
The Effect of Nitrogen Fertilizer and Water Supply Levels on the Growth, Antioxidant Compounds, and Organic Acids of Baby Lettuce



Abstract
:1. Introduction
2. Material and Methods
2.1. Growing Conditions
2.2. Experimental Design
2.3. Measurement of Chlorophyll Concentration, Plant Growth and Dry Matter Content
2.4. Quantification of Organic Acids
2.5. Quantification of Total Antioxidant Compounds
2.6. Data Analysis
3. Results
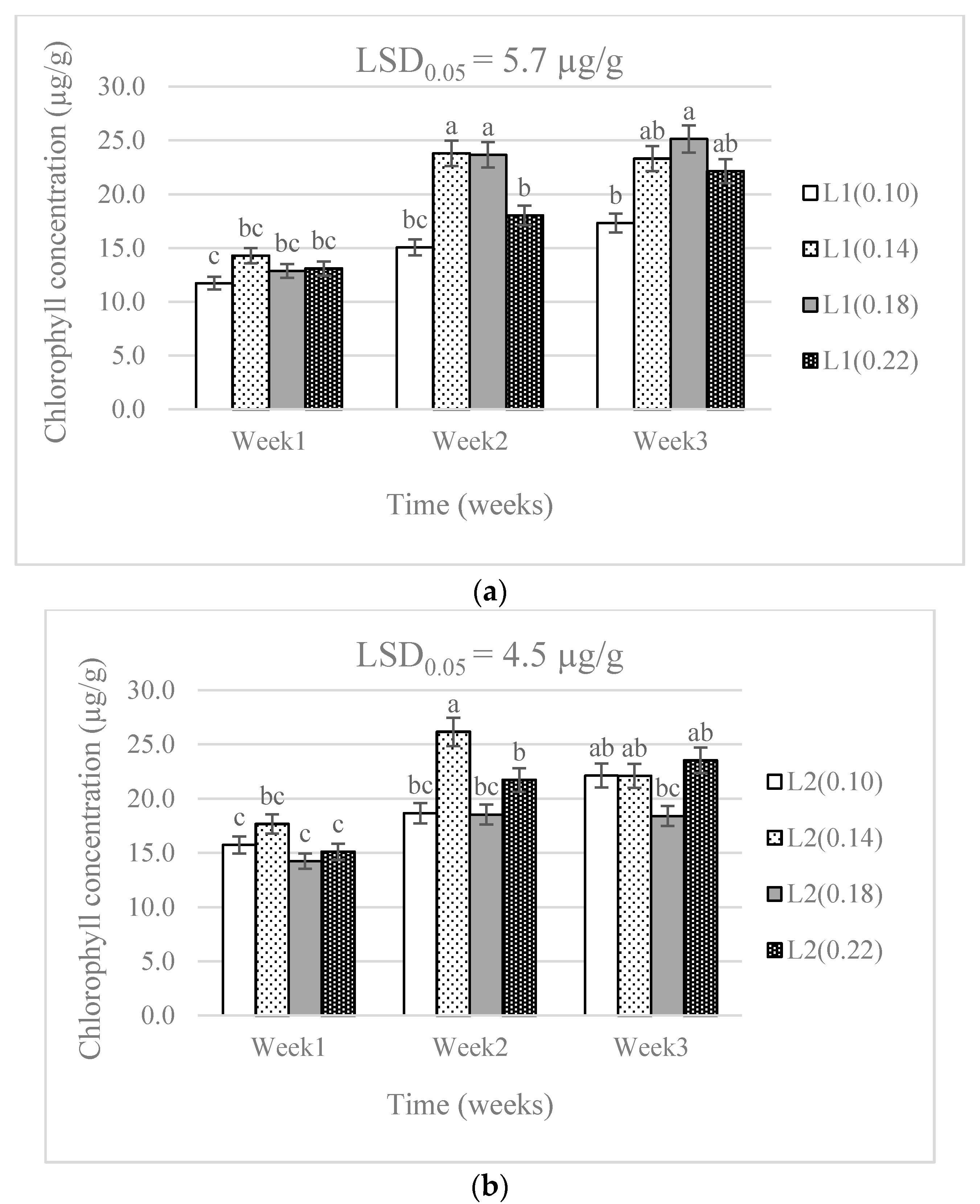
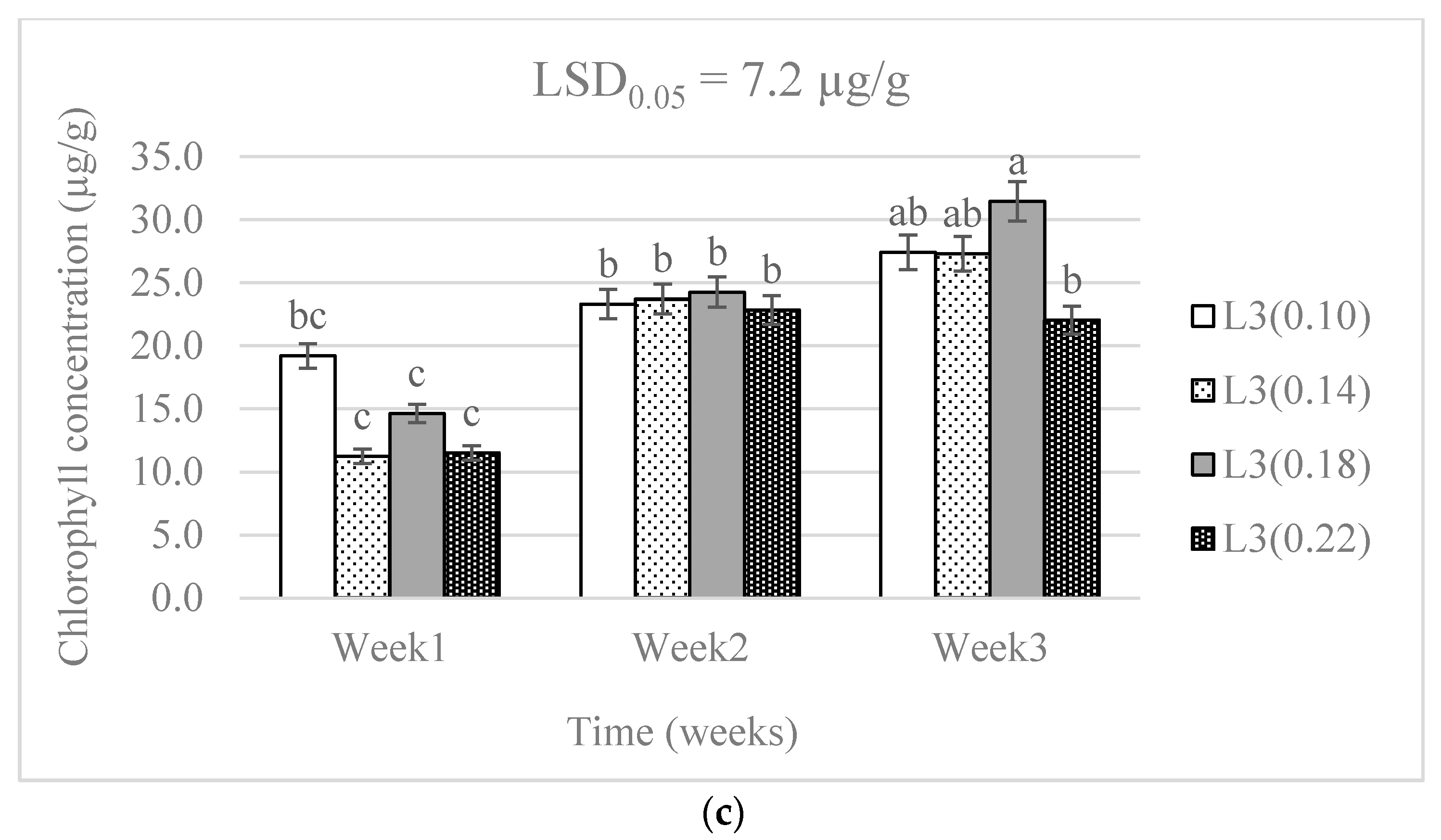
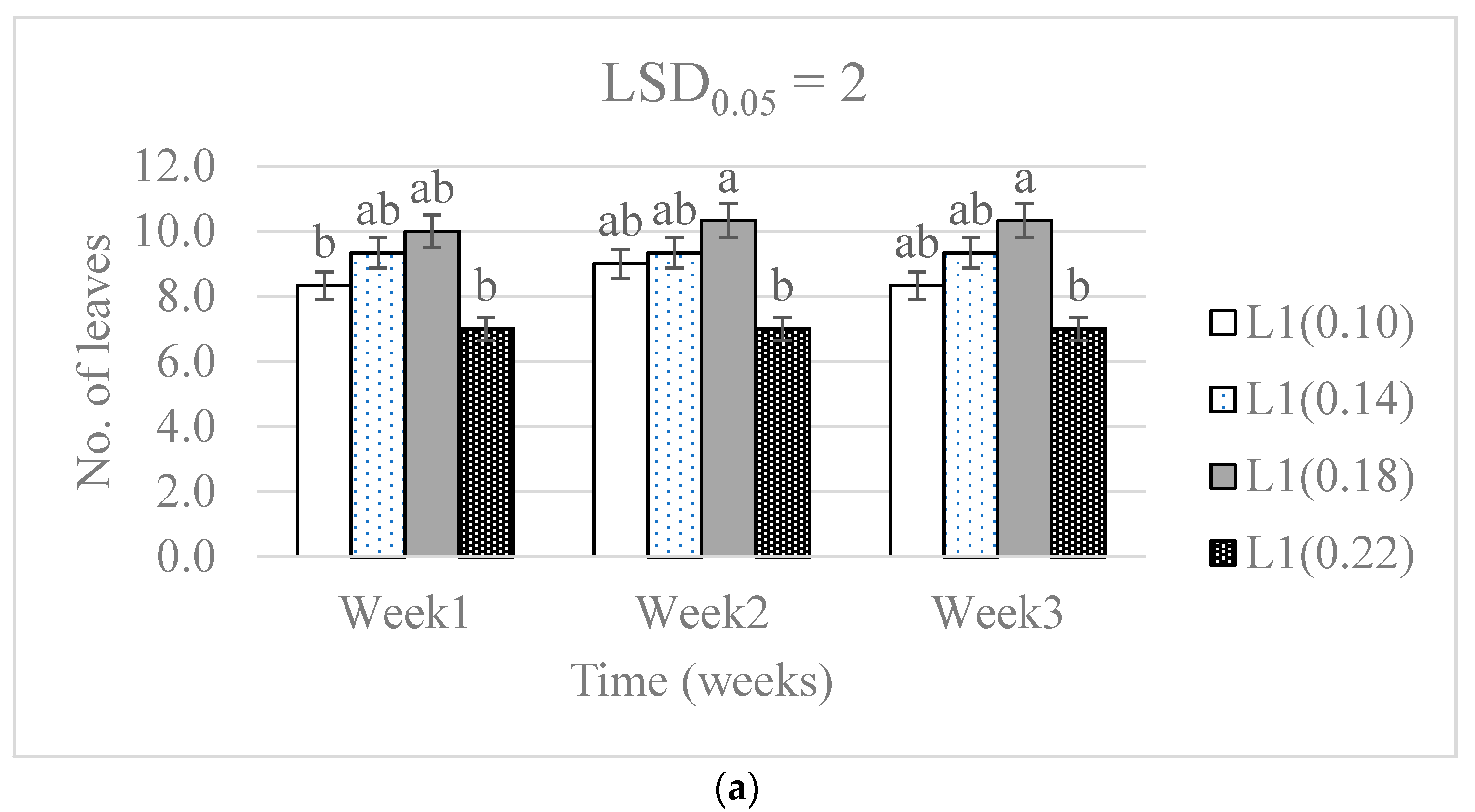
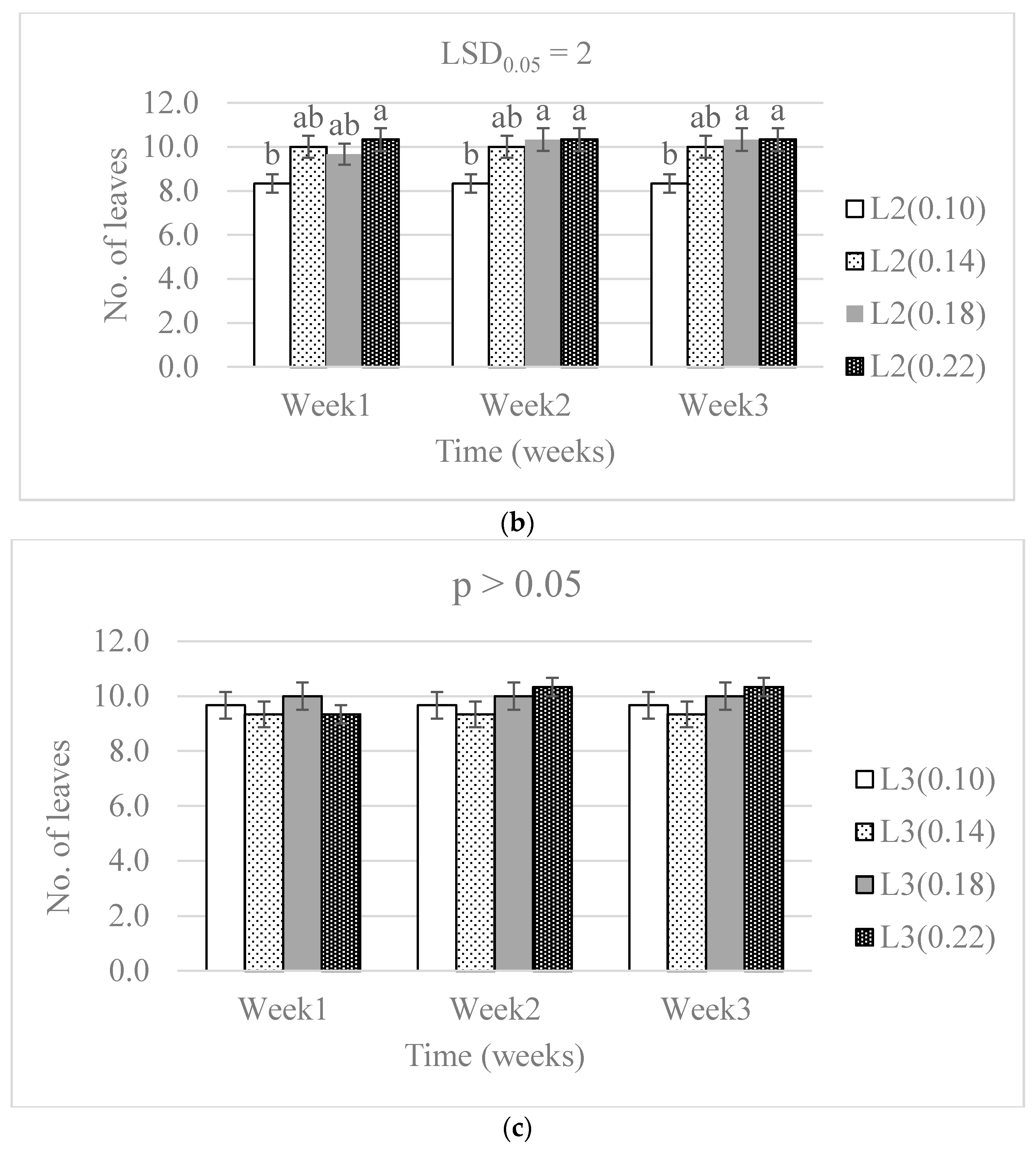
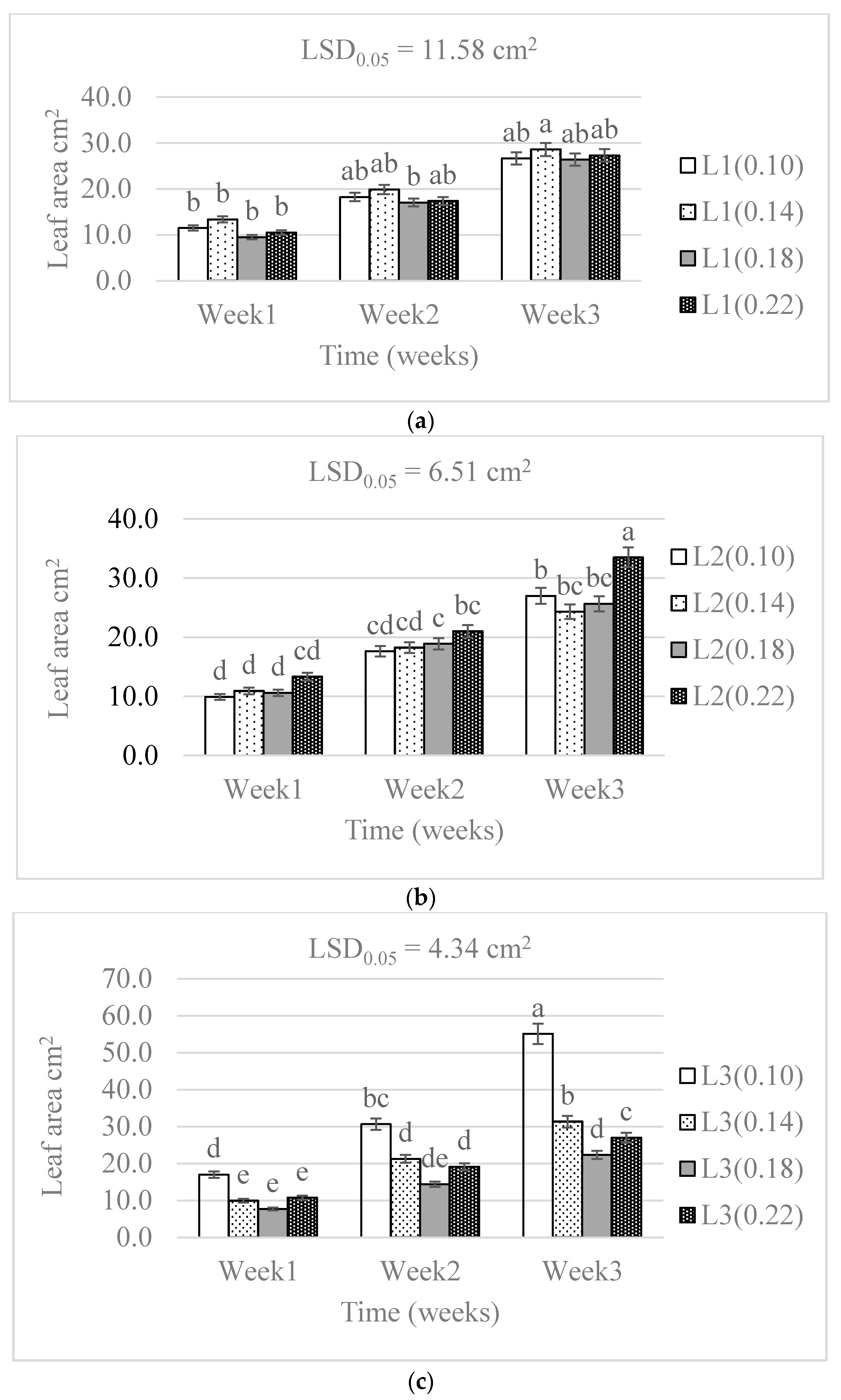
3.1. Chlorophyll Concentration and Plant Growth
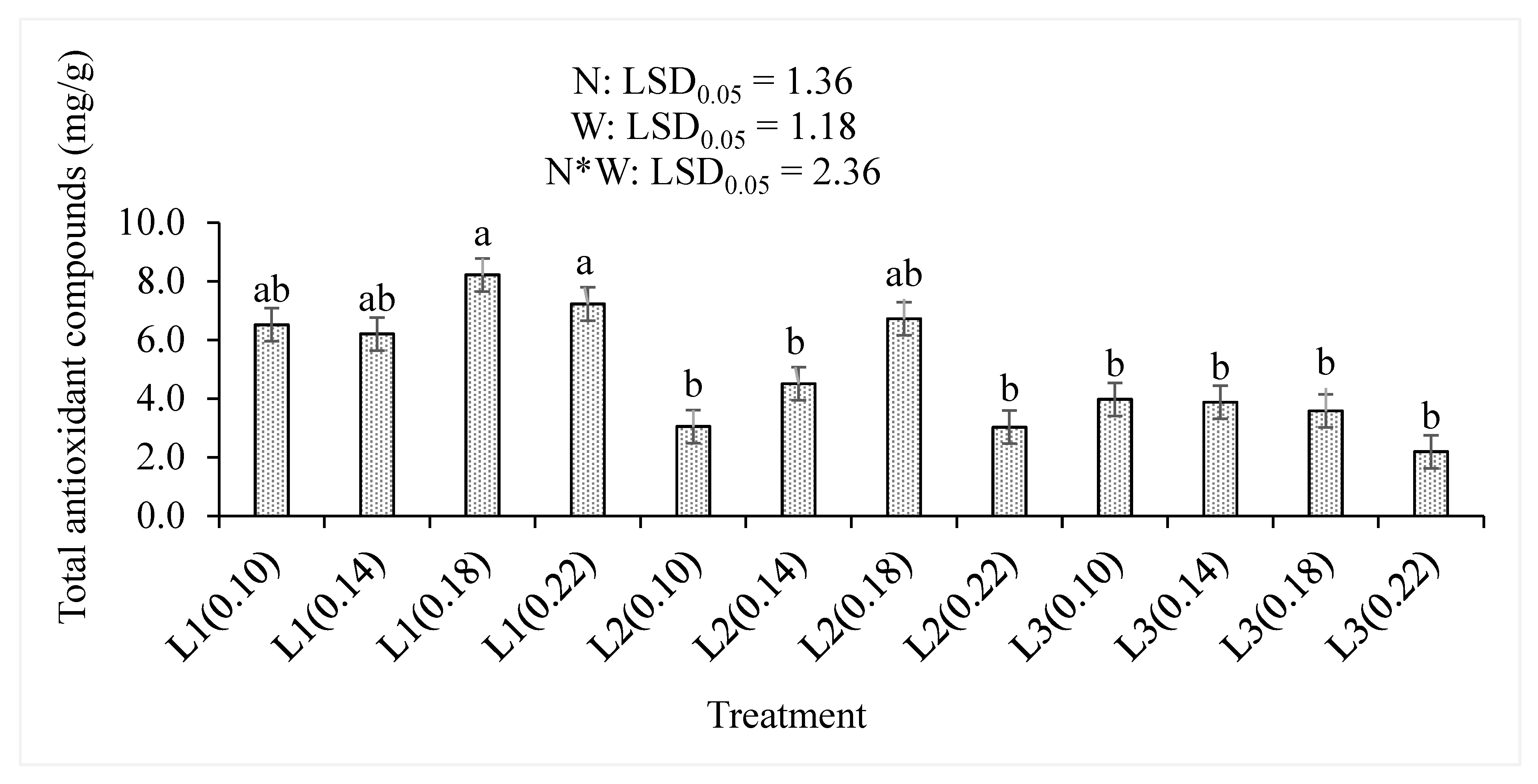
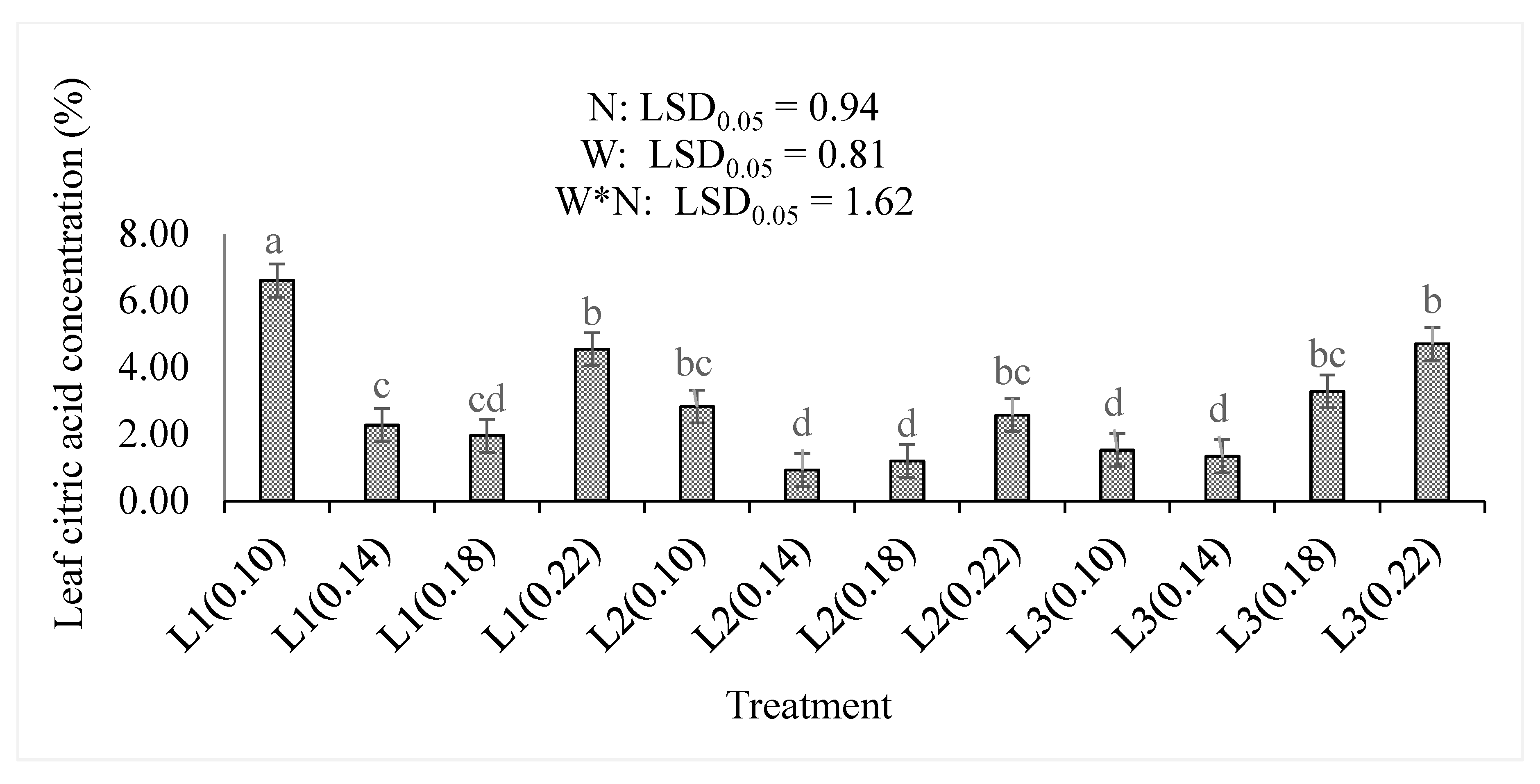
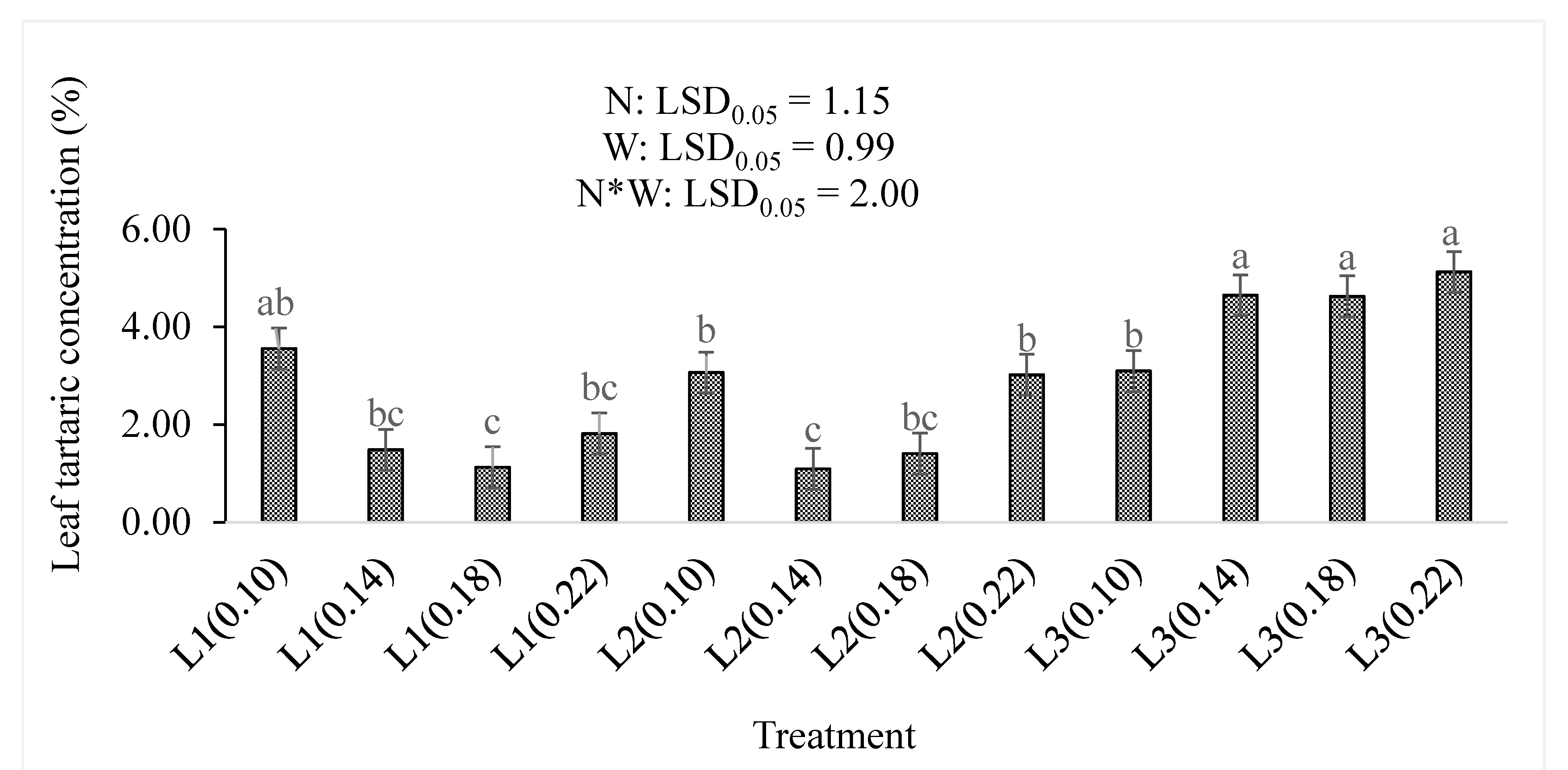
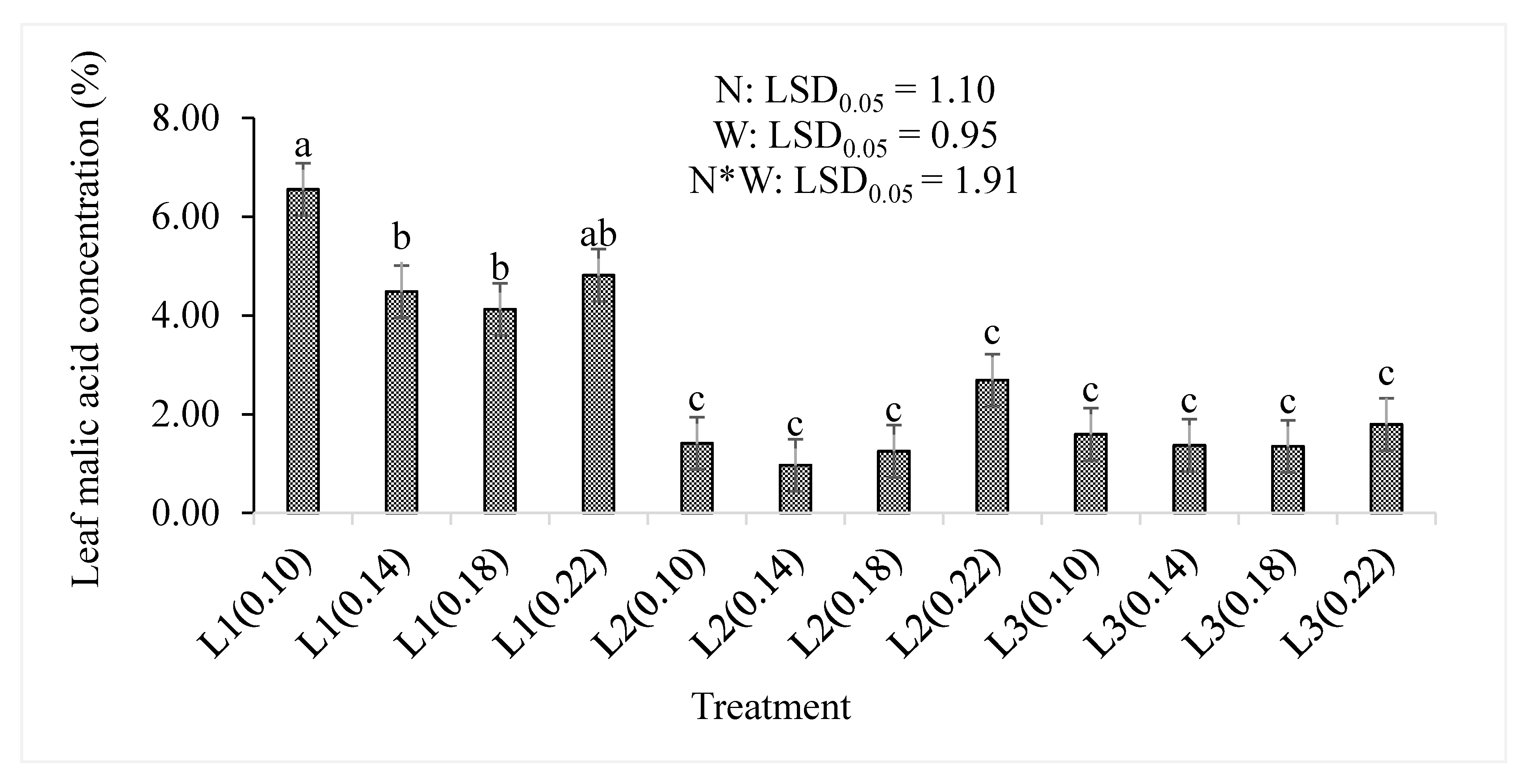
3.2. Total Antioxidant Compounds, Organic Acids, and Dry Matter Content
3.3. Correlation of Growth Parameters to Chlorophyll, Antioxidants, Organic Acids, and Dry Matter Percentage
4. Discussion
4.1. Chlorophyll Concentration and Plant Growth
4.2. Total Antioxidant Compounds, Organic Acids, and Dry Matter Percentages
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kim, M.J.; Moon, Y.; Tou, J.C.; Mou, B.; Waterland, N.L. Nutritional value, bioactive compounds and health benefits of lettuce (Lactuca sativa L.). J. Food Compos. Anal. 2016, 49, 19–34. [Google Scholar] [CrossRef]
- Dimitrov, I.; Stancheva, I.; Mitova, J.; Atanasova, E. Comparative study of some quality parameters of lettuce in dependence on way of cultivation. Bulg. J. Agric. Sci. 2006, 12, 421. [Google Scholar]
- Abdullah, M.; Jamil, R.T.; Attia, F.N. Vitamin C (Ascorbic Acid). StatPearls. 2020. Available online: https://www.ncbi.nlm.nih.gov/books/NBK499877/ (accessed on 13 February 2021).
- Castañer, M.; Gil, M.I.; Artés, F. Organic acids as browning inhibitors on harvested “Baby” lettuce and endive. Z. Lebensm. und-Forsch. A 1997, 205, 375–379. [Google Scholar] [CrossRef]
- Wang, J.; Tao, D.; Liu, Y.; Han, S.; Zheng, F.; Wu, Z. Comparison of generally recognized as safe organic acids for disinfecting fresh-cut lettuce. BioRxiv 2018. [Google Scholar] [CrossRef]
- Wang, J.; Tao, D.; Wang, S.; Li, C.; Li, Y.; Zheng, F.; Wu, Z. Disinfection of lettuce using organic acids: An ecological analysis using 16S rRNA sequencing. RSC Adv. 2019, 9, 17514–17520. [Google Scholar] [CrossRef] [Green Version]
- Akbas, M.Y.; Ölmez, H. Inactivation of Escherichia coli and Listeria monocytogenes on iceberg lettuce by dip wash treatments with organic acids. Lett. Appl. Microbiol. 2007, 44, 619–624. [Google Scholar] [CrossRef] [PubMed]
- Shams, M.; Yildirim, E.; Ekinci, M.; Turan, M.; Dursun, A.; Parlakova, F.; Kul, R. Exogenously applied glycine betaine regulates some chemical characteristics and antioxidative defence systems in lettuce under salt stress. Hortic. Environ. Biotechnol. 2016, 57, 225–231. [Google Scholar] [CrossRef]
- Doores, S.; Davidson, P.M.; Sofos, J.N.; Branen, A.L. Organic acids. Food Sci. Technol. 2005, 145, 91. [Google Scholar]
- Booth, I.R. Regulation of cytoplasmic pH in bacteria. Microbiol. Rev. 1985, 49, 359. [Google Scholar] [CrossRef]
- Huang, Y.; Chen, H. Effect of organic acids, hydrogen peroxide and mild heat on inactivation of Escherichia coli O157:H7 on baby spinach. Food Control. 2011, 22, 1178–1183. [Google Scholar] [CrossRef]
- López, A.; Javier, G.A.; Fenoll, J.; Hellín, P.; Flores, P. Chemical composition and antioxidant capacity of lettuce: Comparative study of regular-sized (Romaine) and baby-sized (Little Gem and Mini Romaine) types. J. Food Compos. Anal. 2014, 33, 39–48. [Google Scholar] [CrossRef]
- Limantara, L.; Dettling, M.; Indrawati, R.; Indriatmoko, I.; Brotosudarmo, T. Analysis on the Chlorophyll Content of Commercial Green Leafy Vegetables. Procedia Chem. 2015, 14, 225–231. [Google Scholar] [CrossRef] [Green Version]
- Cano, A.; Arnao, M.B. Hydrophilic and Lipophilic Antioxidant Activity in Different Leaves of Three Lettuce Varieties. Int. J. Food Prop. 2005, 8, 521–528. [Google Scholar] [CrossRef]
- Adu, M.O.; Yawson, D.O.; Abano, E.E.; Asare, P.A.; Armah, F.A.; Opoku, E.K. Does water-saving irrigation improve the quality of fruits and vegetables? Evidence from meta-analysis. Irrig. Sci. 2019, 37, 669–690. [Google Scholar] [CrossRef]
- Raun, W.R.; Dhillon, J.; Aula, L.; Eickhoff, E.; Weymeyer, G.; Figueirdeo, B.; Lynch, T.; Omara, P.; Nambi, E.; Oyebiyi, F.; et al. Unpredictable Nature of Environment on Nitrogen Supply and Demand. Agron. J. 2019, 111, 2786–2791. [Google Scholar] [CrossRef] [Green Version]
- Dhakal, C.; Lange, K.; Parajulee, M.N.; Segarra, E. Dynamic Optimization of Nitrogen in Plateau Cotton Yield Functions with Nitrogen Carryover Considerations. J. Agric. Appl. Econ. 2019, 51, 385–401. [Google Scholar] [CrossRef] [Green Version]
- Ncama, K.; Magwaza, L.S.; Tesfay, S.Z.; Mditshwa, A.; Mbili, N.C. In-field application of portable NIR to assess’ Valencia’ orange fruit maturity. In Proceedings of the XXX International Horticultural Congress IHC2018: International Symposium on Strategies and Technologies to Maintain Quality, Istanbul, Turkey, 12–16 August 2018; Volume 1275, pp. 61–68. [Google Scholar]
- Aliyu, A.B.; Ibrahim, M.A.; Musa, A.M.; Musa, A.O.; Kiplimo, J.J.; Oyewale, A.O. Free radical scavenging and total antioxidant capacity of root extracts of Anchomanes difformis Engl. (Araceae). Acta Pol. Pharm. 2013, 70, 115–121. [Google Scholar]
- DAFF. Production Guidelines for Lettuce. Department of Agriculture Fisheries and Forestry, Republic of South Africa. 2020. Available online: https://www.nda.agric.za/docs/Brochures/ProdGuideLettuce.pdf (accessed on 6 July 2021).
- Montemurro, F. Are organic n fertilizing strategies able to improve lettuce yield, use of nitrogen and n status? J. Plant Nutr. 2010, 33, 1980–1997. [Google Scholar] [CrossRef]
- Colom, M.; Vazzana, C. Drought stress effects on three cultivars of Eragrostis curvula: Photosynthesis and water relations. Plant Growth Regul. 2001, 34, 195–202. [Google Scholar] [CrossRef]
- Mu, X.; Chen, Y. The physiological response of photosynthesis to nitrogen deficiency. Plant Physiol. Biochem. 2021, 158, 76–82. [Google Scholar] [CrossRef]
- Karam, F.; Mounzer, O.; Sarkis, F.; Lahoud, R. Yield and nitrogen recovery of lettuce under different irrigation regimes. J. Appl. Hortic. 2002, 4, 70–76. [Google Scholar] [CrossRef]
- Qadir, O.; Siervo, M.; Seal, C.J.; Brandt, K. Manipulation of Contents of Nitrate, Phenolic Acids, Chlorophylls, and Carotenoids in Lettuce (Lactuca sativa L.) via Contrasting Responses to Nitrogen Fertilizer When Grown in a Controlled Environment. J. Agric. Food Chem. 2017, 65, 10003–10010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Nakhel, C.; Pannico, A.; Graziani, G.; Kyriacou, M.C.; Giordano, M.; Ritieni, A.; De Pascale, S.; Rouphael, Y. Variation in macronutrient content, phytochemical constitution and in vitro antioxidant capacity of green and red butter-head lettuce dictated by different developmental stages of harvest maturity. Antioxidants 2020, 9, 300. [Google Scholar] [CrossRef] [Green Version]
- Ncama, K.; Aremu, O.A.; Sithole, N.J. Plant Adaptation to Environmental Stress: Drought, Chilling, Heat, and Salinity. In Environment and Climate-Smart Food Production; Springer: Singapore, 2021; pp. 151–179. [Google Scholar]
- Prado, K.; Maurel, C. Regulation of leaf hydraulics: From molecular to whole plant levels. Front. Plant Sci. 2013, 4, 255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moosavi, S.G. The effect of water deficit stress and nitrogen fertilizer levels on morphology traits, yield and leaf area index in maize. Pak. J. Bot. 2012, 44, 1351–1355. [Google Scholar]
- Wright, I.J.; Reich, P.B.; Westoby, M. Least-cost input mixtures of water and nitrogen for photosynthesis. Am. Nat. 2003, 161, 98–111. [Google Scholar] [CrossRef]
- Rocha, D.C.; da Silva, B.F.I.; Moreira dos Santos, J.M.; Tavares, D.S.; Pauletti, V.; Gomes, M.P. Do nitrogen sources and molybdenum affect the nutritional quality and nitrate concentrations of hydroponic baby leaf lettuce? J. Food Sci. 2020, 85, 1605–1612. [Google Scholar] [CrossRef]
- Zhang, F.; Wan, X.; Zheng, Y.; Sun, L.; Chen, Q.; Zhu, X.; Guo, Y.; Liu, M. Effects of nitrogen on the activity of antioxidant enzymes and gene expression in leaves of Populus plants subjected to cadmium stress. J. Plant Interact. 2014, 9, 599–609. [Google Scholar] [CrossRef]
- Camalle, M.; Standing, D.; Jitan, M.; Muhaisen, R.; Bader, N.; Bsoul, M.; Ventura, Y.; Soltabayeva, A.; Sagi, M. Effect of Salinity and Nitrogen Sources on the Leaf Quality, Biomass, and Metabolic Responses of Two Ecotypes of Portulaca oleracea. Agronomy 2020, 10, 656. [Google Scholar] [CrossRef]
- Wang, Y.; Qiao, Y.; Zhang, M.; Ma, Z.; Xue, Y.; Mi, Q.; Wang, A.; Feng, J. Potential value of small-molecule organic acids for the control of postharvest gray mold caused by Botrytis cinerea. Pestic. Biochem. Physiol. 2021, 177, 104884. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Factor of Variability | Factor Effect | p-Value |
|---|---|---|---|
| Chlorophyll concentration | N | − | 0.343 |
| W | + | 0.036 | |
| N*W | − | 0.049 | |
| Leaf number | N | + | 0.009 |
| W | − | 0.006 | |
| N*W | + | <0.001 | |
| Leaf area | N | − | 0.007 |
| W | + | 0.154 | |
| N*W | + | <0.001 |
| Parameter | Factor of Variability | Factor Effect | p-Value |
|---|---|---|---|
| Total antioxidants (mg/g) | N | + | <0.001 |
| W | − | <0.001 | |
| N*W | + | 0.001 | |
| Citric acid % | N | + | <0.001 |
| W | − | <0.001 | |
| N*W | − | <0.001 | |
| Tartaric acid % | N | − | 0.196 |
| W | + | <0.001 | |
| N*W | + | 0.057 | |
| Malic acid % | N | − | 0.166 |
| W | − | <0.001 | |
| N*W | − | 0.353 | |
| Dry matter % | N | − | <0.001 |
| W | − | <0.001 | |
| N*W | − | 0.004 |
| Chlorophyll (µg/g FM) | No. of Leaves | Leaf Area (cm2) | TAO (mg/g FM) | Citric Acid % | Tartaric Acid % | Malic Acid % | Dry Matter % | |
|---|---|---|---|---|---|---|---|---|
| Chlorophyll (µg/g FM) | 1 | |||||||
| No. of leaves | 0.15 | 1 | ||||||
| Leaf area (cm2) | 0.58 | 0.11 | 1 | |||||
| TAO (mg/g FM) | −0.02 | 0.20 | −0.18 | 1 | ||||
| Citric acid (%) | −0.19 | −0.31 | −0.14 | 0.11 | 1 | |||
| Tartaric acid (%) | −0.03 | 0.03 | 0.01 | −0.13 | 0.42 | 1 | ||
| Malic acid (%) | −0.11 | −0.23 | −0.20 | 0.22 | 0.74 | 0.10 | 1 | |
| Dry matter (%) | 0.10 | −0.26 | 0.03 | 0.09 | 0.37 | −0.46 | 0.56 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ncama, K.; Sithole, N.J. The Effect of Nitrogen Fertilizer and Water Supply Levels on the Growth, Antioxidant Compounds, and Organic Acids of Baby Lettuce. Agronomy 2022, 12, 614. https://doi.org/10.3390/agronomy12030614
Ncama K, Sithole NJ. The Effect of Nitrogen Fertilizer and Water Supply Levels on the Growth, Antioxidant Compounds, and Organic Acids of Baby Lettuce. Agronomy. 2022; 12(3):614. https://doi.org/10.3390/agronomy12030614
Chicago/Turabian StyleNcama, Khayelihle, and Nkanyiso Justice Sithole. 2022. "The Effect of Nitrogen Fertilizer and Water Supply Levels on the Growth, Antioxidant Compounds, and Organic Acids of Baby Lettuce" Agronomy 12, no. 3: 614. https://doi.org/10.3390/agronomy12030614