
Comparative Analysis of the Effects of Plastic Mulch Films on Soil Nutrient, Yields and Soil Microbiome in Three Vegetable Fields

Abstract
:1. Introduction
2. Material and Methods
2.1. Soil Sample Collection
2.2. Determination of Soil Properties
2.3. Sequences Processing and Analysis
2.4. Diversity Analysis
2.5. Statistical Analysis
3. Results
3.1. Effects of Different Covered Plastic Mulch Films on the Properties of Rhizosphere and Non-Rhizosphere Soils
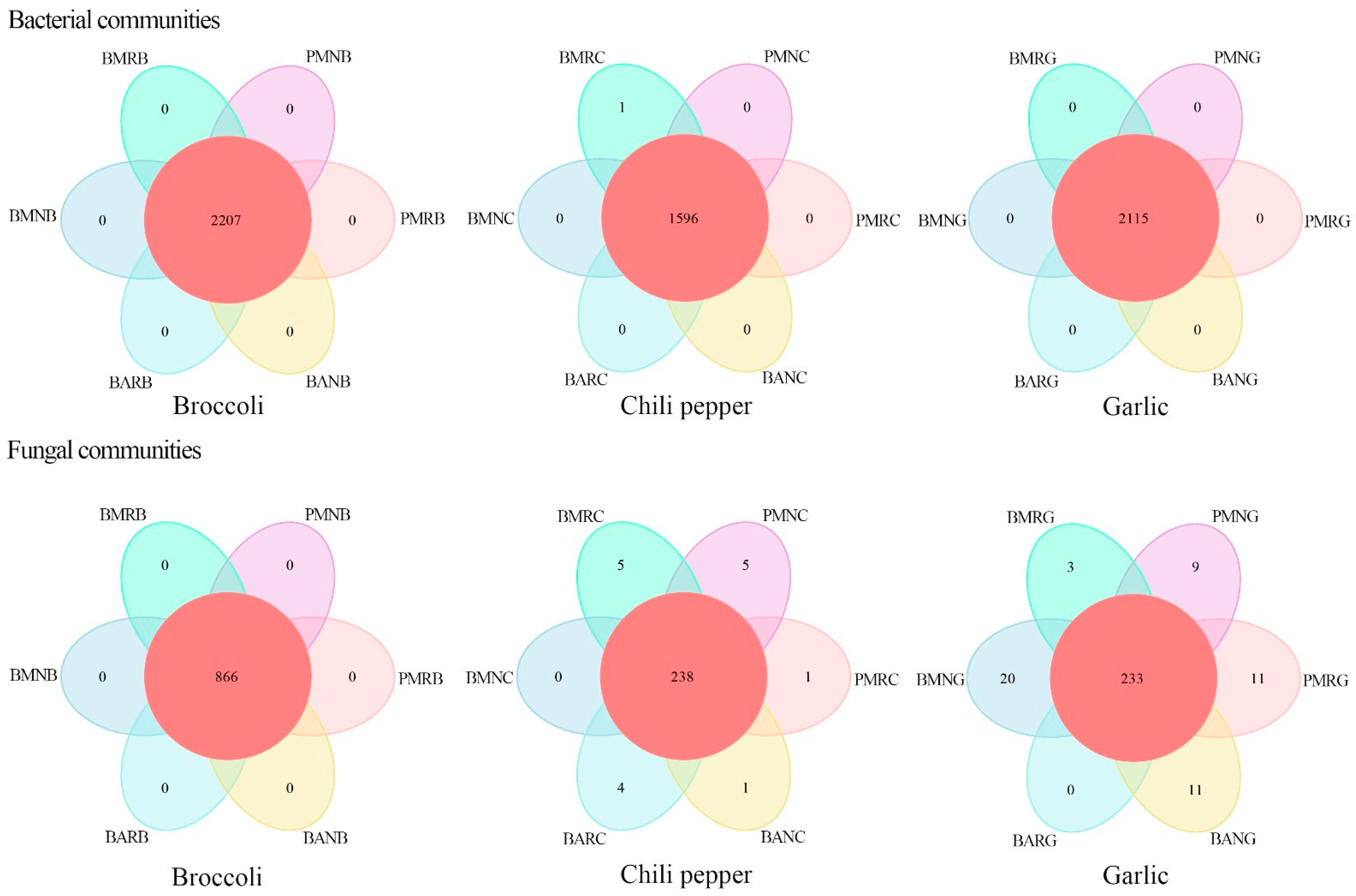
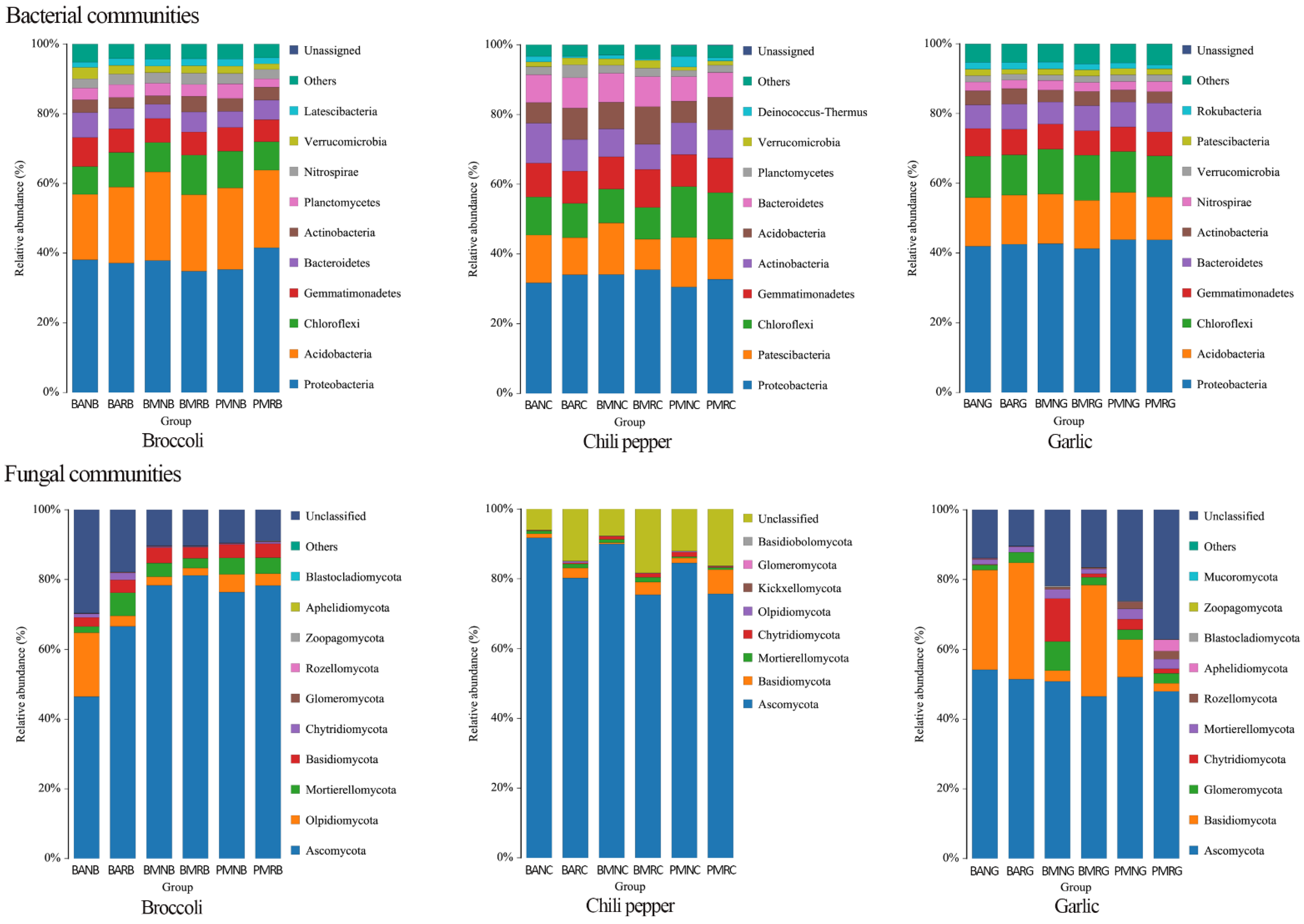
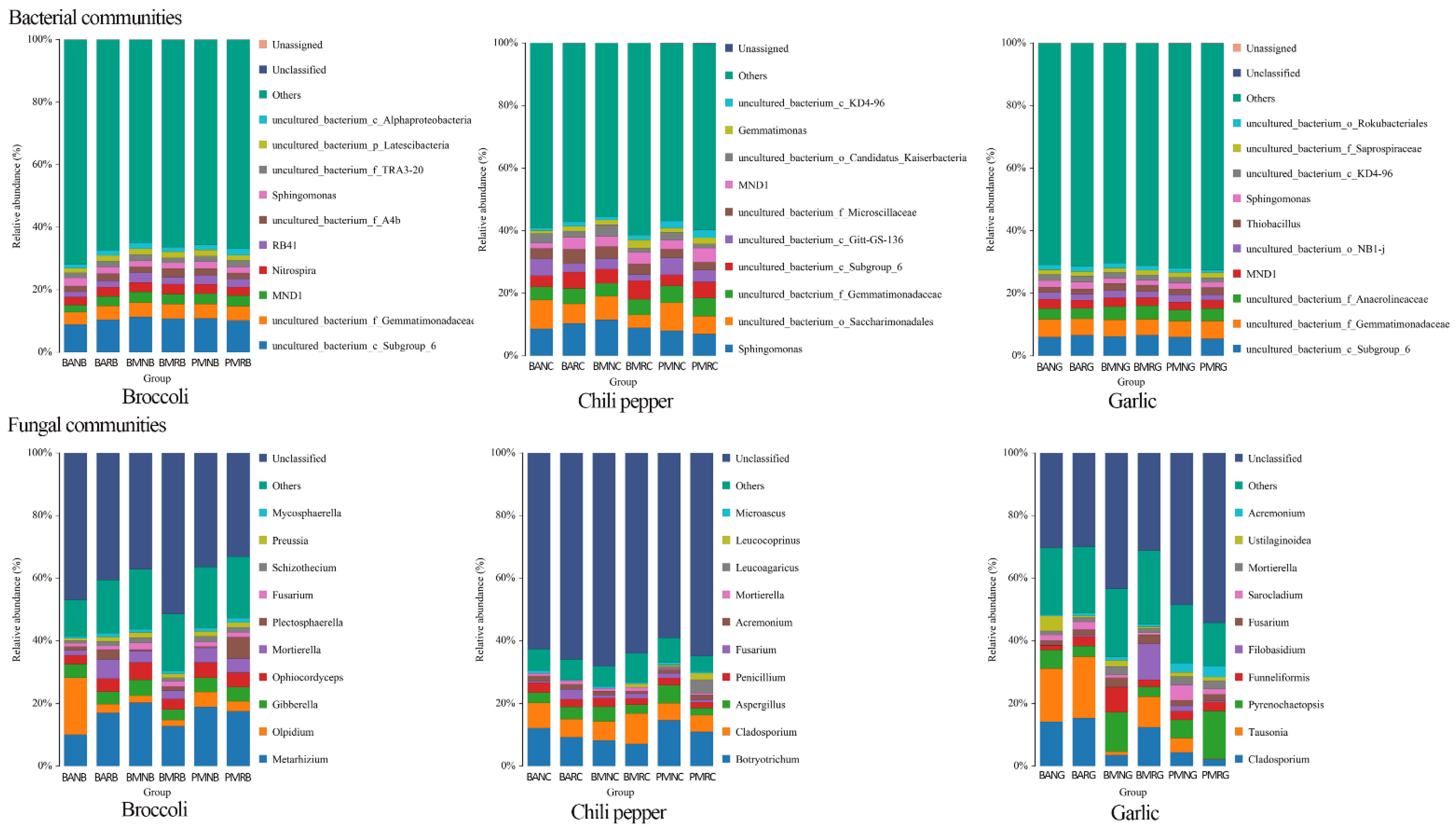
3.2. Soil Microbiome Analysis
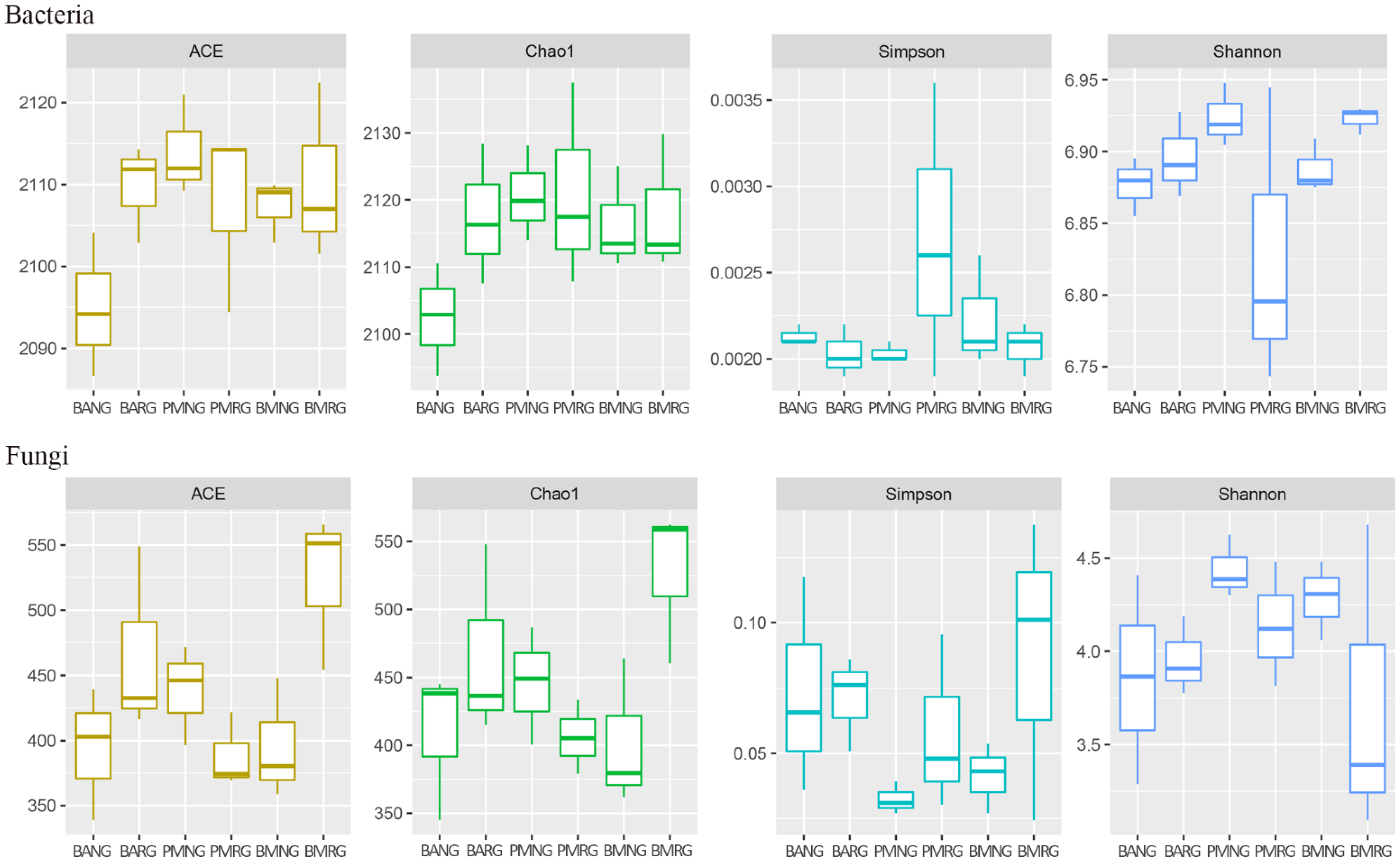
3.3. α-Diversity Comparison among Different Samples
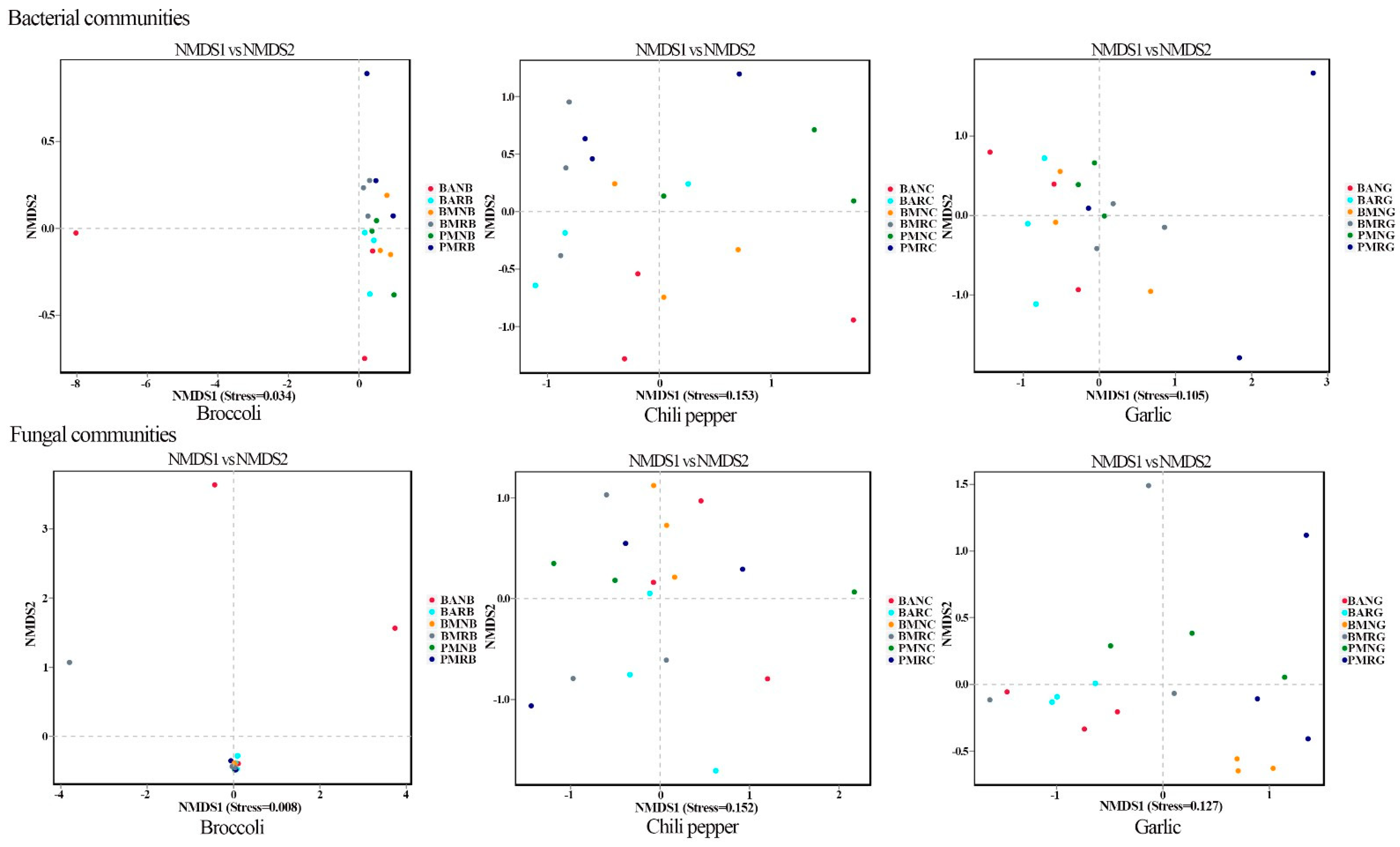
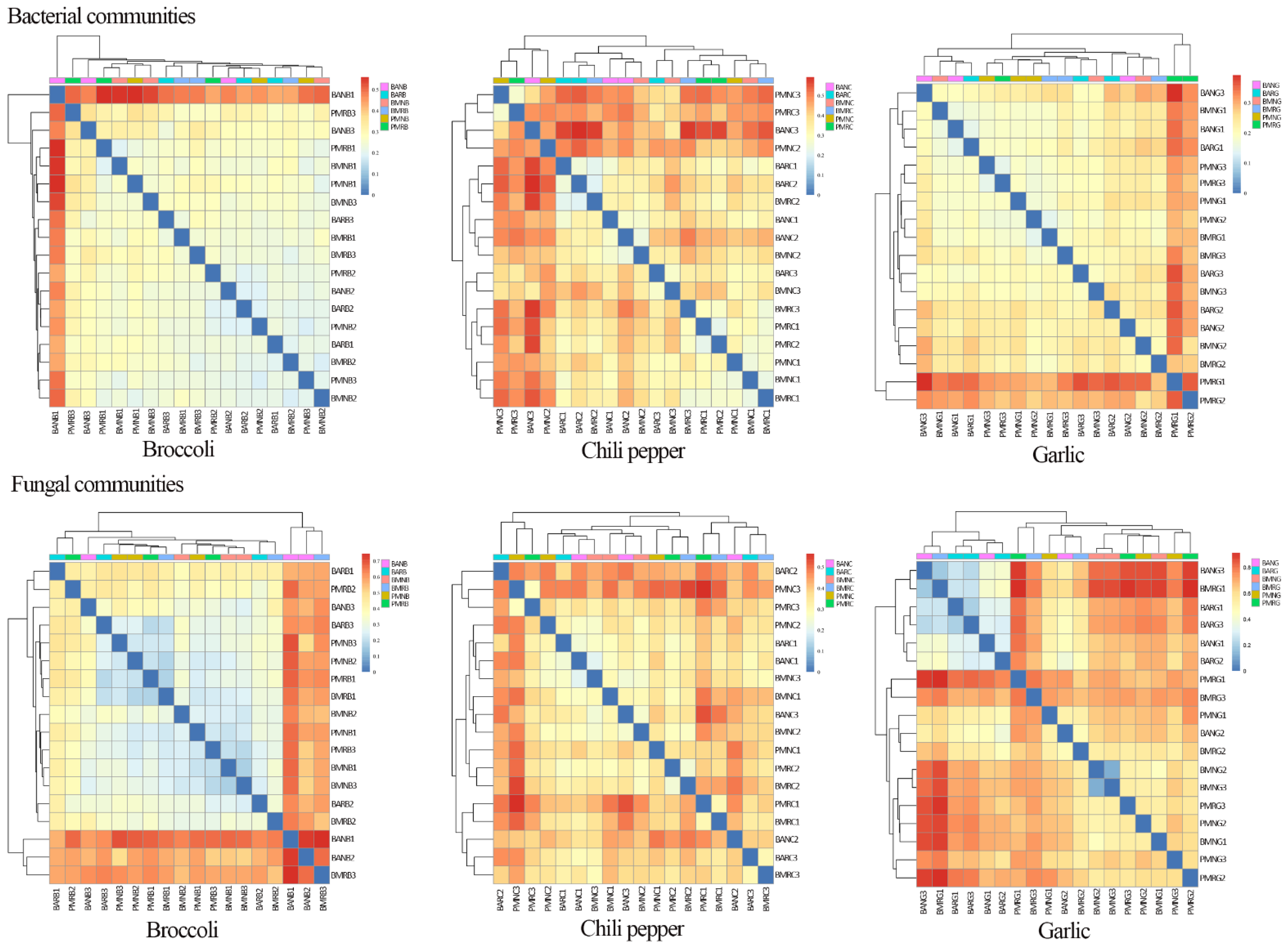
3.4. β-Diversity Comparison among Different Samples
3.5. Effects of Mulch Films on Rhizosphere and Non-Rhizosphere Soil Microbial Community Composition
3.6. Effects of Mulch Films on Crop Yields
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kasirajan, S.; Ngouajio, M. Polyethylene and biodegradable mulches for agricultural applications: A review. Agron. Sustain. Dev. 2012, 32, 501–529. [Google Scholar] [CrossRef]
- Kader, M.A.; Senge, M.; Mojid, M.A.; Ito, K. Recent advances in mulching materials and methods for modifying soil environment. Soil Tillage Res. 2017, 168, 155–166. [Google Scholar] [CrossRef]
- Martín-Closas, L.; Costa, J.; Pelacho, A.M. Agronomic effects of biodegradable films on crop and field environment. In Soil Degradable Bioplastics for a Sustainable Modern Agriculture; Malinconico, M., Ed.; Springer: Berlin/Heidelberg, Germany, 2017; pp. 67–104. [Google Scholar]
- Ren, X.L.; Zhang, P.; Chen, X.L.; Jia, Z.K. Impacts of ridge-furrow rainfall concentration systems and mulches on corn growth and yield in the semiarid region of China. J. Sci. Food Agric. 2016, 96, 3882–3889. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.L.; Zhang, P.; Chen, X.L.; Guo, J.J.; Jia, Z.K. Effect of Different Mulches under Rainfall Concentration System on Corn Production in the Semi-arid Areas of the Loess Plateau. Sci. Rep. 2016, 6, 19019. [Google Scholar] [CrossRef] [Green Version]
- Braunack, M.V.; Adhikari, R.; Freischmidt, G.; Johnston, P.; Filipovi, V. Initial experimental experience with a sprayable biodegradable polymer membrane (SBPM) technology in cotton. Agronomy 2020, 10, 584. [Google Scholar] [CrossRef] [Green Version]
- Abel, D.; Kloas, W.; Zarfl, C.; Stefan, H.; Matthias, C. Microplastics as an emerging threat to terrestrial ecosystems. Glob. Chang. Biol. 2018, 24, 1405–1416. [Google Scholar]
- Qian, H.; Meng, Z.; Liu, G.F.; Lu, T.; Qu, Q.; Du, B.; Pan, X. Effects of Soil Residual Plastic Film on Soil Microbial Community Structure and Fertility. Water Air Soil Pollut. 2018, 229, 261. [Google Scholar] [CrossRef]
- Steinmetz, Z.; Wollmann, C.; Schaefer, M.; Buchmann, C.; David, J.; Troger, J. Plastic mulching in agriculture. Trading short-term agronomic benefits for long-term soil degradation? Sci. Total Environ. 2016, 550, 690–705. [Google Scholar] [CrossRef]
- Sreejata, B.; Lluis, M.C.; Pelacho, A.M. Biodegradable Plastic Mulch Films: Impacts on Soil Microbial Communities and Ecosystem Functions. Front. Microbiol. 2018, 9, 819. [Google Scholar]
- Marechal, F. Biodegradable plastics. In Biodegradable Polymers and Plastics; Chiellini, E., Solaro, R., Eds.; Springer: New York, NY, USA, 2003. [Google Scholar]
- Brodhagen, M.; Peyron, M.; Miles, C.; Inglis, D.A. Biodegradable plastic agricultural mulches and key features of microbial degradation. Appl. Microbiol. Biotechnol. 2015, 99, 1039–1056. [Google Scholar] [CrossRef]
- Kyrikou, I.; Briassoulis, D. Biodegradation of agricultural plastic films: A critical review. J. Polym. Environ. 2007, 15, 125–150. [Google Scholar] [CrossRef]
- Li, C.; Moore-Kucera, J.; Lee, J.; Corbin, A.; Brodhagen, M.; Miles, C. Effects of biodegradable mulch on soil quality. Appl. Soil Ecol. 2014, 79, 59–69. [Google Scholar] [CrossRef]
- Yamamoto-Tamura, K.; Hiradate, S.; Watanabe, T.; Koitabashi, M.; Sameshima-Yamashita, Y.; Yarimizu, T. Contribution of soil esterase to biodegradation of aliphatic polyester agricultural mulch film in cultivated soils. AMB Express 2015, 5, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koitabashi, M.; Noguchi, M.; Sameshima-Yamashita, Y.; Hiradate, S.; Suzuki, K. Degradation of biodegradable plastic mulch films in soil environment by phylloplane fungi isolated from gramineous plants. AMB Express 2012, 2, 40. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Moore-Kucera, J.; Miles, C.; Leonas, K.; Lee, J.; Corbin, A. Degradation of potentially biodegradable plastic mulch films at three diverse US locations. Agroecol. Sustain. Food Syst. 2014, 38, 861–889. [Google Scholar] [CrossRef]
- Muroi, F.; Tachibana, Y.; Kobayashi, Y.; Sakurai, T.; Kasuya, K. Influences of poly(butylene adipate-co-terephthalate) on soil microbiota and plant growth. Polym. Degrad. Stab. 2016, 129, 338–346. [Google Scholar] [CrossRef]
- Rychter, P.; Biczak, R.; Herman, B.; Smylla, A.; Kurcok, P.; Adamus, G. Environmental degradation of polyester blends containing atactic poly(3-hydroxybutyrate). Biodegradation in soil and ecotoxicological impact. Biomacromolecules 2006, 7, 3125–3131. [Google Scholar] [CrossRef]
- Ma, Z.; Ma, Y.; Qin, L.; Liu, J.; Su, H. Preparation and characteristics of biodegradable mulching films based on fermentation industry wastes. Int. Biodeterior. Biodegrad. 2016, 111, 54–61. [Google Scholar] [CrossRef]
- Qin, W.; Hu, C.; Oenema, O. Soil mulching significantly enhances yields and water and nitrogen use efficiencies of maize and wheat: A meta-analysis. Sci. Rep. 2015, 5, 16210. [Google Scholar] [CrossRef]
- Nan, W.; Yue, S.; Huang, H.; Li, S.; Shen, Y. Effects of plastic film mulching on soil greenhouse gases (CO2, CH4 and N2O) concentration within soil profiles in maize fields on the Loess Plateau, China. J. Integr. Agric. 2016, 15, 451–464. [Google Scholar] [CrossRef] [Green Version]
- Lu, R.K. The Analysis Method of Soil Agricultural Chemistry; China Agricultural Science and Technology Press: Beijing, China, 2000; pp. 146–190. [Google Scholar]
- Edgar, R. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Koljalg, U.; Nilsson, R.; Abarenkov, K.; Tedersoo, L.; Taylor, A.; Bahram, M.; Bates, S. Towards a unified paradigm for sequence-based identification of fungi. Mol. Ecol. 2013, 22, 5271–5277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Garrity, G.; Tiedje, J.; Cole, J. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chao, A. Nonparametric estimation of the number of classes in a population. Scand. J. Stat. 1984, 11, 265–270. [Google Scholar]
- Zhou, D.; Feng, H.; Schuelke, T.; De Santiago, A.; Zhang, Q.; Zhang, J. Rhizosphere microbiomes from root knot nematode non-infested plants suppress nematode infection. Microb. Ecol. 2019, 78, 470–481. [Google Scholar] [CrossRef] [Green Version]
- Looft, T.; Johnson, T.; Allen, H. In-feed antibiotic effects on the swine intestinal microbiome. Proc. Natl. Acad. Sci. USA 2012, 109, 1691–1696. [Google Scholar] [CrossRef] [Green Version]
- Qiao, Q.; Wang, F.; Zhang, J.; Chen, Y.; Zhang, C.; Liu, G. The variation in the rhizosphere microbiome of cotton with soil type, genotype and developmental stage. Sci. Rep. 2017, 7, 3940. [Google Scholar] [CrossRef] [Green Version]
- Sreejata, B.; González, J.; Henderson, K.; Anunciado, M.; Hayes, D.; DeBruyn, J. Soil Microbial Communities Associated With Biodegradable Plastic Mulch Films. Front. Microbiol. 2020, 11, 587074. [Google Scholar]
- Jordan, A.; Zavala, L.; Munoz-Rojas, M. Mulching, effects on soil physical properties. In Encyclopedia of Earth Sciences; Springer: Dordrecht, The Netherlands, 2011; pp. 492–496. [Google Scholar]
- Kapanen, A.; Schettini, E.; Giuliano, V.; Itävaara, M. Performance and environmental impact of biodegradable films in agriculture: A field study on protected cultivation. J. Polym. Environ. 2008, 16, 109–122. [Google Scholar] [CrossRef]
- Moreno, M.; Moreno, A. Effect of different biodegradable and polyethylene mulches on soil properties and production in a tomato crop. Sci. Hortic. 2008, 116, 256–263. [Google Scholar] [CrossRef]
- Bowen, P.; Frey, B. Response of plasticultured bell pepper to staking, irrigation frequency, and fertigated nitrogen rate. Hort. Sci. 2002, 37, 95–100. [Google Scholar] [CrossRef] [Green Version]
- Romic, D.; Romic, M.; Borosic, J. Mulching decreases nitrate leaching in bell pepper cultivation. Agric. Water Manag. 2003, 60, 87–97. [Google Scholar] [CrossRef] [Green Version]
- Sintim, H.; Bandopadhyay, S.; English, M.; Bary, A.; DeBruyn, J. Impacts of biodegradable plastic mulches on soil health. Agric. Ecosyst. Environ. 2019, 273, 36–49. [Google Scholar] [CrossRef]
- Zuber, S.; Behnke, G.; Nafziger, E.; Villamil, M. Crop rotation and tillage effects on soil physical and chemical properties in Illinois. Agron. J. 2015, 107, 971–978. [Google Scholar] [CrossRef] [Green Version]
- Alam, M.; Islam, M.; Salahin, N.; Hasanuzzaman, M. Effect of tillage practices on soil properties and crop productivity in wheat–mungbean–rice cropping system under subtropical climatic conditions. Sci. World J. 2014, 2014, 437283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Busari, M.; Kukal, S.; Kaur, A.; Bhatt, R.; Dulazi, A. Conservation tillage impacts on soil, crop and the environment. Int. Soil Water Conserv. Res. 2015, 3, 119–129. [Google Scholar] [CrossRef] [Green Version]
- Farmer, J.; Zhang, B.; Jin, X.; Zhang, P.; Wang, J. Long-term effect of plastic film mulching and fertilization on bacterial communities in a brown soil revealed by high through-put sequencing. Arch. Agron. Soil Sci. 2017, 63, 230–241. [Google Scholar] [CrossRef]
- Munoz, K.; Schmidt-Heydt, M.; Stoll, D.; Diehl, D.; Ziegler, J.; Geisen, R.; Schaumann, G.E. Effect of plastic mulching on mycotoxin occurrence and mycobiome abundance in soil samples from asparagus crops. Mycotoxin Res. 2015, 31, 191–201. [Google Scholar]
- Lundberg, D.; Lebeis, S.; Paredes, S.; Yourstone, S.; Gehring, J.; Malfatti, S. Defining the core Arabidopsis thaliana root microbiome. Nature 2012, 488, 86–90. [Google Scholar] [CrossRef] [Green Version]
- Bulgarelli, D.; Garrido-Oter, R.; Münch, P.; Weiman, A.; Dröge, J.; Pan, Y. Structure and function of the bacterial root microbiota in wild and domesticated barley. Cell Host Microbe 2015, 17, 392–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendes, R.; Garbeva, P.; Raaijmakers, J. The rhizosphere microbiome: Significance of plant beneficial, plant pathogenic and human pathogenic microorganisms. FEMS Microbiol. Rev. 2013, 37, 634–663. [Google Scholar] [CrossRef] [PubMed]
- Weinert, N.; Piceno, Y.; Ding, G.; Meincke, R.; Heuer, H.; Berg, G. PhyloChip hybridization uncovered an enormous bacterial diversity in the rhizosphere of different potato cultivars: Many common and few cultivar-dependent taxa. FEMS Microbiol. Ecol. 2011, 75, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Pathak, V.M.; Navneet. Review on the current status of polymer degradation: A microbial approach. Bioresour. Bioprocess. 2017, 4, 15. [Google Scholar] [CrossRef]
- Xu, M.; Xian, Y.; Wu, J.; Gu, Y.; Yang, G.; Zhang, X. Effect of biogas slurry addition on soil properties, yields, and bacterial composition in the rice-rape rotation ecosystem over 3 years. J. Soils Sediments 2019, 19, 2534–2542. [Google Scholar] [CrossRef]
- Souza, R.; Mendes, I.; Reis-Junior, F.; Carvalho, F.; Nogueira, M.; Vasconcelos, A.; Vicente, V.A.; Hungria, M. Shifts in taxonomic and functional microbial diversity with agriculture: How fragile is the Brazilian Cerrado? BMC Microbiol. 2016, 16, 42. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Liang, Y.; Jiang, Y. Planting increases the abundance and structure complexity of soil core functional genes relevant to carbon and nitrogen cycling. Sci. Rep. 2015, 5, 14345. [Google Scholar] [CrossRef] [Green Version]
- Sun, Q.; Rui, W.; Hu, Y.; Yao, L.; Guo, S.; Jorge, P. Spatial variations of soil respiration and temperature sensitivity along a steep slope of the semiarid Loess Plateau. PLoS ONE 2018, 13, e0195400. [Google Scholar] [CrossRef]
- Procter, A.; Ellis, J.; Fay, P.; Polley, H.; Jackson, R. Fungal community responses to past and future atmospheric CO2 differ by soil type. App. Environ. Microbiol. 2014, 80, 7364–7377. [Google Scholar] [CrossRef] [Green Version]
- Bandopadhyay, S.; Sintim, H.Y.; DeBruyn, J.M. Structural and Functional Responses of Soil Microbial Communities to Biodegradable Plastic Film Mulching in Two Agroecosystems. BioRxiv 2019, 5, 650317. [Google Scholar]
- Deng, L.; Yu, Y.; Zhang, H.; Wang, Q.; Yu, R. The Effects of Biodegradable Mulch Film on the Growth, Yield, and Water Use Efficiency of Cotton and Maize in an Arid Region. Sustainability 2019, 11, 7039. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vegetables Fields | Sowing Date | Harvesting Date |
|---|---|---|
| Broccoli | January, 2020 | May, 2020 |
| Chili pepper | November, 2019 | May, 2020 |
| Garlic | September, 2019 | May, 2020 |
| Vegetables | Treatments | Organic Matter (g/kg) | Ammonium-N (mg/kg) | Nitrate-N (mg/kg) | Alkaline Hydrolysis-N (mg/kg) | Exchangeable Potassium (mg/kg) | Available Phosphorus (mg/kg) |
|---|---|---|---|---|---|---|---|
| Broccoli | BANB | 11.80 ± 0.72 ab | 1.11 ± 0.20 ab | 8.03 ± 1.30 b | 69.00 ± 7.23 a | 109.33 ± 16.05 a | 44.83 ± 9.58 a |
| BARB | 11.93 ± 0.68 ab | 0.98 ± 0.13 ab | 12.68 ± 1.70 b | 75.67 ± 2.73 a | 99.67 ± 6.01 a | 38.37 ± 4.31 a | |
| BMNB | 13.10 ± 0.45 a | 0.92 ± 0.03 ab | 10.44 ± 3.98 b | 81.67 ± 3.93 a | 127.33 ± 10.81 a | 58.30 ± 13.25 a | |
| BMRB | 12.53 ± 0.18 ab | 1.12 ± 0.06 ab | 31.13 ± 5.90 a | 78.00 ± 2.08 a | 134.67 ± 10.04 a | 53.40 ± 7.21 a | |
| PMNB | 11.23 ± 0.42 b | 0.78 ± 0.70 b | 9.92 ± 1.07 b | 59.33 ± 0.88 a | 103.67 ± 6.89 a | 33.67 ± 3.44 a | |
| PMRB | 11.23 ± 0.45 b | 1.36 ± 0.24 a | 18.51 ± 3.44 b | 70.00 ± 18.04 a | 144.33 ± 32.04 a | 49.40 ± 9.77 a | |
| Chili pepper | BANC | 11.13 ± 0.63 a | 4.14 ± 1.70 a | 17.19 ± 2.31 b | 64.23 ± 2.52 b | 199.83 ± 25.98 a | 66.8 ± 5.67 b |
| BARC | 11.73 ± 0.94 a | 2.55 ± 0.26 ab | 21.05 ± 1.55 ab | 67.97 ± 2.00 ab | 110.337 ± 7.49 b | 70.03 ± 5.66 ab | |
| BMNC | 12.03 ± 1.02 a | 1.21 ± 0.14 ab | 23.35 ± 0.63 ab | 72.70 ± 1.01 ab | 189.47 ± 16.74 a | 72.80 ± 1.36 ab | |
| BMRC | 12.30 ± 0.99 a | 1.01 ± 0.08 b | 27.59 ± 1.91 a | 74.33 ± 3.54 ab | 108.90 ± 2.17 b | 70.43 ± 0.37 ab | |
| PMNC | 13.00 ± 2.51 a | 1.73 ± 0.42 ab | 22.88 ± 3.59 ab | 71.40 ± 6.70 ab | 188.60 ± 22.93 a | 82.57 ± 5.13 a | |
| PMRC | 13.83 ± 2.88 a | 2.19 ± 0.10 ab | 26.62 ± 1.90 a | 78.87 ± 6.28 a | 126.07 ± 22.79 b | 75.03 ± 5.67 ab | |
| Garlic | BANG | 14.37 ± 0.91 b | 2.03 ± 0.22 b | 15.51 ± 2.35 c | 86.20 ± 5.81 a | 196.93 ± 4.23 a | 70.14 ± 4.82 bc |
| BARG | 16.57 ± 0.88 b | 2.74 ± 0.14 a | 16.87 ± 0.96 bc | 87.27 ± 6.46 a | 170.07 ± 3.79 b | 77.87 ± 1.69 ab | |
| BMNG | 15.63 ± 1.70 b | 2.85 ± 0.09 a | 20.63 ± 0.36 ab | 85.17 ± 0.75 a | 170.70 ± 8.31 b | 63.98 ± 4.12 c | |
| BMRG | 15.77 ± 0.38 b | 2.48 ± 0.07 ab | 23.61 ± 0.48 a | 76.80 ± 2.17 a | 138.57 ± 1.79 c | 78.12 ± 2.94 ab | |
| PMNG | 15.67 ± 1.07 b | 2.42 ± 0.28 ab | 22.99 ± 2.06 a | 87.63 ± 6.38 a | 202.50 ± 12.03 a | 71.80 ± 2.81 bc | |
| PMRG | 21.77 ± 1.32 a | 2.52 ± 0.04 ab | 21.62 ± 1.25 a | 85.87 ± 5.24 a | 191.50 ± 9.12 ab | 83.23 ± 1.32 a |
| Treatment Methods | Broccoli | Chili Pepper | Garlic | |
|---|---|---|---|---|
| Yields (kg/hm2) | Bare soil | 12,487.0 ± 292.4 b | 39,605.0 ± 400.4 b | 17,852.5 ± 346.2 b |
| BDMs treatment | 14,686.0 ± 396.6 a | 48,040.0 ± 848.7 a | 22,487.0 ± 388.0 a | |
| PEMs treatment | 14,860.5 ± 310.9 a | 48,825.0 ± 575.3 a | 23,119.5 ± 599.2 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shan, X.; Zhang, W.; Dai, Z.; Li, J.; Mao, W.; Yu, F.; Ma, J.; Wang, S.; Zeng, X. Comparative Analysis of the Effects of Plastic Mulch Films on Soil Nutrient, Yields and Soil Microbiome in Three Vegetable Fields. Agronomy 2022, 12, 506. https://doi.org/10.3390/agronomy12020506
Shan X, Zhang W, Dai Z, Li J, Mao W, Yu F, Ma J, Wang S, Zeng X. Comparative Analysis of the Effects of Plastic Mulch Films on Soil Nutrient, Yields and Soil Microbiome in Three Vegetable Fields. Agronomy. 2022; 12(2):506. https://doi.org/10.3390/agronomy12020506
Chicago/Turabian StyleShan, Xi, Wei Zhang, Zhongliang Dai, Jianbin Li, Weiwei Mao, Fangwei Yu, Jinjun Ma, Shenyun Wang, and Xiaoping Zeng. 2022. "Comparative Analysis of the Effects of Plastic Mulch Films on Soil Nutrient, Yields and Soil Microbiome in Three Vegetable Fields" Agronomy 12, no. 2: 506. https://doi.org/10.3390/agronomy12020506