
Effect of Fungicides on Bayberry Decline Disease by Modulating Rhizosphere Soil Properties, Microflora, and Metabolites


 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Vegetative Growth Parameters Measurement
2.3. Fruit Economic Characters Measurement
2.4. Soil Property Measurements
2.5. Soil Genome Sequencing
2.6. GC-MS Metabolomics Analysis
2.7. Statistical Analysis
3. Results and Discussion
3.1. Effects of Prochloraz on Vegetative Growth and Fruit Quality
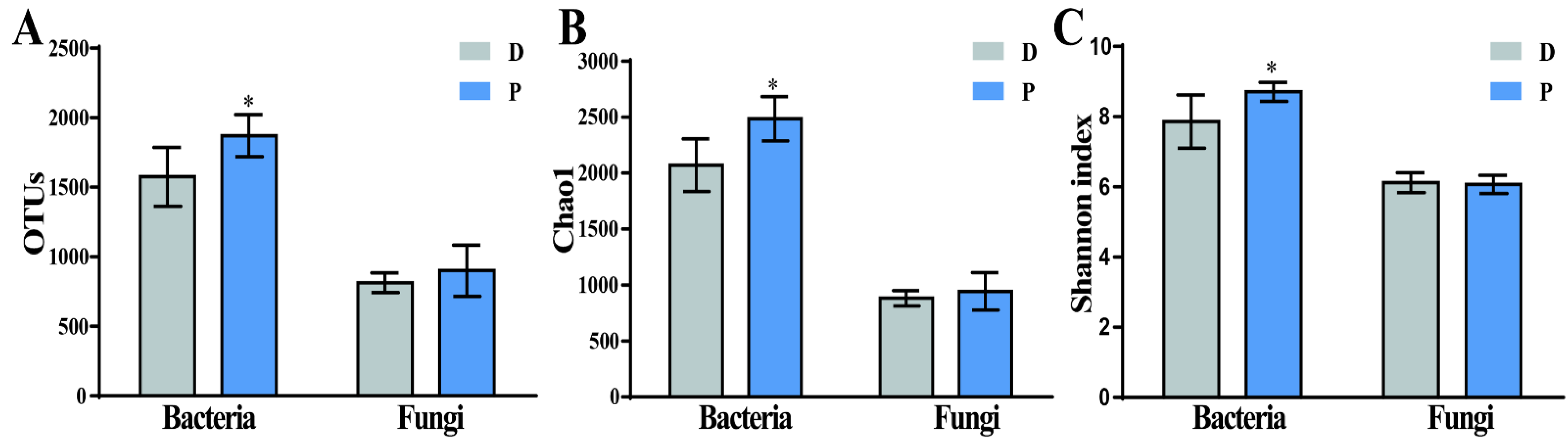
3.2. The Effect of Prochloraz in Microbial Community Diversity
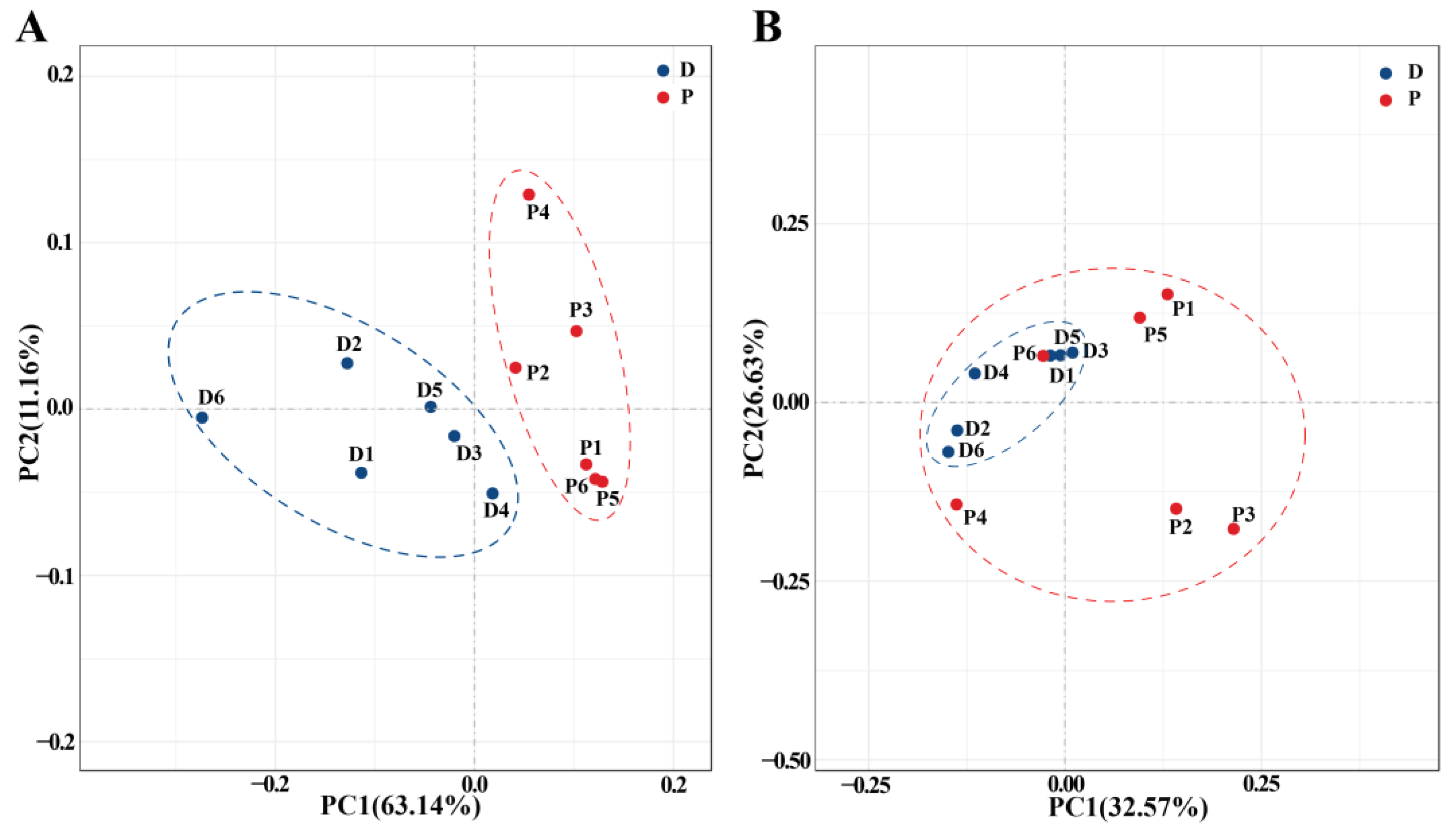
3.3. Effect of Prochloraz on Soil Microbial Community Structure
3.4. Effect of Prochloraz on the Soil Nutrient Status
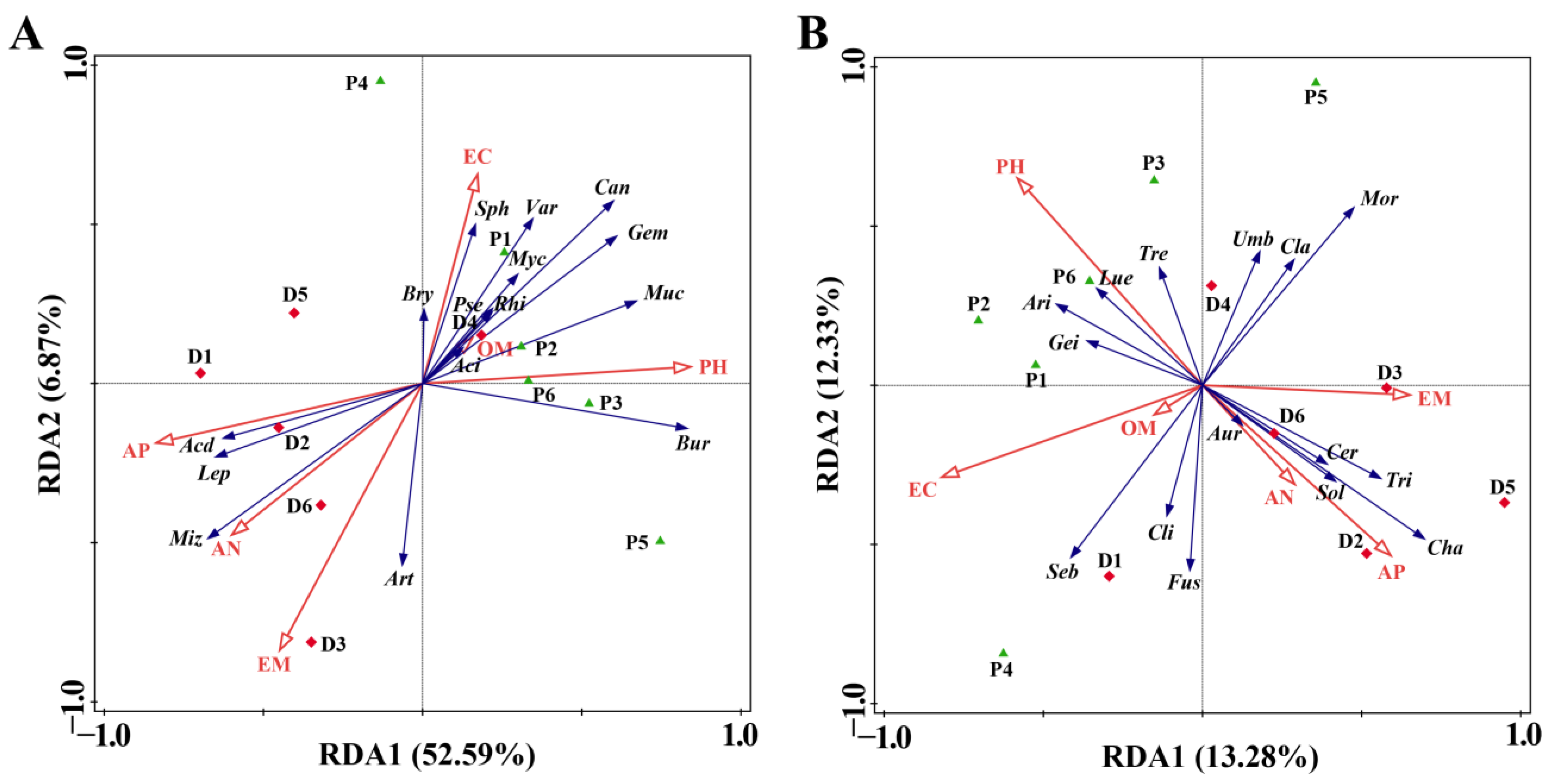
3.5. Effect of Prochloraz on RDA
3.6. Effect of Prochloraz on Rhizosphere Soil Metabolomics
3.7. Effect of Prochloraz on the Metabolic Pathways
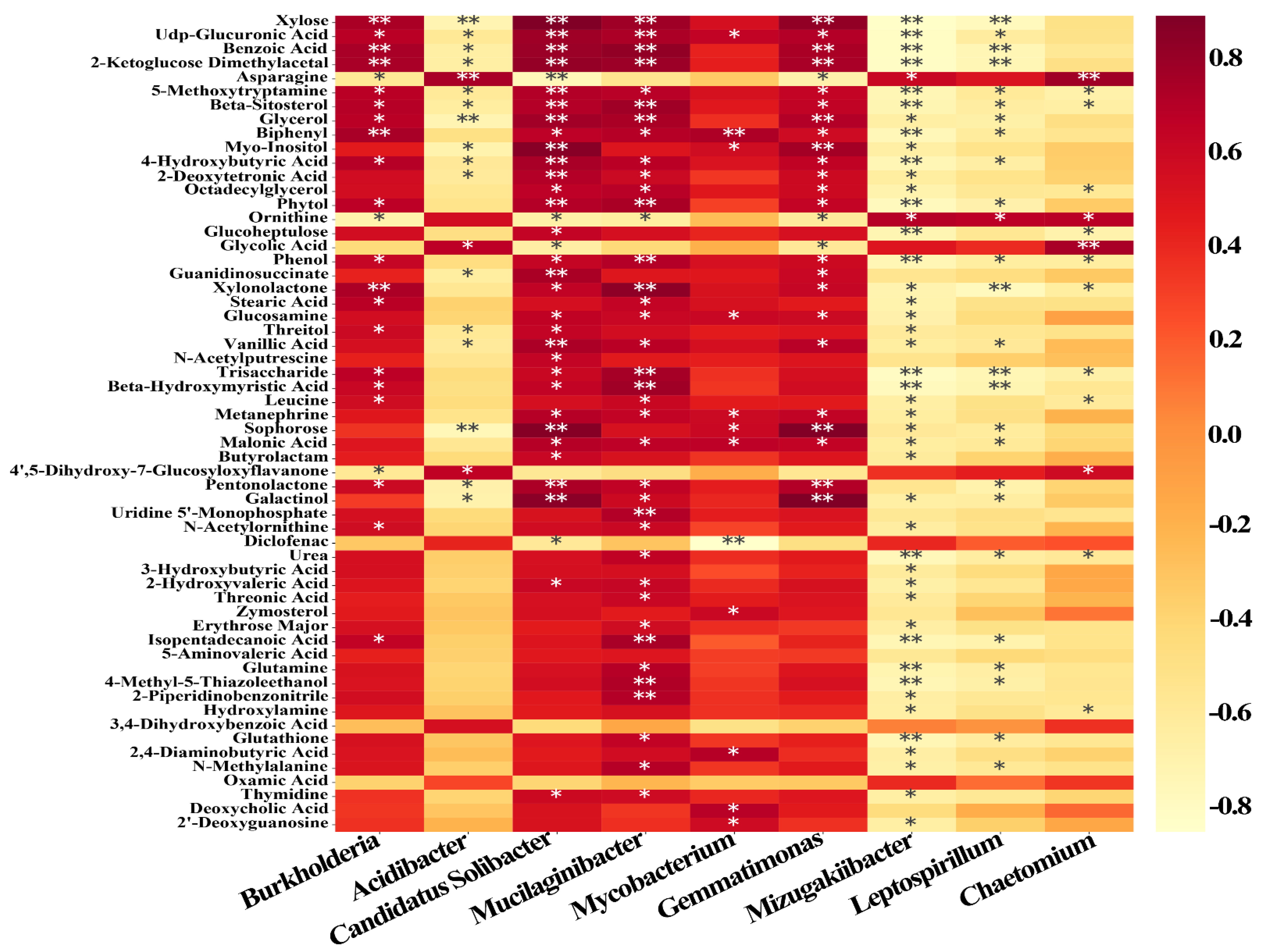
3.8. Correlations of Soil Microbial Communities with Plant Secondary Metabolites
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ren, H.Y.; Yu, H.Y.; Zhang, S.W.; Liang, S.M.; Zheng, X.L.; Zhang, S.J.; Yao, P.; Zheng, H.K.; Qi, X.J. Genome sequencing provides insights into the evolution and antioxidant activity of Chinese bayberry. BMC Genom. 2019, 20, 458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, C.; Huang, H.Z.; Xu, C.J.; Li, X.; Chen, K.S. Biological activities of extracts from Chinese bayberry (Myrica rubra Sieb. et Zucc.): A review. Plant Foods Hum. Nutr. 2013, 68, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Erickson, D.L.; Hamrick, J.L. Genetic and clonal diversity Myrica cerifera for along a spatiotemporal island chronosequence. Heredity 2003, 90, 25–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, H.Y.; Zheng, X.L.; Zhang, S.W.; Liang, S.M.; Wu, C.W.; Wang, K.Q.; Yu, Z.P.; Qi, X.J. The symptoms and nutritional status of decline diseased bayberry. Zhejiang Agri. Sci. 2020, 61, 2043–2048. (In Chinese) [Google Scholar]
- Ren, H.Y.; Wang, H.Y.; Qi, X.J.; Yu, Z.P.; Zheng, X.L.; Zhang, S.W.; Wang, Z.S.; Zhang, M.C.; Ahmed, T.; Li, B. The damage caused by decline disease in bayberry plants through changes in soil properties, rhizosphere microbial community structure and metabolites. Plants 2021, 10, 2083. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.Y.; Wang, J.; Qi, X.J.; Yu, Z.P.; Zhang, Q. The rejuvenation effect of 10 fungicides on decline disease bayberry. Zhejiang Agri. Sci. 2021, 62, 27–30. (In Chinese) [Google Scholar]
- Zhang, R.; Xu, Q.R.; Zhang, Y.C.; Zhu, F.X. Baseline sensitivity and toxic actions of prochloraz to Sclerotinias clerotiorum. Plant Dis. 2018, 102, 2149–2157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frąc, M.; Lipiec, J.; Usowicz, B.; Oszust, K.; Brzezińska, M. Structural and functional microbial diversity of sandy soil under cropland and grassland. PeerJ 2020, 8, e9501. [Google Scholar] [CrossRef]
- Zhang, Y.; Tian, C.; Xiao, J.L.; Wei, L.; Tian, Y.; Liang, Z.H. Soil inoculation of Trichoderma asperellum M45a regulates rhizosphere microbes and triggers watermelon resistance to Fusarium wilt. AMB Express 2020, 10, 189. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Zhou, T.; Zhu, L.; Juhasz, A.; Du, Z.; Li, B.; Wang, J.; Wang, J.; Sun, Y. Response of soil microbes after direct contact with pyraclostrobin in fluvo-aquic soil. Environ. Pollut. 2019, 255 Pt 1, 113164. [Google Scholar] [CrossRef]
- Wang, L.G.; Shu, N.L.; Sun, K.G. The process of soybean continuous cropping obstacle and control teachnology. Soil Fert. Sci. Chin. 2001, 5, 3–8. [Google Scholar]
- Lucie, M.; Jakub, R.; Marketa, P.; Tomas, M.; Ondrej, U. Effects of secondary plant metabolites on microbial populations: Changes in community structure and metabolic activity in contaminated environments. Int. J. Mol. Sci. 2016, 17, 1205. [Google Scholar]
- Sogi, D.S.; Siddiq, M.; Dolan, K.D. Total phenolics, carotenoids and antioxidant properties of tommy atkin mango cubes as affected by drying techniques. LWT-Food Sci. Technol. 2015, 62, 564–568. [Google Scholar] [CrossRef]
- Jiang, Y.; Nie, W.J. Chemical properties in fruits of mulberry species from the Xinjiang province of China. Food Chem. 2015, 174, 460–466. [Google Scholar] [CrossRef]
- Yin, H.; Zheng, H.; Zhang, B.; Tariq, A.; Lv, G.; Zeng, F.; Graciano, C. Stoichiometry of C:N:P in the roots of alhagi sparsifolia is more sensitive to soil nutrients than aboveground organs. Front. Plant Sci. 2021, 12, 698961. [Google Scholar] [CrossRef]
- Wu, L.Y.; Wen, C.Q.; Qin, Y.J.; Yin, H.Q.; Tu, Q.C.; Nostrand, J.D.V.; Yuan, T.; Yuan, M.T.; Deng, Y.; Zhou, J.Z. Phasing amplicon sequencing on Illumina Miseq for robust environmental microbial community analysis. BMC Microbio. 2015, 15, 125. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, P.K.; Chandra, J.; Retuerto, M.; Sikaroodi, M.; Brown, R.E.; Jurevic, R.; Salata, R.A.; Lederman, M.M.; Gillevet, P.M.; Ghannoum, M.A. Oral mycobiome analysis of HIV-infected patients: Identification of pichia as an antagonist of opportunistic fungi. PLoS Pathog. 2014, 10, e1003996. [Google Scholar] [CrossRef]
- Reyon, D.; Tsai, S.Q.; Khayter, C.; Foden, J.A.; Sander, J.D.; Joung, J.K. FLASH assembly of TALENs for high-throughput genome editing. Nat. Biotech. 2012, 30, 460–465. [Google Scholar] [CrossRef]
- Chao, J.; Bunge, J. Estimatin the number of species in a stochastic abundance model. Biometrics 2002, 58, 531–539. [Google Scholar] [CrossRef]
- Hill, T.C.; Walsh, K.A.; Harris, J.A.; Moffett, B.F. Using ecological diversity measures with bacterial communities. FEMS Microbiol. Ecol. 2003, 43, 1–11. [Google Scholar] [CrossRef]
- Meng, T.; Ren, G.; Wang, G.; Ma, Y. Impacts on soil microbial characteristics and their restorability with different soil disinfestation approaches in intensively cropped greenhouse soils. Appl. Microbiol. Biotechnol. 2019, 103, 6369–6383. [Google Scholar] [CrossRef] [PubMed]
- Bergsveinson, J.; Perry, B.J.; Sheedy, C.; Braul, L.; Reedyk, S.; Gossen, B.D.; Yost, C.K. Identifying the core bacterial and fungal communities within four agricultural biobeds used for the treatment of pesticide rinsates. J. Appl. Microbiol. 2018, 125, 1333–1342. [Google Scholar] [CrossRef] [PubMed]
- Baćmaga, M.; Wyszkowska, J.; Kucharski, J. Response of soil microorganisms and enzymes to the foliar application of Helicur 250 EW fungicide on Horderum vulgare L. Chemosphere 2020, 242, 125163. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.Y.; Wang, H.Y.; Yu, Z.P.; Zhang, S.W.; Qi, X.J.; Sun, L.; Wang, Z.S.; Zhang, M.C.; Ahmed, T.; Li, B. Effect of two kinds of fertilizers on growth and rhizosphere soil properties of bayberry with decline disease. Plants 2021, 10, 2386. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Zhou, J.J.; Wang, E.T.; Chen, Q.; Xu, J.; Sun, J.G. Multiphasic characterization of a plant growth promoting bacterial strain, Burkholderia sp 7016 and its effect on tomato growth in the field. J. Int. Agric. 2015, 14, 1855–1863. [Google Scholar] [CrossRef] [Green Version]
- Maqsood, A.; Shahid, M.; Hussain, S.; Mahmood, F.; Basit, F. Root colonizing Burkholderia sp. AQ12 enhanced rice growth and upregulated tillering-responsive genes in rice. Appl. Soil. Ecol. 2020, 157, 103769. [Google Scholar] [CrossRef]
- Santísima-Trinidad, A.B.L.; Montiel-Rozas, M.D.M.; Diéz-Rojo, M.A.; Pascual, J.A.; Ros, M. Impact of foliar fungicides on target and non-target soil microbial communities in cucumber crops. Ecotoxicol. Environ. Saf. 2018, 30, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Q.; Su, Y.; Ahmed, T.; Ren, H.Y.; Javed, M.R.; Yao, Y.L.; An, Q.L.; Yan, J.L.; Li, B. Effects of different organic fertilizers on improving soil from newly reclaimed land to crop soil. Agriculture 2021, 11, 560. [Google Scholar] [CrossRef]
- Pierattini, E.C.; Francini, A.; Huber, C.; Sebastiani, L.; Schröder, P. Poplar and diclofenac pollution: A focus on physiology, oxidative stress and uptake in plant organs. Sci. Total Environ. 2018, 15, 944–952. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.S.; Vivanco, J.M.; Manter, D.K. Nitrogen fertilizer rate affects root exudation, the rhizosphere microbiome and nitrogen-use-efficiency of maize. Appl. Soil Ecolo. 2016, 107, 324–333. [Google Scholar] [CrossRef] [Green Version]
- Zhou, W.X.; Branch, W.D.; Gilliam, L.; Marsh, J.A. Phytosterol composition of arachis hypogaea seeds from different maturity classes. Molecules 2018, 24, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scholz, B.; Guth, S.; Engel, K.H.; Steinberg, P. Phytosterol oxidation products in enriched foods: Occurrence, exposure, and biological effects. Mol. Nutr. Food Res. 2015, 59, 1339–1352. [Google Scholar] [CrossRef] [PubMed]
- Xue, H.; Chen, X.; Li, G. Involvement of phospholipids signaling in plant growth and hormone effects. Curr. Opin. Plant Biol. 2007, 10, 483–489. [Google Scholar] [CrossRef]
- Patra, B.; Ray, S.; Richter, A.; Majumder, A.L. Enhanced salt tolerance of transgenic tobacco plants by co-expression of PcINO1 and McIMT1 is accompanied by increased level of myo-inositol and methylated inositol. Protoplasma 2010, 245, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, R.; Mukherjee, A.; Bandyopadhyay, S.; Mukherjee, S.; Sengupta, S.; Ray, S. Selective manipulation of the inositol metabolic pathway for induction of salt-tolerance in indica rice variety. Sci. Rep. 2019, 9, 5358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghimire, B.K.; Ghimire, B.; Yu, C.Y.; Chung, I.M. Allelopathic and autotoxic effects of medicago sativa-derived allelochemicals. Plants 2019, 8, 233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shibl, A.A.; Isaac, A.; Ochsenkuhn, M.A.; Cardenas, A.; Fei, C.; Behringer, G.; Arnoux, M.; Drou, N.; Santos, M.P.; Gunsalus, K.C.; et al. Diatom modulation of select bacteria through use of two unique secondary metabolites. Proc. Natl. Acad. Sci. USA 2020, 117, 27445–27455. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.Y.; Shen, J.P.; Sun, G.X.; Wang, B.; Ji, R.; Zhao, L.J. Foliar application of SiO2 nanoparticles alters soil metabolite profiles and microbial community composition in the pakchoi (Brassica chinensis L.) rhizosphere grown in contaminated mine soil. Environ. Sci. Technol. 2020, 54, 13137–13146. [Google Scholar] [CrossRef]
- Zhao, L.J.; Zhang, H.L.; Jason, C.W.; Chen, X.Q.; Li, H.B.; Qua, X.L.; Ji, R. Metabolomics reveals that engineered nanomaterial exposure in soil alters both soil rhizosphere metabolite profiles and maize metabolic pathways. Environ. Sci. Nano 2019, 6, 1716–1727. [Google Scholar] [CrossRef]
- Zhang, Y.; Tao, Y.; Zhang, H.; Wang, L.; Sun, G.Q.; Sun, X.; Erinle, K.O.; Feng, C.C.; Song, Q.X.; Li, M. Effect of di-n-butyl phthalate on root physiology and rhizosphere microbial community of cucumber seedings. J. Hazard. Mater. 2015, 289, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.C.; Wu, B.D.; Jiang, K. Allelopathic effects of Canada goldenrod leaf extracts on the seed germination and seedling growth of lettuce reinforced under salt stress. Ecotoxicology 2019, 28, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, K.; Dubey, V.K. Fresh insights into the pyrimidine metabolism in the trypanosomatids. Parasit. Vectors 2018, 11, 87. [Google Scholar] [CrossRef] [PubMed]
- Coelho, A.I.; Berry, G.T.; Rubio-Gozalbo, M.E. Galactose metabolism and health. Curr. Opin. Clin. Nutr. Metab. Care 2015, 18, 422–427. [Google Scholar] [CrossRef]
- Appleton, J. Arginine: Clinical potential of a semi-essential amino acid. Altern. Med. Rev. 2002, 7, 512–522. [Google Scholar] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Parameters | ||
|---|---|---|---|
| Length of stem/(mm) | Leaf length/(mm) | ||
| D | 13.21 ± 0.54 | D | 99.51 ± 3.85 |
| P | 33.40 ± 1.45 * | P | 110.57 ± 1.24 * |
| Leaf width/(mm) | Leaf thickness/(mm) | ||
| D | 27.08 ± 0.41 | D | 4.06 ± 0.92 |
| P | 33.12 ± 0.08 * | P | 4.65 ± 0.22 * |
| Rate of photosynthesis/ (mg CO2·10 cm−2·h−1) | Chlorophyll/(SPAD) | ||
| D | 1.87 ± 0.28 | D | 44.62 ± 0.65 |
| P | 3.10 ± 0.17 * | P | 51.98 ± 0.50 * |
| Parameters | Parameters | ||
|---|---|---|---|
| Single fruit weight/g | Soluble solids/% | ||
| D | 11.13 ± 0.51 | D | 8.95 ± 0.25 |
| P | 15.07 ± 0.44 * | P | 12.07 ± 1.11 * |
| Titratable acid/% | Vitamin C/(mg/100 g) | ||
| D | 1.28 ± 0.12 | D | 8.73 ± 0.17 |
| P | 0.91 ± 0.03 # | P | 9.87 ± 0.79 * |
| Parameters | D | P | Parameters | D | P |
|---|---|---|---|---|---|
| pH | 4.69 ± 0.39 | 5.38 ± 0.45 * | AP (mg/kg) | 30.78 ± 0.50 | 15.98 ± 1.72 # |
| OM (%) | 2.80 ± 0.12 | 3.03 ± 0.24 * | EC (mg/kg) | 348.33 ± 9.49 | 376.20 ± 11.74 * |
| AHN (mg/kg) | 117.90 ± 4.58 | 108.27 ± 3.84 # | EM (mg/kg) | 35.54 ± 1.12 | 29.62 ± 1.41 # |
| Soil Environment | Contribution at Genus Level (%) | Soil Environment | Contribution at Genus Level (%) | ||
|---|---|---|---|---|---|
| Bacteria | Fungi | Bacteria | Fungi | ||
| pH | 38.4 | 10.2 | AP | 37.7 | 10.0 |
| OM | 4.0 | 2.0 | EC | 6.6 | 10.8 |
| AHN | 21.0 | 5.0 | EM | 15.5 | 6.6 |
| Metabolite Name | Relative Content | Metabolite Name | Relative Content |
|---|---|---|---|
| Diclofenac | 3,4-Dihydroxybenzoic Acid | ||
| D | 130.89 ± 99.65 | D | 5.52 ± 3.18 |
| P | 6.27 # ± 0.89 | P | 2.13 # ± 0.40 |
| Asparagine | Glycolic Acid | ||
| D | 1.28 ± 0.20 | D | 66.58 ± 12.30 |
| P | 0.74 # ± 0.04 | P | 45.60 # ± 1.26 |
| Oxamic Acid | Ornithine | ||
| D | 44.11 ± 14.76 | D | 0.40 ± 0.14 |
| P | 27.90 # ± 5.43 | P | 0.15 # ± 0.04 |
| 4′,5-Dihydroxy-7-Glucosyloxyflavanone | Thymidine | ||
| D | 10.72 ± 5.57 | D | 1.27 ± 0.23 |
| P | 2.88 # ± 1.24 | P | 1.58 * ± 0.20 |
| Beta-Hydroxymyristic Acid | Isopentadecanoic Acid | ||
| D | 3.71 ± 0.74 | D | 0.38 ± 0.11 |
| P | 5.21 * ± 0.68 | P | 0.56 * ± 0.11 |
| Erythrose Major | 5-Aminovaleric Acid | ||
| D | 1.65 ± 0.40 | D | 0.33 ± 0.07 |
| P | 2.23 * ± 0.33 | P | 0.45 * ± 0.08 |
| 2,4-Diaminobutyric Acid | 2’-Deoxyguanosine | ||
| D | 0.93 ± 0.201 | D | 0.25 ± 0.05 |
| P | 1.17 * ± 0.10 | P | 0.31 * ± 0.03 |
| Glucoheptulose | Hydroxylamine | ||
| D | 3.90 ± 1.51 | D | 20.18 ± 7.32 |
| P | 6.54 * ± 0.38 | P | 28.77 * ± 3.39 |
| Phenol | Uridine 5′-Monophosphate | ||
| D | 18.94 ± 6.24 | D | 0.23 ± 0.09 |
| P | 33.98 * ± 6.72 | P | 0.57 * ± 0.25 |
| Octadecylglycerol | Leucine | ||
| D | 0.25 ± 0.06 | D | 1.75 ± 0.40 |
| P | 0.38 * ± 0.04 | P | 2.43 * ± 0.22 |
| Trisaccharide | Urea | ||
| D | 1022.82 ± 217.23 | D | 150.49 ± 36.48 |
| P | 1393.56 * ± 117.51 | P | 198.22 * ± 17.28 |
| Glutathione | 2-Piperidinobenzonitrile | ||
| D | 0.97 ± 0.27 | D | 0.37 ± 0.11 |
| P | 1.29 * ± 0.14 | P | 0.54 * ± 0.12 |
| N-Methylalanine | Glutamine | ||
| D | 2.55 ± 0.66 | D | 3.52 ± 0.95 |
| P | 3.48 * ± 0.62 | P | 4.71 * ± 0.55 |
| 4-Methyl-5-Thiazoleethanol | Threonic Acid | ||
| D | 0.28 ± 0.07 | D | 0.86 ± 0.14 |
| W | 0.37 * ± 0.05 | P | 1.20 * ± 0.26 |
| Xylonolactone | Udp-Glucuronic Acid | ||
| D | 1.28 ± 0.14 | D | 0.32 ± 0.05 |
| W | 1.56 * ± 0.12 | P | 0.53 * ± 0.05 |
| Beta-Sitostero | Biphenyl | ||
| D | l1.64 ± 0.42 | D | 0.59 ± 0.14 |
| P | 3.50 * ± 0.64 | P | 1.07 * ± 0.17 |
| 5-Methoxytryptamine | Stearic Acid | ||
| D | 38.13 ± 12.79 | D | 1.80 ± 0.53 |
| P | 83.51 * ± 11.83 | P | 3.20 * ± 0.69 |
| Sophorose | Galactinol | ||
| D | 8.91 ± 1.63 | D | 1.18 ± 0.40 |
| P | 14.53 * ± 3.52 | P | 2.22 * ± 0.69 |
| Zymosterol | Deoxycholic Acid | ||
| D | 0.46 ± 0.23 | D | 0.45 ± 0.21 |
| P | 0.73 * ± 0.08 | P | 0.67 * ± 0.04 |
| Malonic Acid | Vanillic Acid | ||
| D | 0.21 ± 0.03 | D | 3.69 ± 0.83 |
| P | 0.25 * ± 0.01 | P | 5.24 * ± 0.55 |
| Metanephrine | N-Acetylputrescine | ||
| D | 0.79 ± 0.35 | D | 4.19 ± 1.06 |
| P | 1.42 * ± 0.23 | P | 7.34 * ± 1.77 |
| Butyrolactam | Phytol | ||
| D | 4.75 ± 0.75 | D | 1.66 ± 0.45 |
| P | 6.70 * ± 1.18 | P | 2.79 * ± 0.47 |
| 2-Deoxytetronic Acid | N-Acetylornithine | ||
| D | 2.52 ± 0.63 | D | 2.52 ± 0.72 |
| P | 6.04 * ± 1.79 | P | 4.35 * ± 1.26 |
| 3-Hydroxybutyric Acid | Benzoic Acid | ||
| D | 63.10 ± 29.70 | D | 2.45 ± 0.40 |
| P | 126.31 * ± 46.02 | P | 4.58 * ± 0.61 |
| 2-Ketoglucose Dimethylacetal | Xylose | ||
| D | 0.16 ± 0.05 | D | 0.79 ± 0.10 |
| P | 0.35 * ± 0.05 | P | 1.20 * ± 0.05 |
| Glycerol | Glucosamine | ||
| D | 195.86 ± 34.98 | D | 0.70 ± 0.18 |
| P | 308.98 * ± 37.76 | P | 1.06 * ± 0.14 |
| 2-Hydroxyvaleric Acid | Pentonolactone | ||
| D | 2.79 ± 0.46 | D | 2.44 ± 0.84 |
| P | 3.55 * ± 0.49 | P | 3.66 * ± 0.27 |
| 4-Hydroxybutyric Acid | Threitol | ||
| D | 13.86 ± 3.64 | D | 0.42 ± 0.10 |
| P | 23.50 * ± 3.05 | P | 0.70 * ± 0.15 |
| Myo-Inositol | Guanidinosuccinate | ||
| D | 15.64 ± 3.93 | D | 0.36 ± 0.12 |
| P | 32.54 * ± 6.86 | P | 0.73 * ± 0.20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ren, H.; Wang, H.; Wang, Q.; Qi, X.; Zhang, S.; Yu, Z.; Ijaz, M.; Zhang, M.; Ahmed, T.; El-Sharnouby, M.; et al. Effect of Fungicides on Bayberry Decline Disease by Modulating Rhizosphere Soil Properties, Microflora, and Metabolites. Agronomy 2022, 12, 677. https://doi.org/10.3390/agronomy12030677
Ren H, Wang H, Wang Q, Qi X, Zhang S, Yu Z, Ijaz M, Zhang M, Ahmed T, El-Sharnouby M, et al. Effect of Fungicides on Bayberry Decline Disease by Modulating Rhizosphere Soil Properties, Microflora, and Metabolites. Agronomy. 2022; 12(3):677. https://doi.org/10.3390/agronomy12030677
Chicago/Turabian StyleRen, Haiying, Hongyan Wang, Qi Wang, Xingjiang Qi, Shuwen Zhang, Zheping Yu, Munazza Ijaz, Muchen Zhang, Temoor Ahmed, Mohamed El-Sharnouby, and et al. 2022. "Effect of Fungicides on Bayberry Decline Disease by Modulating Rhizosphere Soil Properties, Microflora, and Metabolites" Agronomy 12, no. 3: 677. https://doi.org/10.3390/agronomy12030677