
Towards Heat Tolerant Runner Bean (Phaseolus coccineus L.) by Utilizing Plant Genetic Resources

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
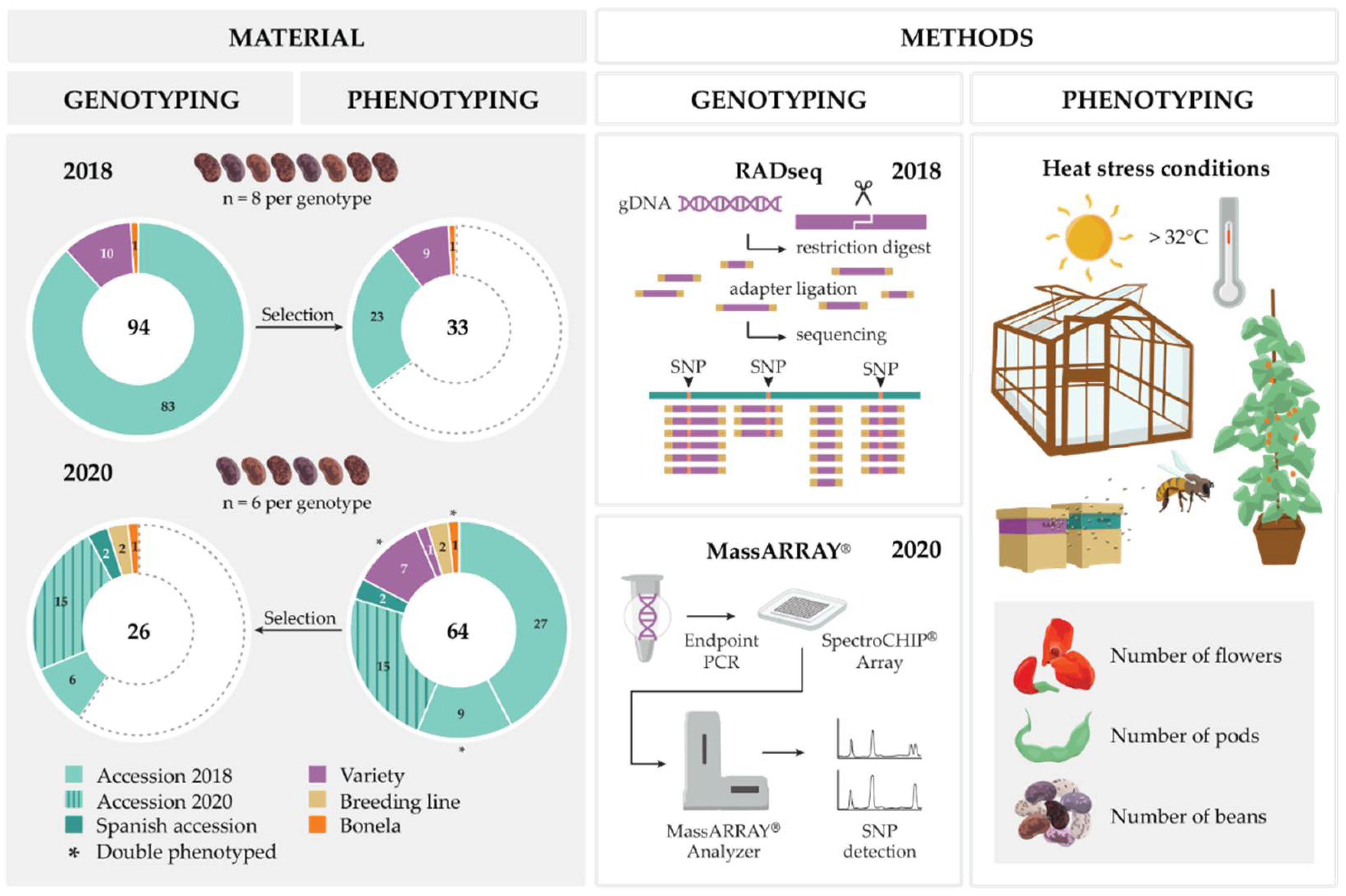
2.1. Plant Material
2.2. Trial Design
2.3. Glasshouse Trials and Phenotyping
2.4. Leaf Sampling and DNA Extraction
2.5. RAD Sequencing and Data Analysis
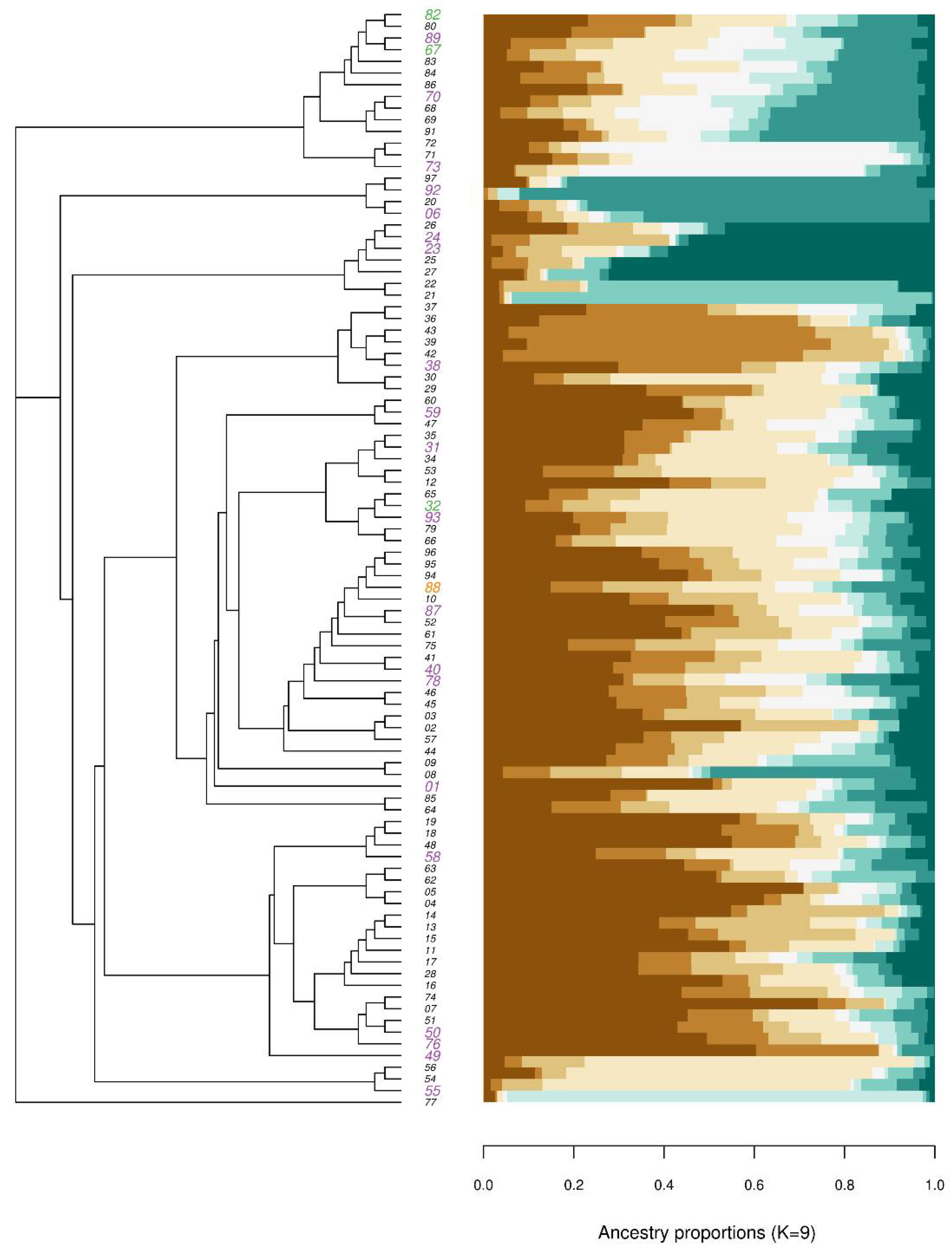
2.6. Phylogeny and Ancestry Matrix
2.7. Genome-Wide Association Studies (GWAS)
2.8. MassARRAY® Design and Genotyping
2.9. SNP Selection and Calculation of Heat Tolerance
3. Results
3.1. Plant Development and Heat Stress Reaction
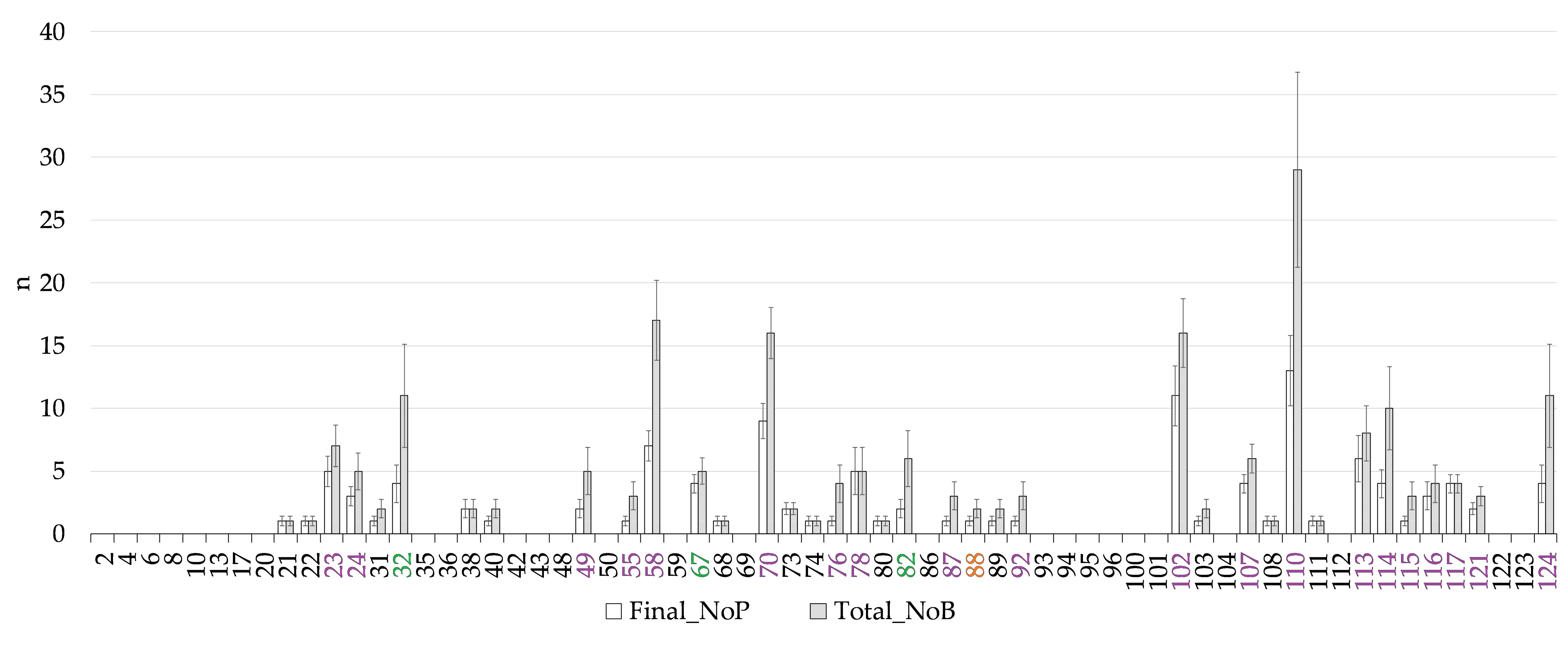
3.2. Phenotypic Diversity
3.3. Phylogenetic Relationship
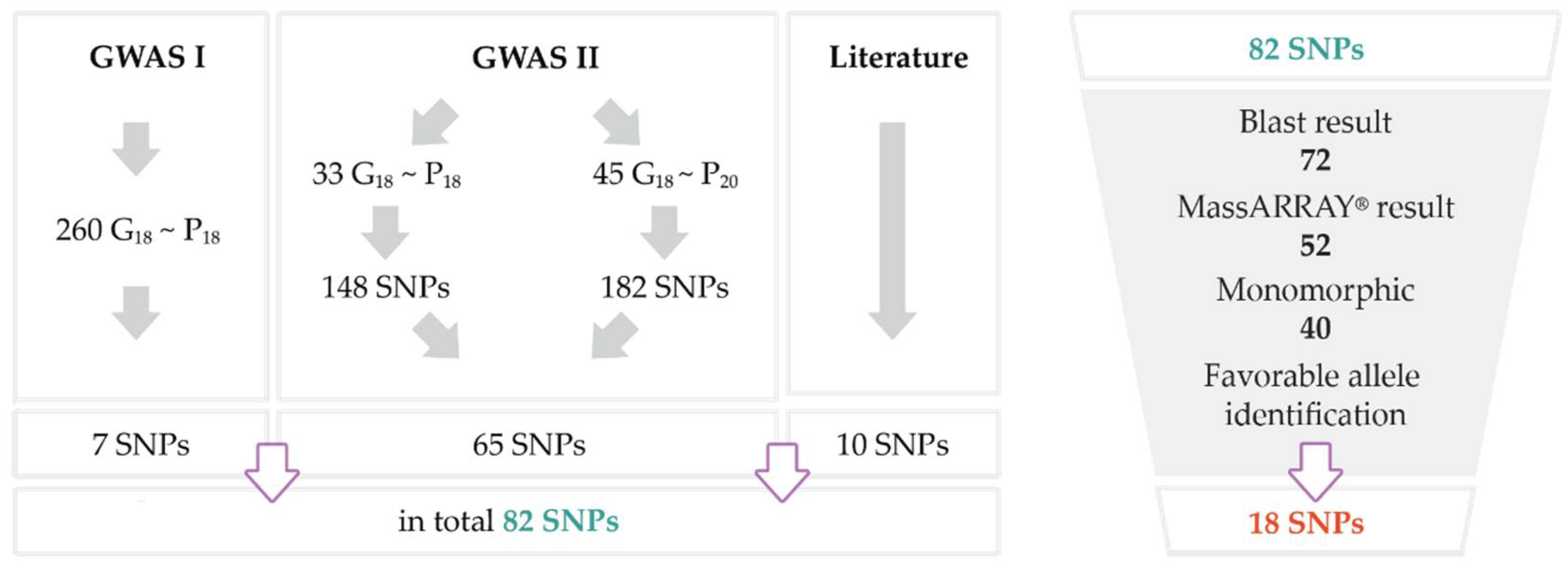
3.4. Results of Genome-Wide Association Studies (GWAS)
3.5. Selection of SNPs
3.6. Characterisation of Selected SNPs
3.7. Estimation of Heat Tolerance
4. Discussion
4.1. Genetic Diversity Analysis for Gene Bank Management and Breeding
4.2. Challenges of Breeding Heat Stress-Tolerant Runner Bean
4.3. Molecular Markers for Runner Bean Breeding
4.4. Towards Heat Tolerant Runner Bean Varieties
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bitocchi, E.; Rau, D.; Bellucci, E.; Rodriguez, M.; Murgia, M.L.; Gioia, T.; Santo, D.; Nanni, L.; Attene, G.; Papa, R. Beans (Phaseolus ssp.) as a model for understanding crop evolution. Front. Plant Sci. 2017, 8, 722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Commission. EU Plant Variety Database. Available online: https://ec.europa.eu/food/plant/plant_propagation_material/plant_variety_catalogues_databases/search/public/index.cfm?event=SearchVariety&ctl_type=H&species_id=31&variety_name=&listed_in=0&show_current=on&show_deleted (accessed on 30 December 2021).
- Rodiño, A.P.; Lema, M.; Pérez-Barbeito, M.; Santalla, M.; De Ron, A.M. Assessment of runner bean (Phaseolus coccineus L.) germplasm for tolerance to low temperature during early seedling growth. Euphytica 2006, 155, 63–70. [Google Scholar] [CrossRef] [Green Version]
- Santalla, M.; Monteagudo, A.B.; González, A.M.; De Ron, A.M. Agronomical and quality traits of runner bean germplasm and implications for breeding. Euphytica 2004, 135, 205–215. [Google Scholar] [CrossRef]
- Sicard, D.; Nanni, L.; Porfiri, O.; Bulfon, D.; Papa, R. Genetic diversity of Phaseolus vulgaris L. and P. coccineus L. landraces in central Italy. Plant Breed. 2005, 124, 464–472. [Google Scholar] [CrossRef]
- De Ron, A.M.; Bebeli, P.J.; Negri, V.; Vaz Patto, M.C.; Revilla, P. Warm season grain legume landraces from the south of Europe for germplasm conservation and genetic improvement. Front. Plant Sci. 2018, 9, 1524. [Google Scholar] [CrossRef] [PubMed]
- Palmero, D.; Iglesias, C.; Cara, M.d.; Tello, J.C.; Camacho, F.G. Diversity and health traits of local landraces of runner bean (Phaseolus coccineus L.) from Spain. J. Food Agric. Environ. 2011, 9, 290–295. [Google Scholar]
- Genesys. Global Portal on Plant Genetic Resources. Available online: https://www.genesys-pgr.org/a/overview/v2r8e87E7XJ (accessed on 11 December 2021).
- AGES Gene Bank. National Inventory. Available online: https://www.genbank.at/en/national-inventory/search-result.html (accessed on 12 December 2021).
- European Commission. Commission Implementing Regulation (EU) 2016/1408 of 12 August 2016 Entering a Name in the Register of Protected Designations of Origin and Protected Geographical Indications (Steirische Käferbohne (PDO)). Off. J. Union 2016. [Google Scholar]
- Statistik Austria. Gemüseproduktion ab 1995. Available online: http://www.statistik.at/web_de/statistiken/wirtschaft/land_und_forstwirtschaft/agrarstruktur_flaechen_ertraege/gemuese/index.html (accessed on 6 January 2022).
- De Ron, A.M.; Rodiño, A.P.; Santalla, M.; González, A.M.; Lema, M.J.; Martín, I.; Kigel, J. Seedling emergence and phenotypic response of common bean germplasm to different temperatures under controlled conditions and in open Field. Front. Plant Sci. 2016, 7, 1087. [Google Scholar] [CrossRef] [Green Version]
- Porch, T.G.; Jahn, M. Effects of high-temperature stress on microsporogenesis in heat-sensitive and heat-tolerant genotypes of Phaseolus vulgaris. Plant Cell Environ. 2001, 24, 723–731. [Google Scholar] [CrossRef]
- Rainey, K.M.; Griffiths, P.D. Differential response of common bean genotypes to high temperatures. J. Am. Soc. Hortic. Sci. 2005, 130, 18–23. [Google Scholar] [CrossRef] [Green Version]
- Vargas, Y.; Mayor-Duran, V.M.; Buendia, H.F.; Ruiz-Guzman, H.; Raatz, B. Physiological and genetic characterization of heat stress effects in a common bean RIL population. PLoS ONE 2021, 16, e0249859. [Google Scholar] [CrossRef] [PubMed]
- Galluzzi, G.; Seyoum, A.; Halewood, M.; López Noriega, I.; Welch, E.W. The role of genetic resources in breeding for climate change: The case of public breeding programmes in eighteen developing countries. Plants 2020, 9, 1129. [Google Scholar] [CrossRef] [PubMed]
- Hodgkin, T.; Bordoni, P. Climate change and the conservation of plant genetic resources. J. Crop Improv. 2012, 26, 329–345. [Google Scholar] [CrossRef]
- Assefa, T.; Assibi Mahama, A.; Brown, A.V.; Cannon, E.K.S.; Rubyogo, J.C.; Rao, I.M.; Blair, M.W.; Cannon, S.B. A review of breeding objectives, genomic resources, and marker-assisted methods in common bean (Phaseolus vulgaris L.). Mol. Breed. 2019, 39, 20. [Google Scholar] [CrossRef] [Green Version]
- D’Agostino, N.; Tripodi, P. NGS-based genotyping, high-throughput phenotyping and genome-wide association studies laid the foundations for next-generation breeding in horticultural crops. Diversity 2017, 9, 38. [Google Scholar] [CrossRef] [Green Version]
- Halewood, M.; Chiurugwi, T.; Sackville Hamilton, R.; Kurtz, B.; Marden, E.; Welch, E.; Michiels, F.; Mozafari, J.; Sabran, M.; Patron, N.; et al. Plant genetic resources for food and agriculture: Opportunities and challenges emerging from the science and information technology revolution. New Phytol. 2018, 217, 1407–1419. [Google Scholar] [CrossRef]
- Delfini, J.; Moda-Cirino, V.; Dos Santos Neto, J.; Zeffa, D.M.; Nogueira, A.F.; Ribeiro, L.A.B.; Ruas, P.M.; Gepts, P.; Goncalves, L.S.A. Genome-wide association study identifies genomic regions for important morpho-agronomic traits in Mesoamerican common bean. Front. Plant Sci. 2021, 12, 748829. [Google Scholar] [CrossRef]
- Nadeem, M.A.; Habyarimana, E.; Çiftçi, V.; Nawaz, M.A.; Karaköy, T.; Comertpay, G.; Shahid, M.Q.; Hatipoğlu, R.; Yeken, M.Z.; Ali, F.; et al. Characterization of genetic diversity in Turkish common bean gene pool using phenotypic and whole-genome DArTseq-generated silicoDArT marker information. PLoS ONE 2018, 13, e0205363. [Google Scholar] [CrossRef]
- Mascher, M.; Schreiber, M.; Scholz, U.; Graner, A.; Reif, J.C.; Stein, N. Genebank genomics bridges the gap between the conservation of crop diversity and plant breeding. Nat. Genet. 2019, 51, 1076–1081. [Google Scholar] [CrossRef]
- Arriagada, O.; Schwember, A.R.; Greve, M.J.; Urban, M.O.; Cabeza, R.A.; Carrasco, B. Morphological and molecular characterization of selected Chilean runner bean (Phaseolus coccineus L.) genotypes shows moderate agronomic and genetic variability. Plants 2021, 10, 1688. [Google Scholar] [CrossRef]
- Rodriguez, M.; Rau, D.; Angioi, S.A.; Bellucci, E.; Bitocchi, E.; Nanni, L.; Knupffer, H.; Negri, V.; Papa, R.; Attene, G. European Phaseolus coccineus L. landraces: Population structure and adaptation, as revealed by cpSSRs and phenotypic analyses. PLoS ONE 2013, 8, e57337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spataro, G.; Tiranti, B.; Arcaleni, P.; Bellucci, E.; Attene, G.; Papa, R.; Spagnoletti Zeuli, P.; Negri, V. Genetic diversity and structure of a worldwide collection of Phaseolus coccineus L. Theor. Appl. Genet. 2011, 122, 1281–1291. [Google Scholar] [CrossRef]
- López-Hernández, F.; Cortés, A.J. Last-generation genome-environment associations reveal the genetic basis of heat tolerance in common bean (Phaseolus vulgaris L.). Front. Genet. 2019, 10, 954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oladzad, A.; Porch, T.; Rosas, J.C.; Moghaddam, S.M.; Beaver, J.; Beebe, S.E.; Burridge, J.; Jochua, C.N.; Miguel, M.A.; Miklas, P.N.; et al. Single and multi-trait GWAS identify genetic factors associated with production traits in common bean under abiotic stress environments. G3 Genes Genomes Genet. 2019, 9, 1881–1892. [Google Scholar] [CrossRef]
- Tibbs Cortes, L.; Zhang, Z.; Yu, J. Status and prospects of genome-wide association studies in plants. Plant Genome 2021, 14, e20077. [Google Scholar] [CrossRef]
- Ellis, J.A.; Ong, B. The MassARRAY® system for targeted SNP genotyping. Methods Mol. Biol. 2017, 1492, 77–94. [Google Scholar] [CrossRef]
- Sudan, J.; Singh, R.; Sharma, S.; Salgotra, R.K.; Sharma, V.; Singh, G.; Sharma, I.; Sharma, S.; Gupta, S.K.; Zargar, S.M. ddRAD sequencing-based identification of inter-genepool SNPs and association analysis in Brassica juncea. BMC Plant Biol. 2019, 19, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Smith, H.M.; Smith, B.P.; Morales, N.B.; Moskwa, S.; Clingeleffer, P.R.; Thomas, M.R. SNP markers tightly linked to root knot nematode resistance in grapevine (Vitis cinerea) identified by a genotyping-by-sequencing approach followed by Sequenom MassARRAY validation. PLoS ONE 2018, 13, e0193121. [Google Scholar] [CrossRef] [Green Version]
- BAES (Ed.) Österreichische Sortenliste 2022; Schriftenreihe: Berlin, Germany, 2022; ISSN 1560-635X. [Google Scholar]
- van der Beek, J.G.; Verkerk, R.; Zabel, P.; Lindhout, P. Mapping strategy for resistance genes in tomato based on RFLPs between cultivars: Cf9 (resistance to Cladosporium fulvum) on chromosome 1. Theor. Appl. Genet. 1992, 84, 106–112. [Google Scholar] [CrossRef]
- Baird, N.A.; Etter, P.D.; Atwood, T.S.; Currey, M.C.; Shiver, A.L.; Lewis, Z.A.; Selker, E.U.; Cresko, W.A.; Johnson, E.A. Rapid SNP discovery and genetic mapping using sequenced RAD markers. PLoS ONE 2008, 3, e3376. [Google Scholar] [CrossRef]
- Aronesty, E. Comparison of sequencing utility programs. Open Bioinform. J. 2013, 7, 1–8. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. 2010. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 24 November 2021).
- Ewels, P.; Magnusson, M.; Lundin, S.; Kaller, M. MultiQC: Summarize analysis results for multiple tools and samples in a single report. Bioinformatics 2016, 32, 3047–3048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eaton, D.A.R.; Overcast, I. ipyrad: Interactive assembly and analysis of RADseq datasets. Bioinformatics 2020, 36, 2592–2594. [Google Scholar] [CrossRef] [PubMed]
- Danecek, P.; Auton, A.; Abecasis, G.; Albers, C.A.; Banks, E.; DePristo, M.A.; Handsaker, R.E.; Lunter, G.; Marth, G.T.; Sherry, S.T.; et al. The variant call format and VCFtools. Bioinformatics 2011, 27, 2156–2158. [Google Scholar] [CrossRef]
- Jombart, T. adegenet: A R package for the multivariate analysis of genetic markers. Bioinformatics 2008, 24, 1403–1405. [Google Scholar] [CrossRef] [Green Version]
- Paradis, E.; Schliep, K. ape 5.0: An environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics 2019, 35, 526–528. [Google Scholar] [CrossRef]
- Kamvar, Z.; Tabima, J.; Grünwald, N. Poppr: An R package for genetic analysis of populations with clonal, partially clonal, and/or sexual reproduction. PeerJ 2014, 2, e281. [Google Scholar] [CrossRef] [Green Version]
- Frichot, E.; François, O. LEA: An R package for landscape and ecological association studies. Methods Ecol. Evol. 2015, 6, 925–929. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Wickham, H. ggplot2. WIREs Comput. Stat. 2011, 3, 180–185. [Google Scholar] [CrossRef]
- Ahlmann-Eltze, C.; Patil, I. ggsignif: R package for displaying significance brackets for ‘ggplot2’. PsyArvix 2021. [Google Scholar] [CrossRef]
- Bundesanstalt für Pflanzenbau. Feuerbohne Hara. In Proceedings of the Zuchtbuchkommission des Bundesministeriums für Land- und Forstwirtschaft, Sorten gärtnerischer Kulturpflanzen, Vienna, Austria, 19 December 1985; p. 1. [Google Scholar]
- Bundesanstalt für Pflanzenbau. Stangenfeuerbohne Bonela. In Proceedings of the Zuchtbuchkommission des Bundesministeriums für Land- und Forstwirtschaft, Sorten Gärtnerischer Kulturpflanzen, Vienna, Austria, 20 December 1988; p. 9. [Google Scholar]
- Lapinskas, P.G.W.; Evans, A.M. Growth habit studies in Phaseolus coccineus. Annu. Rep. Bean Improv. Coop. 1977, 20, 46–47. [Google Scholar]
- González, A.M.; De Ron, A.M.; Lores, M.; Santalla, M. Effect of the inbreeding depression in progeny fitness of runner bean (Phaseolus coccineus L.) and it is implications for breeding. Euphytica 2014, 200, 413–428. [Google Scholar] [CrossRef] [Green Version]
- van Hintum, T.J.L. Duplication within and between germplasm collections. III. A quantitative model. Genet. Resour. Crop Evol. 2000, 47, 507–513. [Google Scholar] [CrossRef]
- van Hintum, T.J.L.; Visser, D.L. Duplication within and between germplasm collections. Genet. Resour. Crop Evol. 1995, 42, 135–145. [Google Scholar] [CrossRef]
- Koltowski, Z. Flowering biology, nectar secretion and insect foraging of the runner bean (Phaseolus coccineus L.). J. Apic. Sci. 2004, 48, 53–60. [Google Scholar]
- Goswami, S.; Kumar, R.R.; Sharma, S.K.; Kala, Y.K.; Singh, K.; Gupta, R.; Dhavan, G.; Rai, G.K.; Singh, G.P.; Pathak, H.; et al. Calcium triggers protein kinases-induced signal transduction for augmenting the thermotolerance of developing wheat (Triticum aestivum) grain under the heat stress. J. Plant Biochem. Biotechnol. 2015, 24, 441–452. [Google Scholar] [CrossRef]
- Larkindale, J.; Knight, M.R. Protection against heat stress-induced oxidative damage in Arabidopsis involves calcium, abscisic acid, ethylene, and salicylic acid. Plant Physiol. 2002, 128, 682–695. [Google Scholar] [CrossRef]
- Wang, M.; Zhang, X.; Li, Q.; Chen, X.; Li, X. Comparative transcriptome analysis to elucidate the enhanced thermotolerance of tea plants (Camellia sinensis) treated with exogenous calcium. Planta 2019, 249, 775–786. [Google Scholar] [CrossRef]
- Wang, X.; Huang, B. Lipid- and calcium-signaling regulation of HsfA2c-mediated heat tolerance in tall fescue. Environ. Exp. Bot. 2017, 136, 59–67. [Google Scholar] [CrossRef]
- Naeem, M.; Traub, J.R.; Athar, H.-u.-R.; Loescher, W. Exogenous calcium mitigates heat stress effects in common bean: A coordinated impact of photoprotection of PSII, up-regulating antioxidants, and carbohydrate metabolism. Acta Physiol. Plant. 2020, 42, 180. [Google Scholar] [CrossRef]
- Qiao, B.; Zhang, Q.; Liu, D.; Wang, H.; Yin, J.; Wang, R.; He, M.; Cui, M.; Shang, Z.; Wang, D.; et al. A calcium-binding protein, rice annexin OsANN1, enhances heat stress tolerance by modulating the production of H2O2. J. Exp. Bot. 2015, 66, 5853–5866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jisha, K.; Shackira, A.; Puthur, J. GABA/BABA priming causes signaling of defense pathways related to abiotic stress tolerance in plants. In Neurotransmitters in Plants; CRC Press: Boca Raton, FL, USA, 2018; pp. 219–230. [Google Scholar] [CrossRef]
- Baranašić, J.; Mihalak, A.; Gruić-Sovulj, I.; Bauer, N.; Rokov-Plavec, J. Expression of genes for selected plant aminoacyl-tRNA synthetases in the abiotic stress. Acta Bot. Croat. 2021, 80, 35–42. [Google Scholar] [CrossRef]
- Huang, R.; Liu, Z.; Xing, M.; Yang, Y.; Wu, X.; Liu, H.; Liang, W. Heat stress suppresses Brassica napus seed oil accumulation by inhibition of photosynthesis and BnWRI1 pathway. Plant Cell Physiol. 2019, 60, 1457–1470. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Wen, X.; Lu, Q.; Yang, Z.; Cheng, Z.; Lu, C. Heat stress induces an aggregation of the light-harvesting complex of photosystem II in spinach plants. Plant Physiol. 2007, 143, 629–638. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Hu, T.; Amombo, E.; Wang, G.; Xie, Y.; Fu, J. The alleviation of heat damage to photosystem II and enzymatic antioxidants by exogenous spermidine in tall fescue. Front. Plant Sci. 2017, 8, 1747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Q.; Chen, W.; Bian, J.; Xie, H.; Li, Y.; Xu, C.; Ma, J.; Guo, S.; Chen, J.; Cai, X.; et al. Proteomics and phosphoproteomics of heat stress-responsive mechanisms in spinach. Front. Plant Sci. 2018, 9, 800. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait Group | Trait-ID | Trait Explanation | Trial 2018 | Trial 2020 | GWAS II | GWAS I |
|---|---|---|---|---|---|---|
| Flowers | %_F_Ind | Percentage of individuals of one genotype that flower | NA | 31 | NA | NA |
| Total_MaxF | The highest number of flowers observed on one day over the course of the trial | 29 | 18 | 3 | 0 | |
| Total_NoF | The sum of all flowers counted over the course of the trial | 27 | 18 | 5 | 0 | |
| Minipods | Final_NoMP | The final number of mini pods (<5 cm) harvested at the end of the trial | NA | 32 | NA | NA |
| Total_MaxMP | The highest number of mini pods (<5 cm) observed on one day over the course of the trial | NA | 36 | NA | NA | |
| Pods | %_P_Ind | Percentage of individuals of one genotype that had formed pods (>5 cm) | NA | 45 | NA | NA |
| Final_NoP | The final number of pods (>5 cm) harvested at the end of the trial | 18 | 48 | 3 | 6 | |
| Total_MaxP | The highest number of pods (>5 cm) observed on one day over the course of the trial | 15 | 48 | 1 | 0 | |
| Beans | NoB_per_P | The average number of beans per pod (>5 cm) harvested at the end of the trial | 20 | 21 | 2 | NA |
| Total_NoB | The total number of beans harvested at the end of the trial | 20 | 62 | 1 | 1 | |
| W_B | The weight of all beans harvested for all individuals of one genotype | 19 | 52 | 1 | NA | |
| W_per_B | The average weight per bean harvested at the end of the trial | 87 | 33 | 13 | 1 | |
| Total associations (number of SNPs) | 235 (148) | 444 (182) | 29 (22) | 8 (7) |
| SNP | GWAS | Associated Traits 2018 | Associated Traits 2020 | Alleles | BLAST | Protein | Molecular Function | Biological Function |
|---|---|---|---|---|---|---|---|---|
| RAD_220_83 | II | W_per_B | %_P_Ind, Final_NoP, Total_MaxP, NoB_per_P, Total_NoB, W_B | C/G | Vigna angularis var. angularis DNA, chromosome 10, Range 1: 3,212,827 to 3,212,913 | |||
| RAD_414_35 | II | Final_NoP, Total_MaxP, NoB_per_P, Total_NoB, W_B | %_P_Ind, Final_NoP, Total_MaxP, NoB_per_P, Total_NoB, W_B | C/G | PHAVU_001G030000g | aspartyl-tRNA synthetase | aspartate-tRNA ligase activity; ATP binding; DNA binding | aspartyl-tRNA aminoacylation |
| RAD_611_31 | II | W_per_B | W_per_B | T/C | PHAVU_005G108600g | uncharacterized protein | photosystem II assembly | |
| RAD_1199_80 | II | NoB_per_P | %_F_Ind | T/A | PHAVU_008G292100g | uncharacterized protein | calcium ion binding | |
| RAD_1289_78 | II | Total_NoF, NoB_per_P, W_per_B | W_per_B | A/C | Vigna unguiculata cultivar Xiabao 2 chromosome Vu03, Range 1: 70,689,604 to 70,689,680 | |||
| RAD_1340_7 | II | NoB_per_P | Total_MaxMP, Final_NoP, Total_MaxP, Total_NoB | A/G | Vigna unguiculata cultivar Xiabao 2 chromosome Vu04, Range 1: 7,129,018 to 7,129,088 | |||
| RAD_1525_41 | II | W_per_B | Final_NoMP, Total_MaxMP | A/C | PHAVU_009G248400g | uncharacterized protein | ATPase-coupled transmembrane transporter activity; ATP binding | |
| RAD_1534_20 | II | Total_MaxF, W_per_B | %_F_Ind, Total_MaxF, Total_NoF, Final_NoMP, Total_MaxMP, %_P_Ind, Final_NoP, Total_MaxP, NoB_per_P, Total_NoB, W_B, W_per_B | T/A | PHAVU_007G000900g | Exocyst subunit Exo70 family protein | exocytosis; protein transport | |
| RAD_2111_42 | II | Total_NoF | %_F_Ind | T/A | Vigna unguiculata cultivar Xiabao 2 chromosome Vu11, Range 1: 4,515,333 to 4,515,417 | |||
| RAD_2341_68 | II | Total_MaxF, Total_NoF, Final_NoP, W_B | %_F_Ind, Total_MaxF, Total_NoF, Final_NoP | C/T | PHAVU_010G032700g | protein transport protein SEC23 | zinc ion binding | COPII-coated vesicle budding; intracellular protein transport |
| RAD_2589_39 | II | W_per_B | Final_NoMP | T/C | Vigna angularis var. angularis DNA, chromosome 8, Range 1: 13,616,071 to 13,616,149 | |||
| RAD_3146_19 | II | NoB_per_P, W_per_B | Total_MaxF, Total_NoF, Total_MaxMP | C/T | PHAVU_003G137000g | beta-galactosidase | beta-galactosidase activity; carbohydrate binding | carbohydrate metabolic process |
| RAD_3232_18 | II | W_per_B | %_P_Ind, Final_NoP, Total_MaxP | T/C | Vigna unguiculata cultivar Xiabao 2 chromosome LG9, Range 1: 25,868,642 to 25,868,730 | |||
| RAD_3235_24 | II | W_B | Final_NoMP, Total_MaxMP | T/C | Vigna unguiculata vacuolar protein sorting-associated protein 54, chloroplastic (LOC114163195) | vacuolar protein sorting-associated protein 54 | endocytic recycling; retrograde transport, endosome to Golgi | |
| RAD_3342_71 | II | Total_MaxF, Total_NoF | %_F_Ind | G/T | Vigna unguiculata cultivar Xiabao 2 chromosome Vu03, Range 1: 80,449,805 to 80,449,873 | |||
| RAD_3432_30 | I | W_per_B | NA | A/G | Vigna angularis var. angularis DNA, chromosome 8, Range 1: 14,570,377 to 14,570,462 | |||
| RAD_3517_34 | I | Final_NoP | NA | C/T | PHAVU_001G190500g | uncharacterized protein | ||
| RAD_4014_50 | II | W_per_B | %_F_Ind | G/C | PHAVU_002G205700g | uncharacterized protein | defense response to fungus |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bomers, S.; Sehr, E.M.; Adam, E.; von Gehren, P.; Hansel-Hohl, K.; Prat, N.; Ribarits, A. Towards Heat Tolerant Runner Bean (Phaseolus coccineus L.) by Utilizing Plant Genetic Resources. Agronomy 2022, 12, 612. https://doi.org/10.3390/agronomy12030612
Bomers S, Sehr EM, Adam E, von Gehren P, Hansel-Hohl K, Prat N, Ribarits A. Towards Heat Tolerant Runner Bean (Phaseolus coccineus L.) by Utilizing Plant Genetic Resources. Agronomy. 2022; 12(3):612. https://doi.org/10.3390/agronomy12030612
Chicago/Turabian StyleBomers, Svenja, Eva M. Sehr, Eveline Adam, Philipp von Gehren, Karin Hansel-Hohl, Noémie Prat, and Alexandra Ribarits. 2022. "Towards Heat Tolerant Runner Bean (Phaseolus coccineus L.) by Utilizing Plant Genetic Resources" Agronomy 12, no. 3: 612. https://doi.org/10.3390/agronomy12030612