
The Adenine at the 4th Exon of the DRO1 Gene Provides Drought-Tolerance Capacity to Hybrid Rice Deyou4727 and Its Maintainer Line Dexiang074B

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Experimental Design
2.3. Samples and Measurements
2.3.1. Root Distribution and Morphology
2.3.2. Antioxidant Enzyme Activity
2.3.3. Gene Expression
2.3.4. Deeper Rooting 1 (DRO1) Gene Sequencing
2.3.5. Tools and Instruments
2.4. Statistical Methods
3. Results
3.1. Response of Yield and Yield Components under Irrigation and Drought Conditions
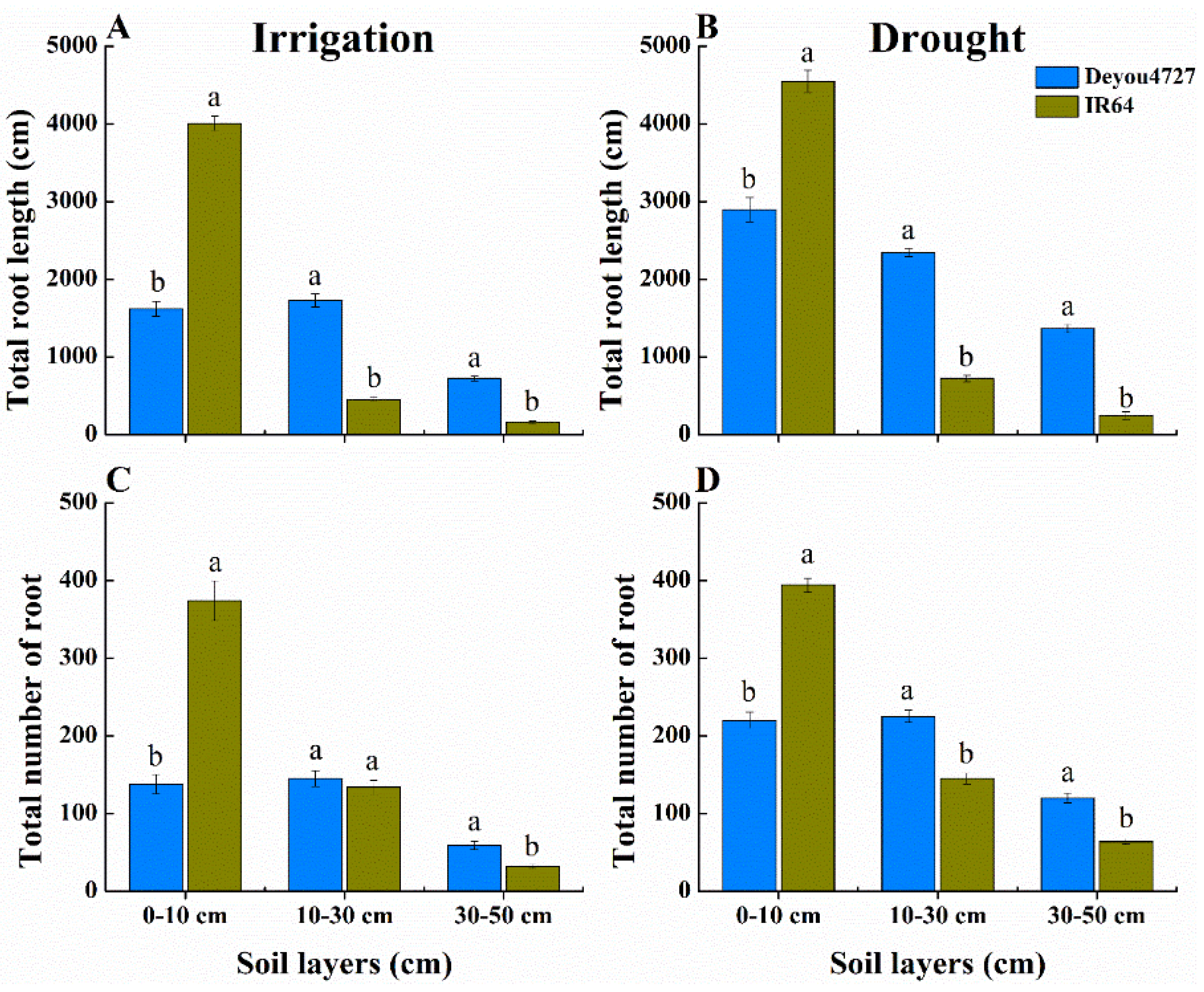
3.2. Root Distribution of Deyou4727 in 0–50 cm Soil
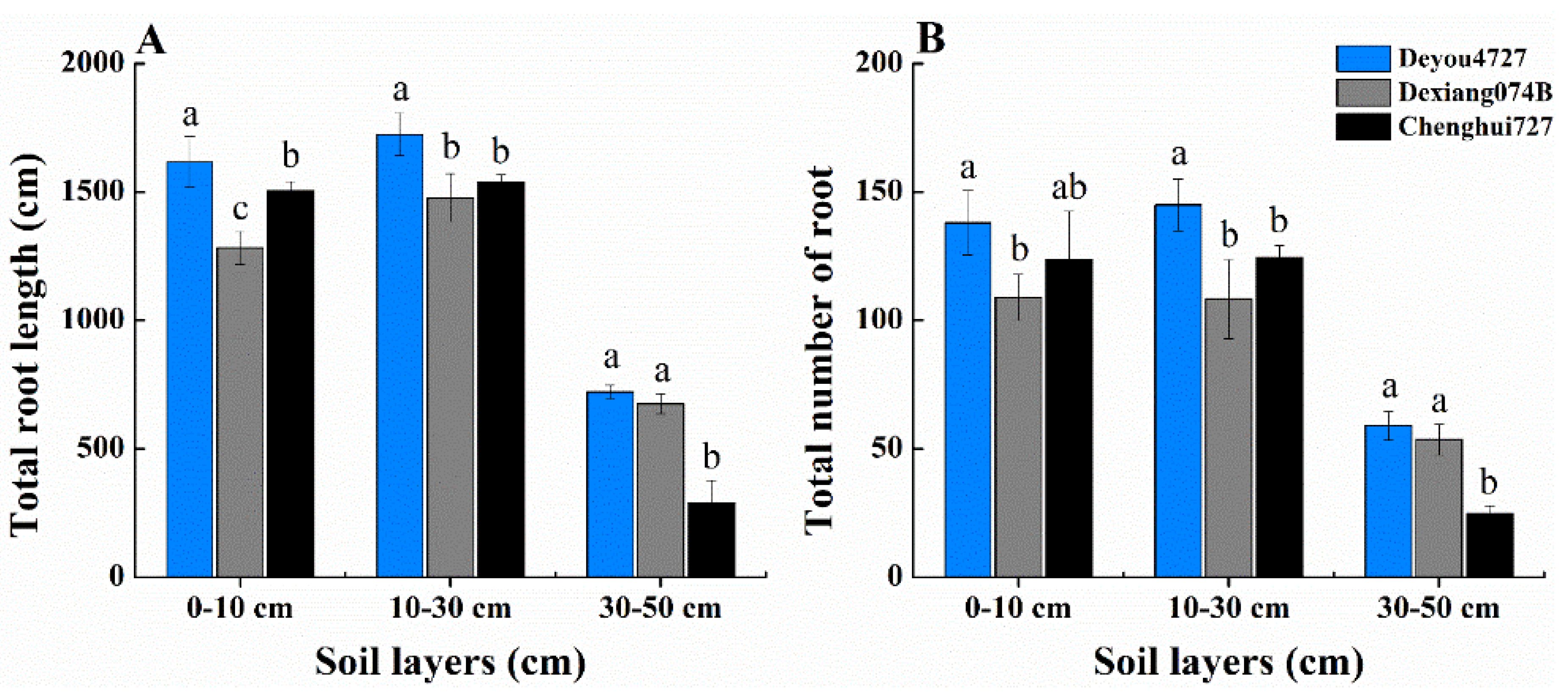
3.3. Root Distribution and Morphology Comparison with Parents
3.4. The Activity of the Antioxidant Enzyme
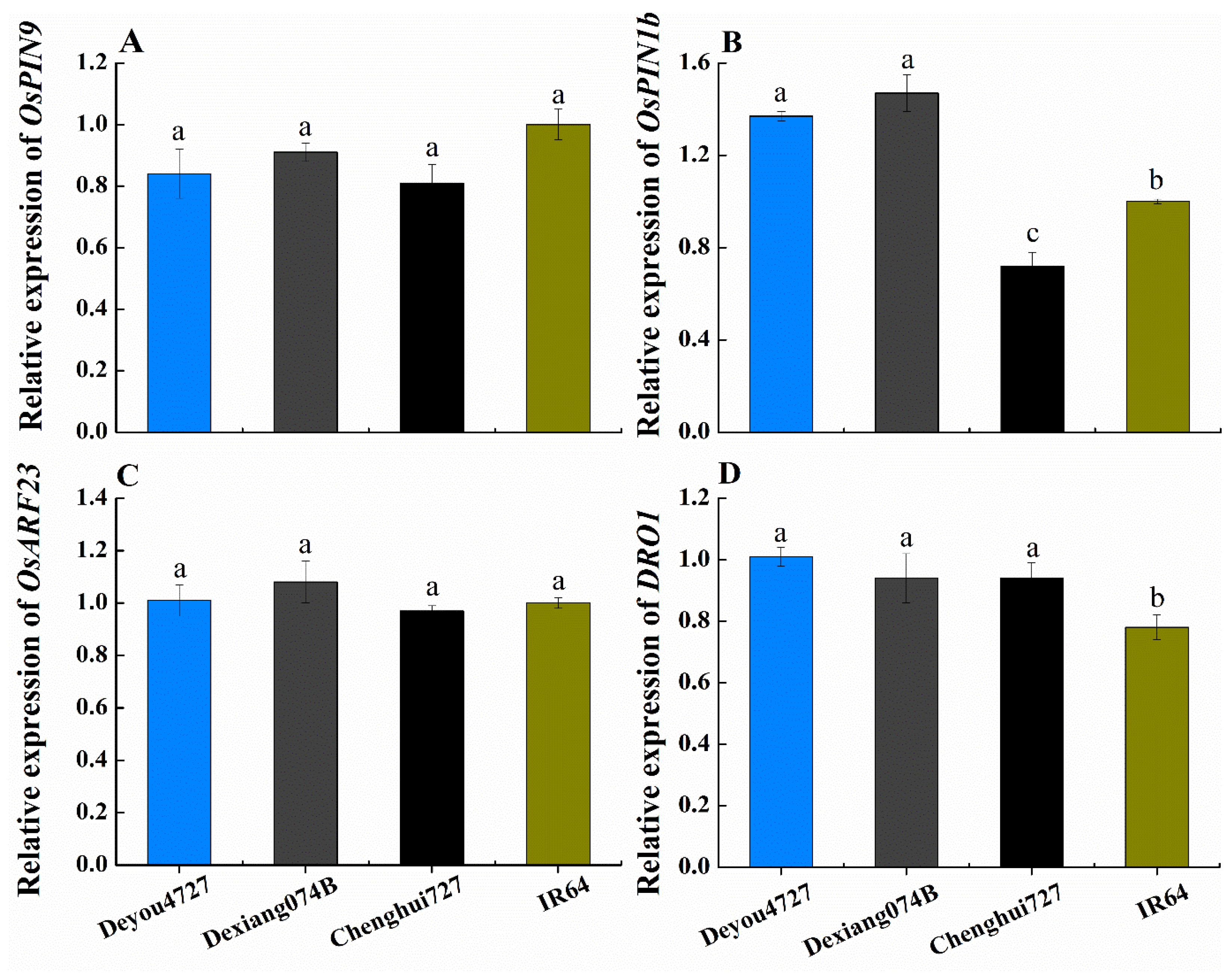
3.5. Gene Expression of OsPIN9, OsPIN1b, OsARF23 and DRO1
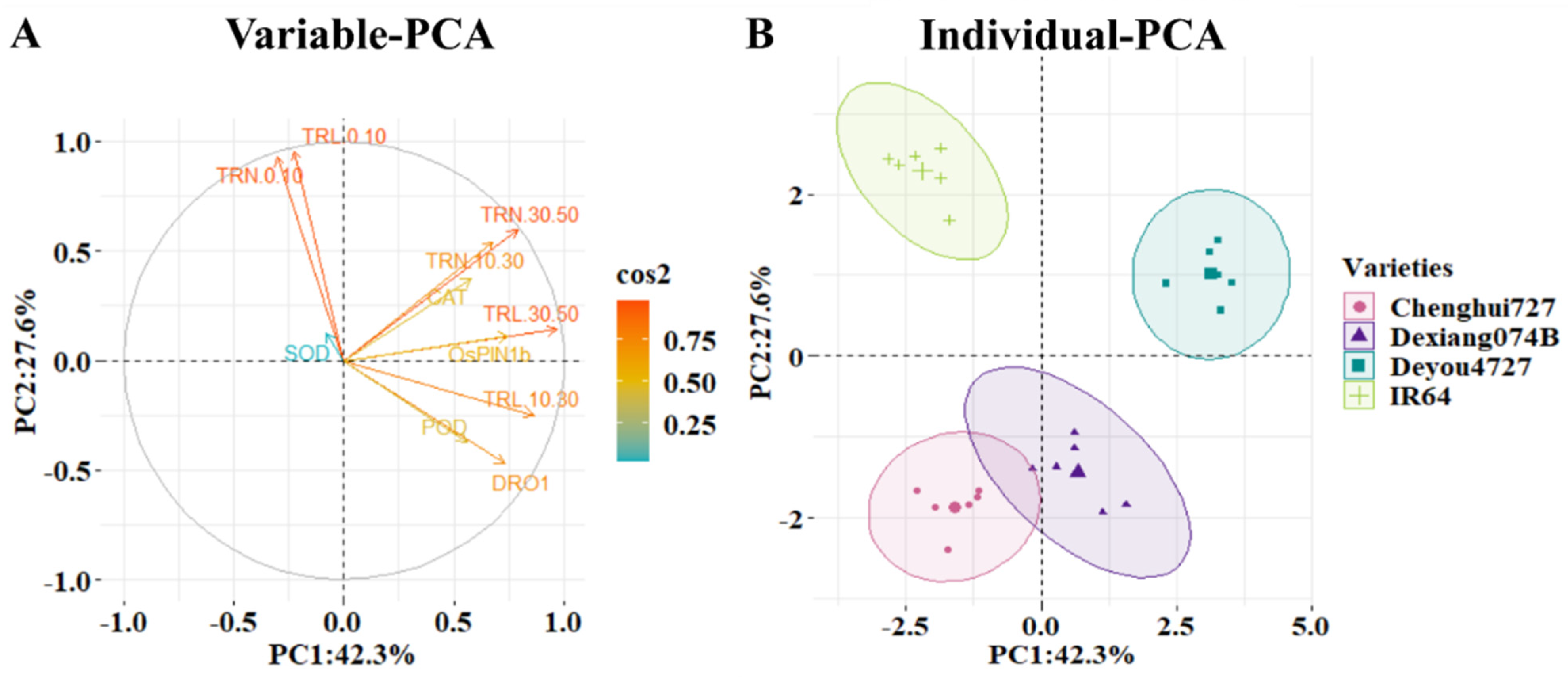
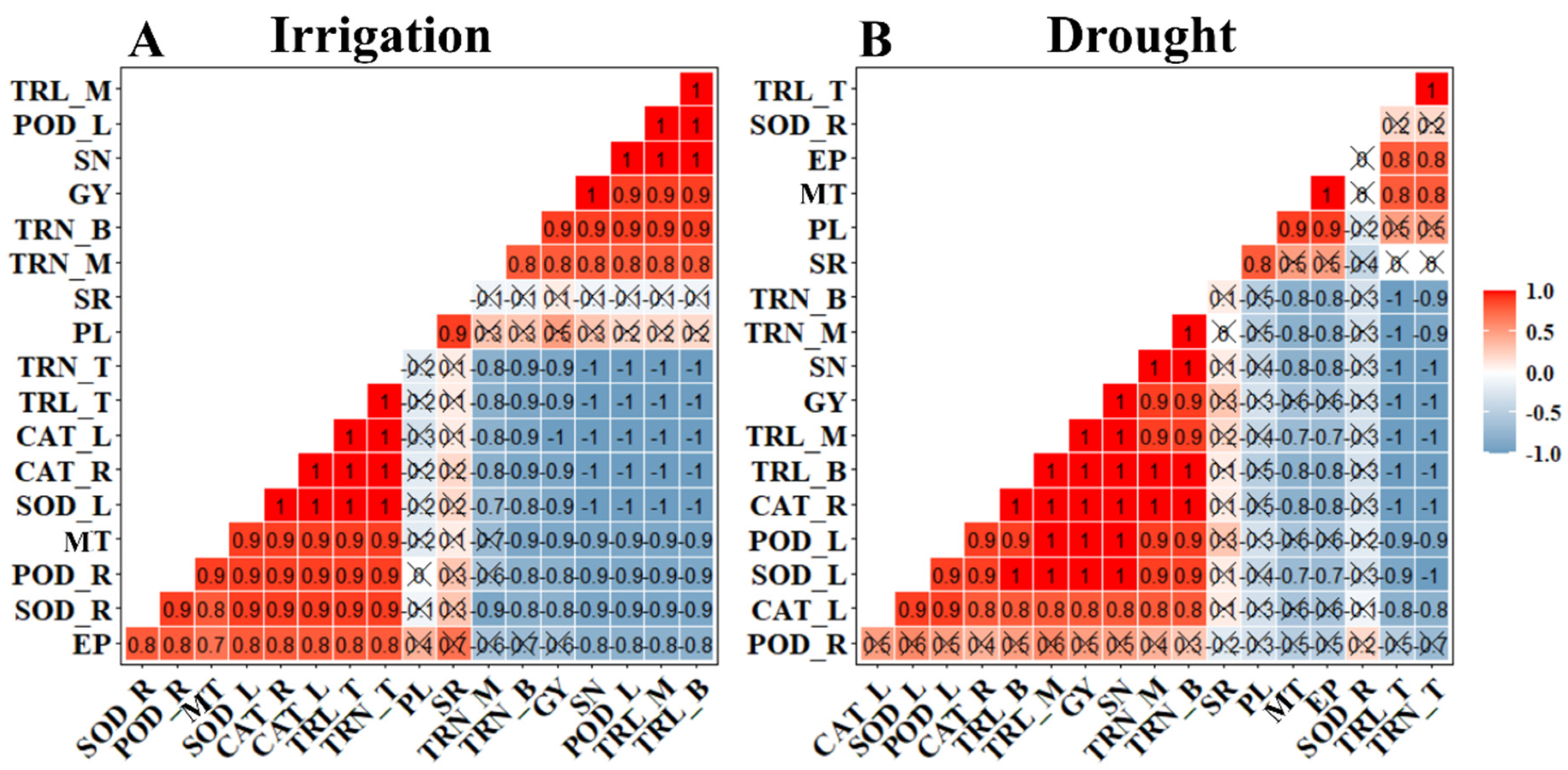
3.6. Multivariate and Pearson’s Correlation Coefficient Analysis of Root Traits, Responsible Genes, and Enzymatic System
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Samal, R.; Roy, P.S.; Sahoo, A.; Kar, M.K.; Patra, B.C.; Marndi, B.C.; Gundimeda, J.N.R. Morphological and molecular dissection of wild rices from eastern India suggests distinct speciation between O. rufipogon and O. nivara populations. Sci. Rep. 2018, 8, 2773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zafar, S.A.; Zaidi, S.S.-E.-A.; Gaba, Y.; Singla-Pareek, S.L.; Dhankher, O.P.; Li, X.; Mansoor, S.; Pareek, A. Engineering abiotic stress tolerance via CRISPR/Cas-mediated genome editing. J. Exp. Bot. 2020, 71, 470–479. [Google Scholar] [CrossRef] [PubMed]
- Bergeret, P. The future of food and agriculture: Trends and challenges [Note de lecture]. Bibliogr. Prospect. 2017, 29, 3. [Google Scholar]
- He, G.; Geng, C.; Zhao, Y.; Wang, J.; Jiang, S.; Zhu, Y.; Wang, Q.; Wang, L.; Mu, X. Food habit and climate change impacts on agricultural water security during the peak population period in China. Agric. Water Manag. 2021, 258, 107211. [Google Scholar] [CrossRef]
- Pérez-Escamilla, R. Can experience-based household food security scales help improve food security governance? Glob. Food Sec. 2012, 1, 120–125. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, M.S.; Wu, B.; Wang, H.; Kang, D. Field Screening of Rice Germplasm (Oryza sativa L. ssp. japonica) Based on Days to Flowering for Drought Escape. Plants 2020, 9, 609. [Google Scholar] [CrossRef]
- Ahmad, H.; Zafar, S.A.; Naeem, M.K.; Shokat, S.; Inam, S.; Naveed, A.S.; Xu, J.; Li, Z.; Ali, G.M.; Khan, M.R. Impact of pre-anthesis drought stress on physiology, yield-related traits and drought responsive genes in green super rice. bioRxiv 2021. [Google Scholar] [CrossRef]
- Serraj, R.; McNally, K.L.; Slamet-Loedin, I.; Kohli, A.; Haefele, S.M.; Atlin, G.; Kumar, A. Drought resistance improvement in rice: An integrated genetic and resource management strategy. Plant Prod. Sci. 2011, 14, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Wang, B.; Chen, L.; Li, P.; Cao, C. The different influences of drought stress at the flowering stage on rice physiological traits, grain yield, and quality. Sci. Rep. 2019, 9, 3742. [Google Scholar] [CrossRef] [Green Version]
- Eisenstein, M. Natural solutions for agricultural productivity. Nature 2020, 588, S58. [Google Scholar] [CrossRef]
- Punchkhon, C.; Plaimas, K.; Buaboocha, T.; Siangliw, J.L.; Toojinda, T.; Comai, L.; De Diego, N.; Spíchal, L.; Chadchawan, S. Drought-Tolerance Gene Identification Using Genome Comparison and Co-Expression Network Analysis of Chromosome Substitution Lines in Rice. Genes 2020, 11, 1197. [Google Scholar] [CrossRef] [PubMed]
- Pandey, V.; Shukla, A. Acclimation and tolerance strategies of rice under drought stress. Rice Sci. 2015, 22, 147–161. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, M.S.; Wu, B.; Wang, H.; Kang, D. Identification of Drought Tolerance on the Main Agronomic Traits for Rice (Oryza sativa L. ssp. japonica) Germplasm in China. Agronomy 2021, 11, 1740. [Google Scholar] [CrossRef]
- Yuan, L.-P. Hybrid rice achievements, development and prospect in China. J. Integr. Agric. 2015, 14, 197–205. [Google Scholar]
- Upadhyaya, H.; Panda, S.K. Drought stress responses and its management in rice. In Advances in Rice Research for Abiotic Stress Tolerance; Elsevier: Amsterdam, The Netherlands, 2019; pp. 177–200. [Google Scholar]
- Gupta, A.; Rico-Medina, A.; Caño-Delgado, A.I. The physiology of plant responses to drought. Science 2020, 368, 266–269. [Google Scholar] [CrossRef]
- Melandri, G.; AbdElgawad, H.; Riewe, D.; Hageman, J.A.; Asard, H.; Beemster, G.T.; Kadam, N.; Jagadish, K.; Altmann, T.; Ruyter-Spira, C. Biomarkers for grain yield stability in rice under drought stress. J. Exp. Bot. 2020, 71, 669–683. [Google Scholar] [CrossRef] [Green Version]
- Fukai, S.; Cooper, M. Development of drought-resistant cultivars using physiomorphological traits in rice. Field Crops Res. 1995, 40, 67–86. [Google Scholar] [CrossRef]
- Gowda, V.R.; Henry, A.; Yamauchi, A.; Shashidhar, H.; Serraj, R. Root biology and genetic improvement for drought avoidance in rice. Field Crops Res. 2011, 122, 1–13. [Google Scholar] [CrossRef]
- Uga, Y.; Okuno, K.; Yano, M. Dro1, a major QTL involved in deep rooting of rice under upland field conditions. J. Exp. Bot. 2011, 62, 2485–2494. [Google Scholar] [CrossRef] [Green Version]
- Kulkarni, M.; Soolanayakanahally, R.; Ogawa, S.; Uga, Y.; Selvaraj, M.G.; Kagale, S. Drought response in wheat: Key genes and regulatory mechanisms controlling root system architecture and transpiration efficiency. Front. Chem. 2017, 5, 106. [Google Scholar] [CrossRef] [Green Version]
- Mai, C.D.; Phung, N.T.; To, H.; Gonin, M.; Hoang, G.T.; Nguyen, K.L.; Do, V.N.; Courtois, B.; Gantet, P. Genes controlling root development in rice. Rice 2014, 7, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uga, Y.; Sugimoto, K.; Ogawa, S.; Rane, J.; Ishitani, M.; Hara, N.; Kitomi, Y.; Inukai, Y.; Ono, K.; Kanno, N. Control of root system architecture by DEEPER ROOTING 1 increases rice yield under drought conditions. Nat. Genet. 2013, 45, 1097–1102. [Google Scholar] [CrossRef]
- Yoo, S.-C.; Cho, S.-H.; Paek, N.-C. Rice WUSCHEL-related homeobox 3A (OsWOX3A) modulates auxin-transport gene expression in lateral root and root hair development. Plant Signal. Behav. 2013, 8, e25929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.; Singh, Y.; Xalaxo, S.; Verulkar, S.; Yadav, N.; Singh, S.; Singh, N.; Prasad, K.; Kondayya, K.; Rao, P.R. From QTL to variety-harnessing the benefits of QTLs for drought, flood and salt tolerance in mega rice varieties of India through a multi-institutional network. Plant Sci. 2016, 242, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Swamy, B.P.M.; Ahmed, H.U.; Henry, A.; Mauleon, R.; Dixit, S.; Vikram, P.; Tilatto, R.; Verulkar, S.B.; Perraju, P.; Mandal, N.P. Genetic, physiological, and gene expression analyses reveal that multiple QTL enhance yield of rice mega-variety IR64 under drought. PLoS ONE 2013, 8, e62795. [Google Scholar] [CrossRef]
- Xinqiao, Y.; Center, S.A.G.; Mingshou, L.; Center, S.A.G.; Hanwei, M.; Center, S.A.G. Huyou 2, a new hybrid combination of water-saving and drought resistance rice. Mol. Plant Breed. 2010, 8, 1177–1179. [Google Scholar]
- Yanling, H.; Shimei, W.; Dewen, Z.; Wanlin, W.; Qianjing, Y.; Qisheng, Z. The High Efficiency Water Saving of Drought Resistance ‘Lvhan No. 1’. Chin. Agric. Sci. Bull. 2010, 14. [Google Scholar]
- Zheng, W.; Ma, Z.; Zhao, M.; Xiao, M.; Zhao, J.; Wang, C.; Gao, H.; Bai, Y.; Wang, H.; Sui, G. Research and Development Strategies for Hybrid japonica Rice. Rice 2020, 13, 36. [Google Scholar] [CrossRef]
- Wang, X.; Samo, N.; Li, L.; Wang, M.; Qadir, M.; Jiang, K.; Qin, J.; Rasul, F.; Yang, G.; Hu, Y. Root Distribution and its impacts on the drought tolerance capacity of hybrid rice in the sichuan basin Area of China. Agronomy 2019, 9, 79. [Google Scholar] [CrossRef] [Green Version]
- Villa, J.E.; Henry, A.; Xie, F.; Serraj, R. Hybrid rice performance in environments of increasing drought severity. Field Crops Res. 2012, 125, 14–24. [Google Scholar] [CrossRef]
- Seo, J.S.; Joo, J.; Kim, M.J.; Kim, Y.K.; Nahm, B.H.; Song, S.I.; Cheong, J.J.; Lee, J.S.; Kim, J.K.; Choi, Y.D. OsbHLH148, a basic helix-loop-helix protein, interacts with OsJAZ proteins in a jasmonate signaling pathway leading to drought tolerance in rice. Plant J. 2011, 65, 907–921. [Google Scholar] [CrossRef] [PubMed]
- Vohník, M.; Pánek, M.; Fehrer, J.; Selosse, M.-A. Experimental evidence of ericoid mycorrhizal potential within Serendipitaceae (Sebacinales). Mycorrhiza 2016, 26, 831–846. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Zveushe, O.K.; Dong, F.; Ling, Q.; Chen, Y.; Sajid, S.; Zhou, L.; de Dios, V.R. Unraveling the effects of arbuscular mycorrhizal fungi on cadmium uptake and detoxification mechanisms in perennial ryegrass (Lolium perenne). Sci. Total Environ. 2021, 798, 149222. [Google Scholar] [CrossRef] [PubMed]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutases: I. Occurrence in higher plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Upadhyaya, A.; Sankhla, D.; Davis, T.D.; Sankhla, N.; Smith, B. Effect of paclobutrazol on the activities of some enzymes of activated oxygen metabolism and lipid peroxidation in senescing soybean leaves. J. Plant Physiol. 1985, 121, 453–461. [Google Scholar] [CrossRef]
- Aebi, H. [13] Catalase in vitro. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1984; Volume 105, pp. 121–126. [Google Scholar]
- Yang, G.; Nabi, F.; Sajid, S.; Kaleri, A.R.; Jakhar, A.M.; Cheng, L.; Raspor, M.; Muhammad, N.; Ma, J.; Hu, Y. Response of root development and nutrient uptake of two chinese cultivars of hybrid rice to nitrogen and phosphorus fertilization in Sichuan Province, China. Mol. Biol. Rep. 2021, 48, 8009–8021. [Google Scholar] [CrossRef]
- Duan, Y.; Jiang, Y.; Ye, S.; Karim, A.; Ling, Z.; He, Y.; Yang, S.; Luo, K. PtrWRKY73, a salicylic acid-inducible poplar WRKY transcription factor, is involved in disease resistance in Arabidopsis thaliana. Plant Cell Rep. 2015, 34, 831–841. [Google Scholar] [CrossRef] [Green Version]
- Lê, S.; Josse, J.; Mazet, F. Package FactoMineR. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Kassambara, A. ggcorrplot: Visualization of a Correlation Matrix using ‘ggplot2’. Comput. Softw. 2019, 3. [Google Scholar]
- Dar, M.H.; Waza, S.A.; Shukla, S.; Zaidi, N.W.; Nayak, S.; Hossain, M.; Kumar, A.; Ismail, A.M.; Singh, U.S. Drought tolerant rice for ensuring food security in Eastern India. Sustainability 2020, 12, 2214. [Google Scholar] [CrossRef] [Green Version]
- Rauf, S.; Al-Khayri, J.M.; Zaharieva, M.; Monneveux, P.; Khalil, F. Breeding strategies to enhance drought tolerance in crops. In Advances in Plant Breeding Strategies: Agronomic, Abiotic and Biotic Stress Traits; Springer: Berlin/Heidelberg, Germany, 2016; pp. 397–445. [Google Scholar]
- Fang, Y.; Du, Y.; Wang, J.; Wu, A.; Qiao, S.; Xu, B.; Zhang, S.; Siddique, K.H.; Chen, Y. Moderate drought stress affected root growth and grain yield in old, modern and newly released cultivars of winter wheat. Front. Plant Sci. 2017, 8, 672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.; Chung, Y.S.; Lee, E.; Tripathi, P.; Heo, S.; Kim, K.-H. Root response to drought stress in rice (Oryza sativa L.). Int. J. Mol. Sci. 2020, 21, 1513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verma, H.; Borah, J.; Sarma, R. Variability assessment for root and drought tolerance traits and genetic diversity analysis of rice germplasm using SSR markers. Sci. Rep. 2019, 9, 16513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henry, A.; Wehler, R.; Grondin, A.; Franke, R.; Quintana, M. Environmental and physiological effects on grouping of drought-tolerant and susceptible rice varieties related to rice (Oryza sativa) root hydraulics under drought. Ann. Bot. 2016, 118, 711–724. [Google Scholar] [CrossRef] [Green Version]
- Lynch, J.P.; Chimungu, J.G.; Brown, K.M. Root anatomical phenes associated with water acquisition from drying soil: Targets for crop improvement. J. Exp. Bot. 2014, 65, 6155–6166. [Google Scholar] [CrossRef] [Green Version]
- Zafar, S.A.; Hameed, A.; Ashraf, M.; Khan, A.S.; Li, X.; Siddique, K.H. Agronomic, physiological and molecular characterisation of rice mutants revealed the key role of reactive oxygen species and catalase in high-temperature stress tolerance. Funct. Plant Biol. 2020, 47, 440–453. [Google Scholar] [CrossRef]
- Maurino, V.G.; Flügge, U.-I. Experimental systems to assess the effects of reactive oxygen species in plant tissues. Plant Signal. Behav. 2008, 3, 923–928. [Google Scholar] [CrossRef] [Green Version]
- Alscher, R.G.; Erturk, N.; Heath, L.S. Role of superoxide dismutases (SODs) in controlling oxidative stress in plants. J. Exp. Bot. 2002, 53, 1331–1341. [Google Scholar] [CrossRef]
- ElSayed, A.I.; El-Hamahmy, M.A.; Rafudeen, M.S.; Mohamed, A.H.; Omar, A.A. The impact of drought stress on antioxidant responses and accumulation of flavonolignans in milk thistle (Silybum marianum (l.) gaertn). Plants 2019, 8, 611. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, C.B.; Rouina, B.B.; Sensoy, S.; Boukhris, M.; Abdallah, F.B. Changes in gas exchange, proline accumulation and antioxidative enzyme activities in three olive cultivars under contrasting water availability regimes. Environ. Exp. Bot. 2009, 67, 345–352. [Google Scholar] [CrossRef]
- Reddy, A.R.; Chaitanya, K.; Jutur, P.; Sumithra, K. Differential antioxidative responses to water stress among five mulberry (Morus alba L.) cultivars. Environ. Exp. Bot. 2004, 52, 33–42. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B. Reactive species and antioxidants. Redox biology is a fundamental theme of aerobic life. Plant Physiol. 2006, 141, 312–322. [Google Scholar] [CrossRef] [Green Version]
- Kantar, M.; Lucas, S.J.; Budak, H. Drought stress: Molecular genetics and genomics approaches. Adv. Bot. Res. 2011, 57, 445–493. [Google Scholar]
- Kitomi, Y.; Hanzawa, E.; Kuya, N.; Inoue, H.; Hara, N.; Kawai, S.; Kanno, N.; Endo, M.; Sugimoto, K.; Yamazaki, T. Root angle modifications by the DRO1 homolog improve rice yields in saline paddy fields. Proc. Nati. Acad. Sci. USA 2020, 117, 21242–21250. [Google Scholar] [CrossRef]
- Sun, H.; Tao, J.; Bi, Y.; Hou, M.; Lou, J.; Chen, X.; Zhang, X.; Luo, L.; Xie, X.; Yoneyama, K. OsPIN1b is involved in rice seminal root elongation by regulating root apical meristem activity in response to low nitrogen and phosphate. Sci. Rep. 2018, 8, 13014. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Varieties | Grain Yield (g/per Plant) | MT | PL | EP | SN | SSR (%) | |
|---|---|---|---|---|---|---|---|
| Irrigation | IR64 | 25 ± 1.50 b | 23.3 ± 1.11 a | 23.8 ± 1.02 a | 15.3 ± 1.41 a | 85.7 ± 1.21 b | 86.7 ± 2.29 a |
| Deyou4727 | 33 ± 1.58 a | 17.7 ± 1.16 b | 24.2 ± 1.08 a | 12.3 ± 1.26 b | 148.0 ± 2.17 a | 86.3 ± 1.51 a | |
| Drought | IR64 | 20 ± 1.16 b | 20.3 ± 1.28 a | 22.4 ± 1.45 a | 13.0 ± 1.03 a | 55.7 ± 1.59 b | 75.1 ± 1.11 a |
| Deyou4727 | 30 ± 1.62 a | 17.2 ± 1.29 b | 21.1 ± 1.15 a | 10.7 ± 0.97 b | 98.3 ± 1.17 a | 75.3 ± 1.27 a |
| Treatments | Varieties | CAT | POD | SOD | |||
|---|---|---|---|---|---|---|---|
| Leaf | Root | Leaf | Root | Leaf | Root | ||
| Irrigation | IR64 | 635 ± 17 a | 554 ± 17 a | 5 ± 1.2 b | 14 ± 1.3 a | 67 ± 4.6 a | 49 ± 3.8 a |
| Deyou4727 | 560 ± 16 b | 452 ± 14 b | 7 ± 1.3 b | 9 ± 1.1 b | 57 ± 3.4 b | 41 ± 4.6 b | |
| Chenghui727 | 557 ± 15 b | 435 ± 12 b | 6 ± 1.1 b | 8 ± 1.3 b | 40 ± 3.5 c | 40 ± 4.2 b | |
| Dexiang074 B | 638 ± 18 a | 562 ± 15 a | 10 ± 1.2 a | 10 ± 1.1 ab | 57 ± 3.3 b | 53 ± 3.6 a | |
| Drought | IR64 | 756 ± 16 b | 894 ± 19 b | 17 ± 1.1 b | 29 ± 3.2 b | 57 ± 4.3 b | 75 ± 4.2 a |
| Deyou4727 | 800 ± 17 a | 977 ± 16 a | 20 ± 1.8 a | 33 ± 3.4 a | 70 ± 5.2 a | 74 ± 4.3 a | |
| Chenghui727 | 787 ± 14 ab | 675 ± 15 c | 18 ± 2.3 ab | 31 ± 3.2 ab | 78 ± 5.3 a | 74 ± 4.6 a | |
| Dexiang074 B | 820 ± 15 a | 987 ± 18 a | 22 ± 2.9 a | 36 ± 3.2 a | 61 ± 4.6 b | 75 ± 4.7 a | |
| Material | UTR | 5′UTR | EXON 3 | EXON 3 | EXON4 | EXON4 | 3′UTR |
|---|---|---|---|---|---|---|---|
| cDNA | 27 | 150 | 617 | 752 | 943 | 962 | 1133–1134 |
| gDNA | 27 | 150 | 2096 | 2231 | 2513 | 2532 | 2909–2910 |
| Seq (+) | A/G | C/T | T/C | A/C | A/- | C/A | CT/- |
| IR64 | G | T | C | A | - | C | - |
| Deyou4727 | A | T | C | C | A | C | - |
| Chenghui727 | A | T | C | C | A | C | - |
| Dexiang074B | A | T | C | C | A | C | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nabi, F.; Sajid, S.; Yang, G.; Kaleri, A.R.; Zheng, Y.; Qin, J.; Wang, X.; Jiang, K.; Peng, Y.; Hu, Y. The Adenine at the 4th Exon of the DRO1 Gene Provides Drought-Tolerance Capacity to Hybrid Rice Deyou4727 and Its Maintainer Line Dexiang074B. Agronomy 2022, 12, 752. https://doi.org/10.3390/agronomy12030752
Nabi F, Sajid S, Yang G, Kaleri AR, Zheng Y, Qin J, Wang X, Jiang K, Peng Y, Hu Y. The Adenine at the 4th Exon of the DRO1 Gene Provides Drought-Tolerance Capacity to Hybrid Rice Deyou4727 and Its Maintainer Line Dexiang074B. Agronomy. 2022; 12(3):752. https://doi.org/10.3390/agronomy12030752
Chicago/Turabian StyleNabi, Farhan, Sumbal Sajid, Guotao Yang, Abdul Rasheed Kaleri, Yating Zheng, Jian Qin, Xuechun Wang, Kaifeng Jiang, Youlin Peng, and Yungao Hu. 2022. "The Adenine at the 4th Exon of the DRO1 Gene Provides Drought-Tolerance Capacity to Hybrid Rice Deyou4727 and Its Maintainer Line Dexiang074B" Agronomy 12, no. 3: 752. https://doi.org/10.3390/agronomy12030752