. J. Agron. 2009, 31, 133–143. [Google Scholar] [CrossRef]
- Martinez, D.A.; Loening, U.E.; Graham, M.C. Impacts of glyphosate-based herbicides on disease resistance and health of crops: A review. Environ. Sci. Eur. 2018, 30, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Rose, T.J.; Van Zwieten, L.; Claassens, A.; Scanlan, C.; Rose, M.T. Phytotoxicity of soilborne glyphosate residues is influenced by the method of phosphorus fertiliser application. Plant Soil 2018, 422, 455–465. [Google Scholar] [CrossRef]
- Miller, J.D.; Culley, J.; Fraser, K.; Hubbard, S.; Meloche, F.; Ouellet, T.; Seaman, W.L.; Seifert, K.A.; Turkington, K.; Voldeng, H. Effect of tillage practice on Fusarium head blight of wheat. Can. J. Plant Pathol. 1998, 20, 95–103. [Google Scholar] [CrossRef]
- Hofgaard, I.S.; Seehusen, T.; Aamot, H.U.; Riley, H.; Razzaghian, J.; Le, V.H.; Hjelkrem, A.-G.R.; Dill-Macky, R.; Brodal, G. Inoculum potential of Fusarium spp. relates to tillage and straw management in Norwegian fields of spring oats. Front. Microbiol 2016, 7, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hofgaard, I.S.; Aamot, H.U.; Seehusen, T.; Riley, H.; Dill-Macky, R.; Holen, B.; Brodal, G. Fusarium and mycotoxin content of harvested grain was not related to tillage intensity in Norwegian spring wheat fields. World Mycotoxin J. 2020, 13, 473–486. [Google Scholar] [CrossRef]
- Alabouvette, C. Fusarium wilt suppressive soils: An example of disease-suppressive soils. Australas. Plant Pathol. 1999, 28, 57–64. [Google Scholar] [CrossRef]
- Peigné, J.; Messmer, M.; Aveline, A.; Berner, A.; Mäder, P.; Carcea, M.; Narducci, V.; Samson, M.-F.; Thomsen, I.K.; Celette, F.; et al. Wheat yield and quality as influenced by reduced tillage in organic farming. Org. Agric. 2014, 4, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Wong, P.T.W. Effect of Burkholderia (Pseudomonas) cepacia and soil type on the control of crown rot in wheat. Plant Soil 1998, 203, 103–108. [Google Scholar] [CrossRef]
- Knudsen, I.M.B.; Debosz, K.; Hockenhull, J.; Jensen, D.F.; Elmholt, S. Suppressiveness of organically and conventionally managed soils towards brown foot rot of barley. Appl. Soil Ecol. 1999, 12, 61–72. [Google Scholar] [CrossRef]
- Kurek, E.; Jaroszuk-Ściseł, J. Rye (Secale cereale) growth promotion by Pseudomonas fluorescens strains and their interactions with Fusarium culmorum under various soil conditions. Biol. Control. 2003, 26, 48–56. [Google Scholar] [CrossRef]
- Buerstmayr, M.; Steiner, B.; Buerstmayr, H. Breeding for Fusarium head blight resistance in wheat—Progress and challenges. Plant Breed. 2020, 139, 429–454. [Google Scholar] [CrossRef]
- Ghimire, B.; Sapkota, S.; Bahri, B.A.; Martinez-Espinoza, A.D.; Buck, J.W.; Mergoum, M. Fusarium head blight and rust diseases in soft red winter wheat in the Southeast United States: State of the art, challenges and future perspective for breeding. Front Plant Sci. 2020, 11, 1080. [Google Scholar] [CrossRef]
- Bissonnette, K.M.; Kolb, F.L.; Ames, K.A.; Bradley, C.A. Effect of wheat cultivar on the concentration of Fusarium mycotoxins in wheat stems. Plant Dis. 2018, 102, 2539–2544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, F.; Wu, J.; Zhao, H.; Xu, J.; Shi, J. Relationship of deoxynivalenol content in grain, chaff, and straw with Fusarium head blight severity in wheat varieties with various levels of resistance. Toxins 2015, 7, 728–742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perincherry, L.; Lalak-Kańczugowska, J.; Stępień, Ł. Fusarium-produced mycotoxins in plant-pathogen interactions. Toxins 2019, 11, 664. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.; Sato, M.; Takeuchi, T. Evaluation of the effects of five QTL regions on Fusarium head blight resistance and agronomic traits in spring wheat (Triticum aestivum L.). Breed. Sci. 2012, 62, 11–17. [Google Scholar] [CrossRef] [Green Version]
- Jenkinson, P.; Parry, D.W. Splash dispersal of conidia of Fusarium culmorum and Fusarium avenaceum. Mycol. Res. 1994, 98, 506–510. [Google Scholar] [CrossRef]
- Magistrali, A.; Vavera, R.; Janovská, D.; Rempelos, L.; Cakmak, I.; Leifert, C.; Grausgruber, H.; Butler, G.; Wilkinson, A.; Bilsborrow, P. Evaluating the effect of agronomic management practices on the performance of differing spelt (Triticum spelta) cultivars in contrasting environments. Field Crops Res. 2020, 255, 107869. [Google Scholar] [CrossRef]
- Lori, M.; Symnaczik, S.; Mäder, P.; De Deyn, G.; Gattinger, A. Organic farming enhances soil microbial abundance and activity—A meta-analysis and meta-regression. PLoS ONE 2017, 12, e0180442. [Google Scholar] [CrossRef]
- Xiong, W.; Jousset, A.; Guo, S.; Karlsson, I.; Zhao, Q.; Wu, H.; Kowalchuk, G.A.; Shen, Q.; Li, R.; Geisen, S. Soil protist communities form a dynamic hub in the soil microbiome. Isme J. 2018, 12, 634–638. [Google Scholar] [CrossRef] [Green Version]
- Bertola, M.; Ferrarini, A.; Visioli, G. Improvement of soil microbial diversity through sustainable agricultural practices and its evaluation by -omics approaches: A perspective for the environment, food quality and human safety. Microorganisms 2021, 9, 1400. [Google Scholar] [CrossRef]
- Karlsson, I.; Friberg, H.; Kolseth, A.K.; Steinberg, C.; Persson, P. Organic farming increases richness of fungal taxa in the wheat phyllosphere. Mol. Ecol. 2017, 26, 3424–3436. [Google Scholar] [CrossRef]
- Karlsson, I.; Friberg, H.; Steinberg, C.; Persson, P. Fungicide effects on fungal community composition in the wheat phyllosphere. PLoS ONE 2014, 9, e111786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karlsson, I.; Friberg, H.; Kolseth, A.K.; Steinberg, C.; Persson, P. Agricultural factors affecting Fusarium communities in wheat kernels. Int. J. Food Microbiol. 2017, 252, 53–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karlsson, I.; Persson, P.; Friberg, H. Fusarium head blight from a microbiome perspective. Front. Microbiol. 2021, 12, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Luz, W.C.d.; Stockwell, C.A.; Bergstrom, G.C. Biological control of Fusarium graminearum. In Fusarium Head Blight of Wheat and Barley; Leonard, K.J., Bushnell, W.R., Eds.; American Phytopathological Society (APS Press): St. Paul, MI, USA, 2003; pp. 381–394. [Google Scholar]


{kind=link}
| Study Type Cereal Species | Country | Sampling Year (s) | No. of Samples Org/Con 1 | % Positive Samples Org/Con 1 | Mean Org/Con 1 (µg/kg) | Median Org/Con 1 (µg/kg) | Result of Statistical Analysis | Reference No. |
|---|---|---|---|---|---|---|---|---|
| Experiments | ||||||||
| Wheat | Poland | 2014 | 64/85 | NS | [27] | |||
| Wheat | Canada | 2009 | 3900/5500 | O < C | [28] | |||
| Wheat | Canada | 2010 | 340/460 | NS | [28] | |||
| Wheat | Slovakia | 2007–2008 | 192/362 | O < C | [29] | |||
| Durum | Italy | 2006–2008 | 27/74 | O < C | [30] | |||
| Wheat | Czechia | 2004–2006 | 151/369 2 | O < C | [31] | |||
| Wheat | Czechia 3 | 2004–2006 | 151/246 3 | NS | [31] | |||
| Wheat | France | 2000–2002 | 270/460 | NS | [32] | |||
| Wheat | Germany | 1999–2001 | 179/283 4 | O < C | [33] | |||
| Wheat | Switzerland | 1998,2000 | 100/100 | 49/82 | O < C | [34,35] | ||
| Wheat | Germany | 1997–1998 | 200/265 4 | O < C | [36] | |||
| Oats | Finland | 1997–1998 | 109/148 | NS | [37] | |||
| Surveys | ||||||||
| Wheat | Czechia | 2015–2017 | 154/330 | 12/21 | <20 5/80 | <20/<20 | O < C | [38] |
| Wheat 6 | Czechia | 2015–2017 | 154/154 6 | 12/14 | <20 5/23 | <20/<20 | NS | [38] |
| Durum | Spain | 2006–2007 | 50/67 | 28/31 | 95/194 | NS | [39] | |
| Wheat 6 | Norway | 2002–2004 | 92/92 6 | 86/170 | 29/51 | O < C | [40] | |
| Wheat | Belgium | 2002–2003 | 51/42 | 96/100 | 204/493 | O < C | [24] | |
| Wheat | UK | 2001–2005 | 247/1377 | 86 | 230 7 | 42 7 | NS | [41] |
| Wheat | Germany | 2000–2007 | 110/355 | 23/42 | 55/242 4,8 | O < C | [42] | |
| Wheat | Germany | 1998 | 46/150 | 54/69 | 760/1540 | 230/270 | O < C | [43] |
| Wheat | Germany | 1991 | 50/51 | 76/88 | 381/376 | NS | [44] | |
| Oats 6 | Norway | 2002–2004 | 101/101 6 | 114/426 | 24/36 | NS | [40] | |
| Barley | Switzerland | 2013–2014 | 42/225 | 24/201 | O < C | [13] | ||
| Barley | UK | 2002–2005 | 108/338 | 57 7 | 19 7 | 11 7 | NS | [45] |
| Barley 6 | Norway | 2002–2004 | 108/108 6 | 44/44 | <20/<20 | NS | [40] | |
| Rye | Germany | 2000–2007 | 173/337 | 14/36 | <50 9/62 4,8 | O < C | [42] | |
| Rye | Germany | 1991 | 50/50 | 56/40 | 261/94 | O > C | [44] |
| Study Type Cereal Species | Country | Sampling Year (s) | No. of Samples Org/Con 1 | % Positive Samples Org/Con 1 | Mean Org/Con 1 (µg/kg) | Median Org/Con 1 (µg/kg) | Result of Statistical Analysis | Ref. No. |
|---|---|---|---|---|---|---|---|---|
| Experiments | ||||||||
| Wheat | Slovakia | 2007–2008 | 8/7 | NS | [29] | |||
| Wheat | Germany | 1999–2001 | <5 2/28 | O < C | [33] | |||
| Surveys | ||||||||
| Wheat | Czechia | 2015–2017 | 154/330 | 4/9 | <2 3/3 | <2/<2 | NS | [38] |
| Wheat | Belgium | 2002–2003 | 51/42 | 27/45 | 10/39 | O < C | [24] | |
| Wheat | Germany | 2001–2007 | 94/308 | 5/16 | 3/11 4 | O < C | [42] | |
| Wheat | UK | 2001–2005 | 247/1377 | 17 5 | <5 5 | NS | [41] | |
| Wheat | Germany | 1991 | 50/51 | 36/16 | 9/1 | O > C | [44] |
| Study Type Cereal Species | Country | Sampling Year (s) | No. of Samples Org/Con 1 | % Positive Samples Org/Con 1 | Mean Org/Con 1 (µg/kg) | Median Org/Con 1 (µg/kg) | Result of Statistical Analysis | Ref. No. |
|---|---|---|---|---|---|---|---|---|
| Experiments | ||||||||
| Wheat | Switzerland | 1998–2000 | 13/44 | ¼ 2,3 | O < C | [35] | ||
| Surveys | ||||||||
| Wheat | UK | 2001–2005 | 247/1377 | 20/36 | <10 4/11 2 | O < C | [41] | |
| Oats | Ireland | 2020 | 114/86 | 52/74 | 137/397 5 | O < C | [47] | |
| Oats | UK | 2002–2005 | 115/343 | 78/97 | 50/264 | 49/292 | O < C | [48] |
| Oats | Germany | 2005 | 35/35 | 100/100 | 8/27 | O < C | [49] | |
| Oats 6 | Norway | 2002–2004 | 101/101 6 | 80/117 2 | <20/62 2 | O < C | [40] | |
| Barley | UK | 2002–2005 | 108/338 | 36 2,7 | 10 2,7 | <10 7 | NS | [45] |
| Barley 6 | Norway | 2002–2004 | 108/108 6 | <20 8/212 | <20/<20 2 | O < C | [40] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bernhoft, A.; Wang, J.; Leifert, C. Effect of Organic and Conventional Cereal Production Methods on Fusarium Head Blight and Mycotoxin Contamination Levels. Agronomy 2022, 12, 797. https://doi.org/10.3390/agronomy12040797
Bernhoft A, Wang J, Leifert C. Effect of Organic and Conventional Cereal Production Methods on Fusarium Head Blight and Mycotoxin Contamination Levels. Agronomy. 2022; 12(4):797. https://doi.org/10.3390/agronomy12040797
Chicago/Turabian StyleBernhoft, Aksel, Juan Wang, and Carlo Leifert. 2022. "Effect of Organic and Conventional Cereal Production Methods on Fusarium Head Blight and Mycotoxin Contamination Levels" Agronomy 12, no. 4: 797. https://doi.org/10.3390/agronomy12040797