
Effect of Seaweed Extract Supplement on Rice Rhizosphere Bacterial Community in Tillering and Heading Stages

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
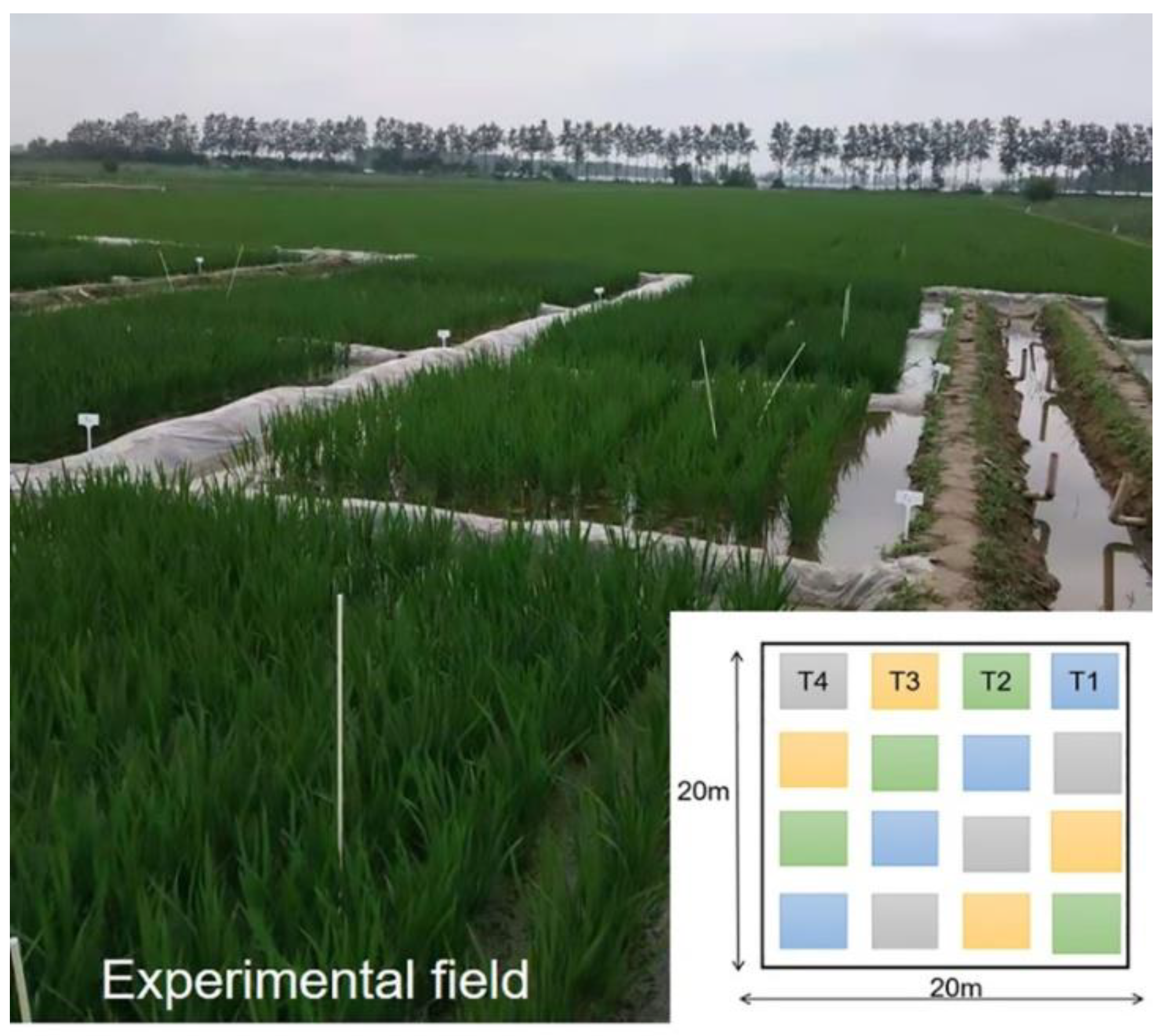
2.1. Sites and Soil Sampling
2.2. Experimental Groups
2.3. Rice Quality
2.4. 16S rRNA Gene Amplicon Sequencing
2.5. Statistical Analyses
3. Results
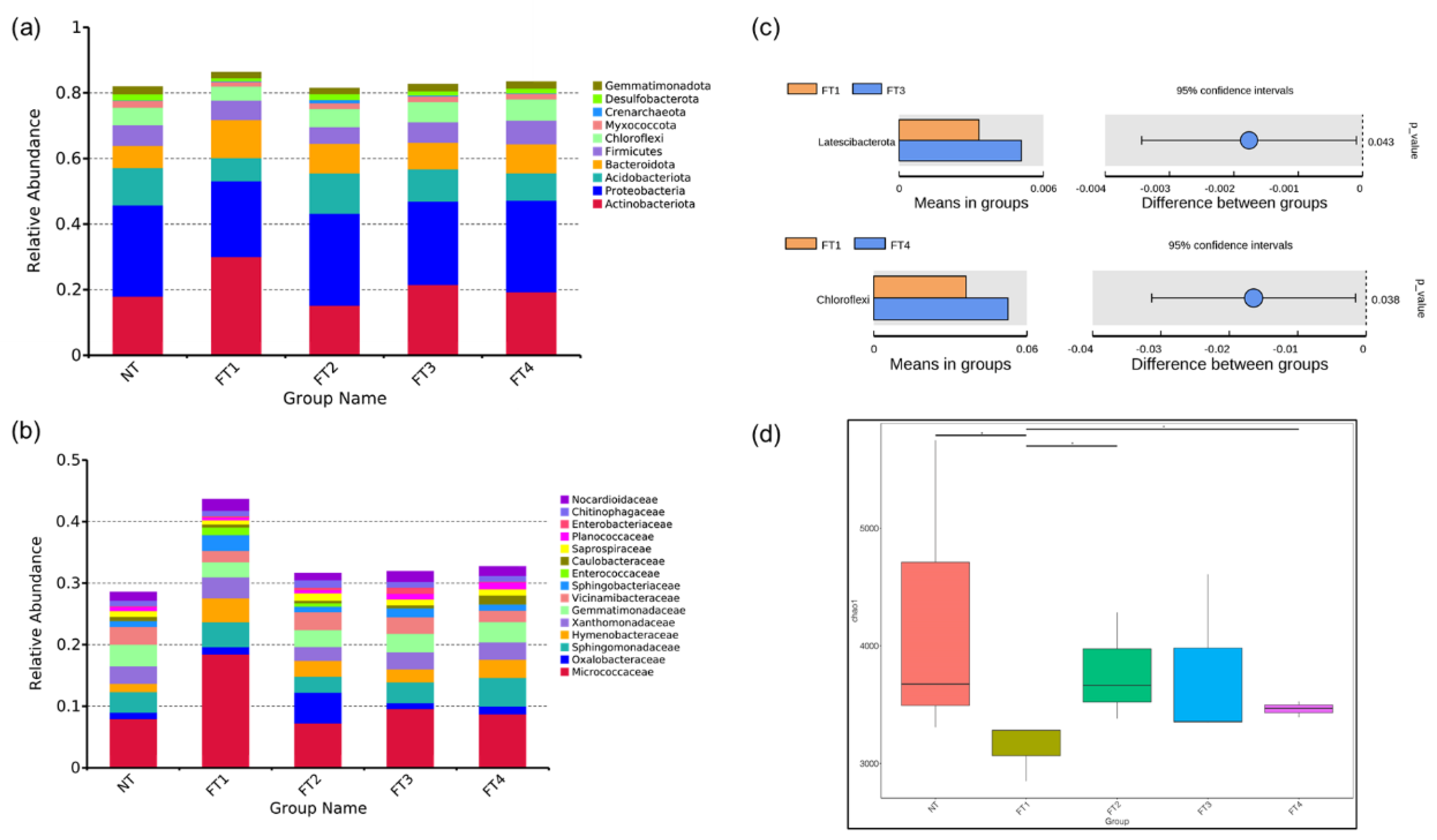
3.1. Changes in Rice Rhizosphere Soil in theTillering Stage
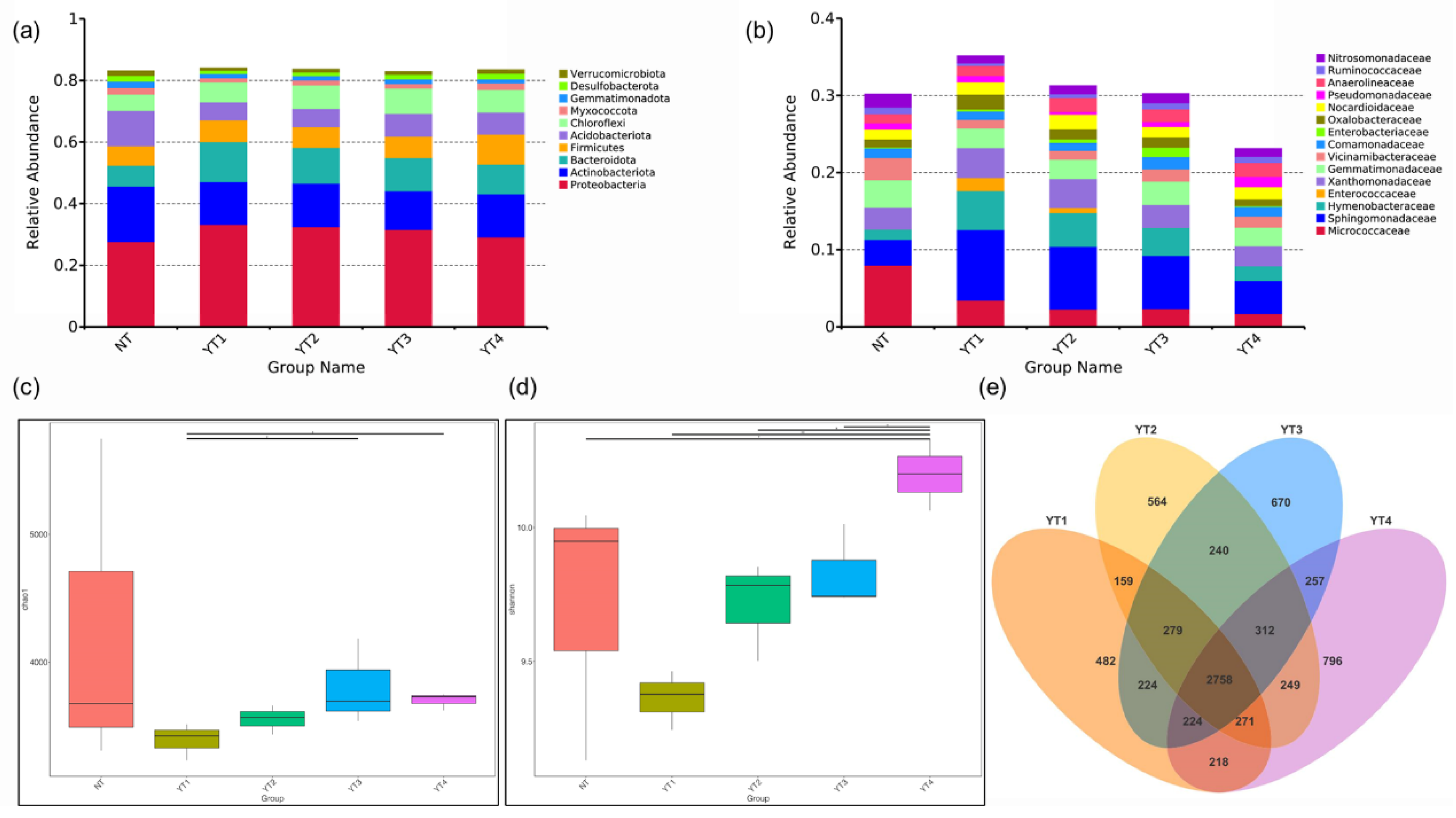
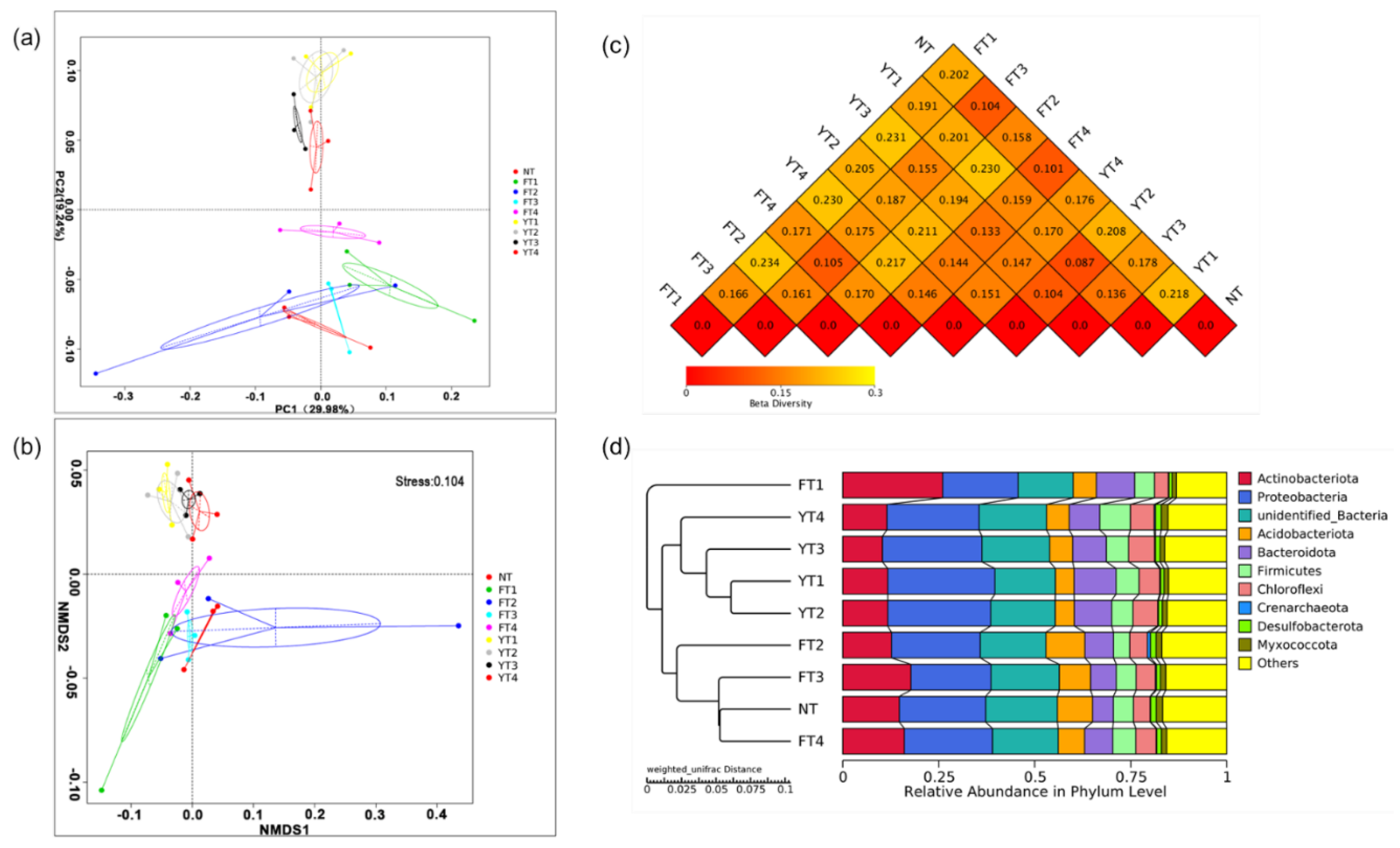
3.2. Changes in Rice Rhizosphere Soil in the Heading Stage
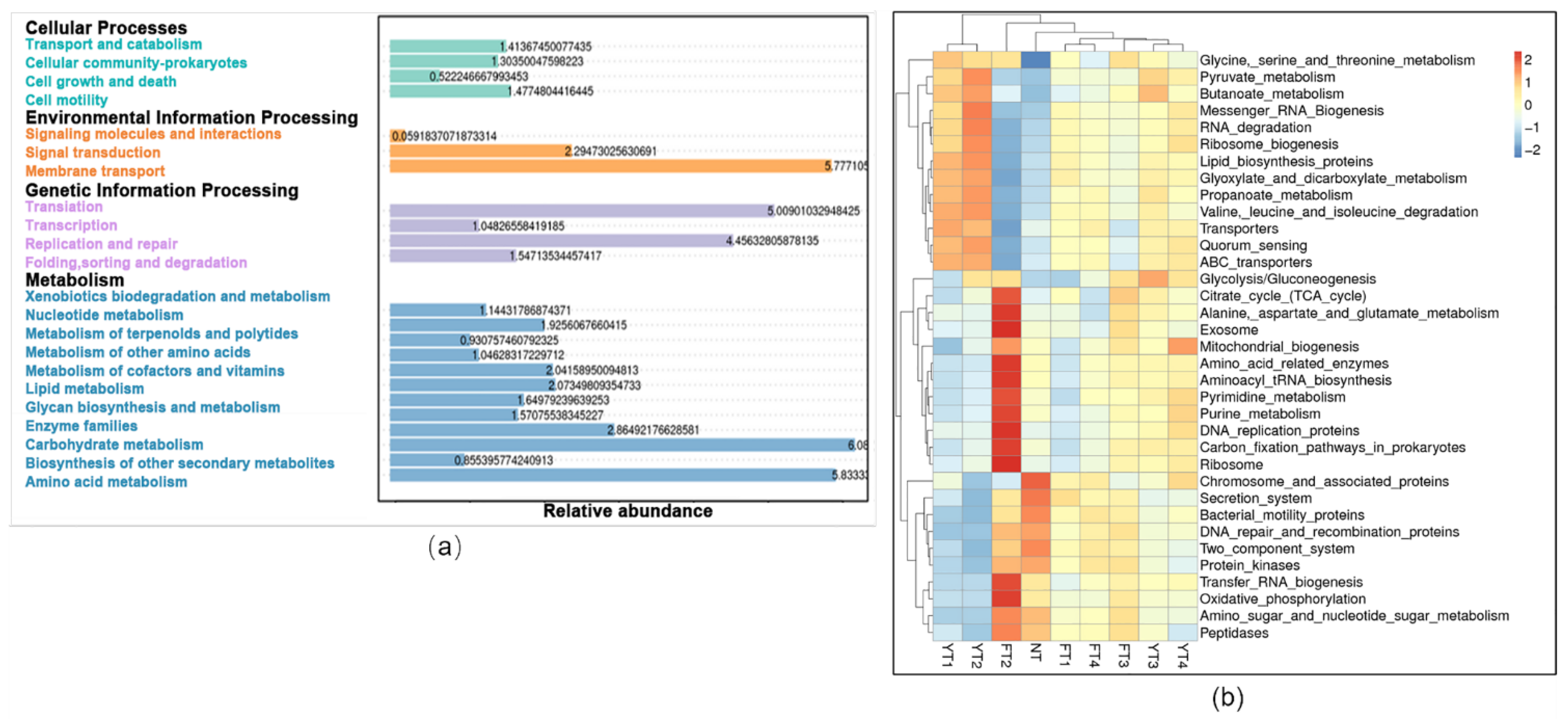
3.3. Bacterial Changes in β-Diversity and Composition Induced Different Functional Responses
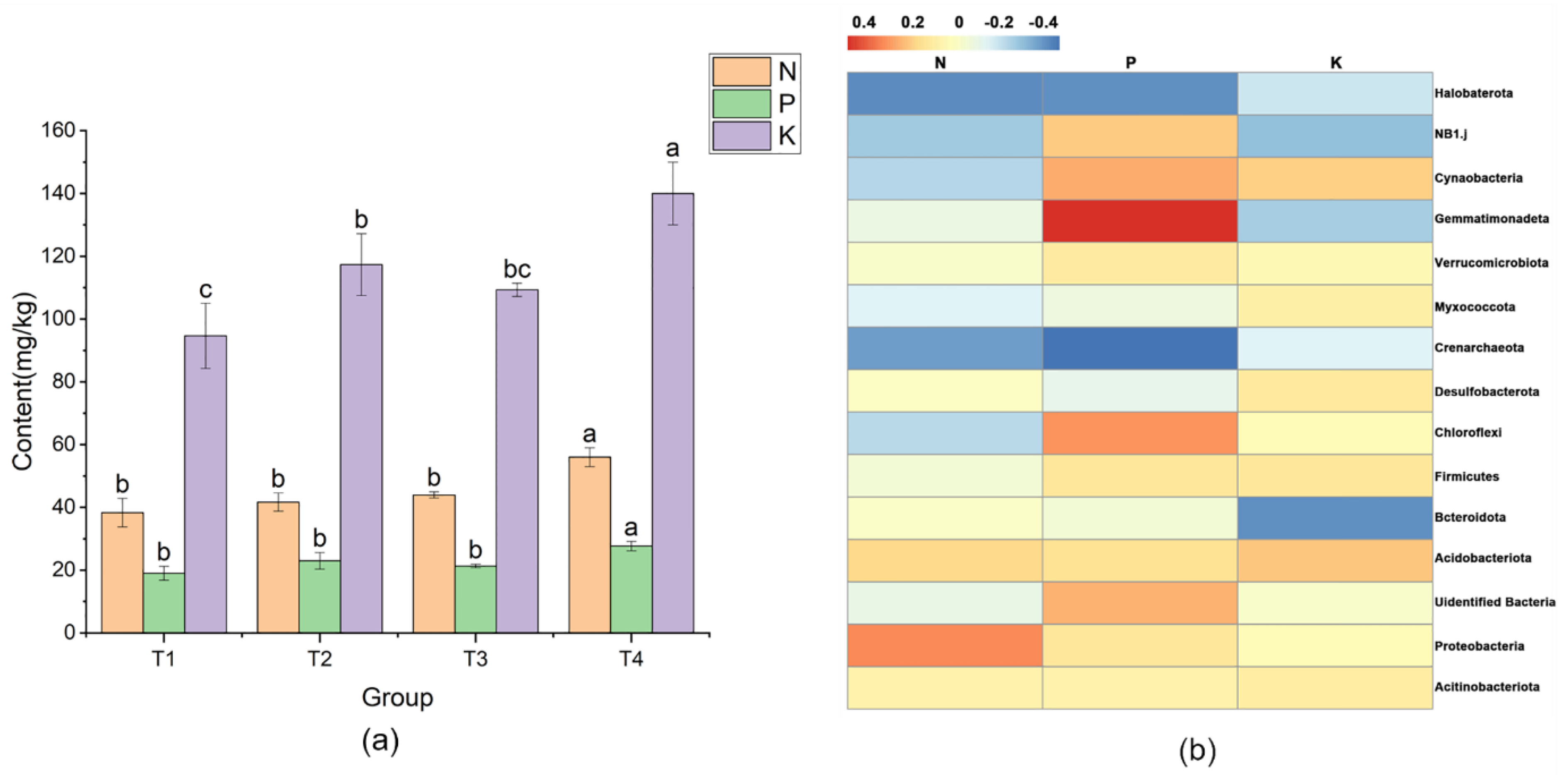
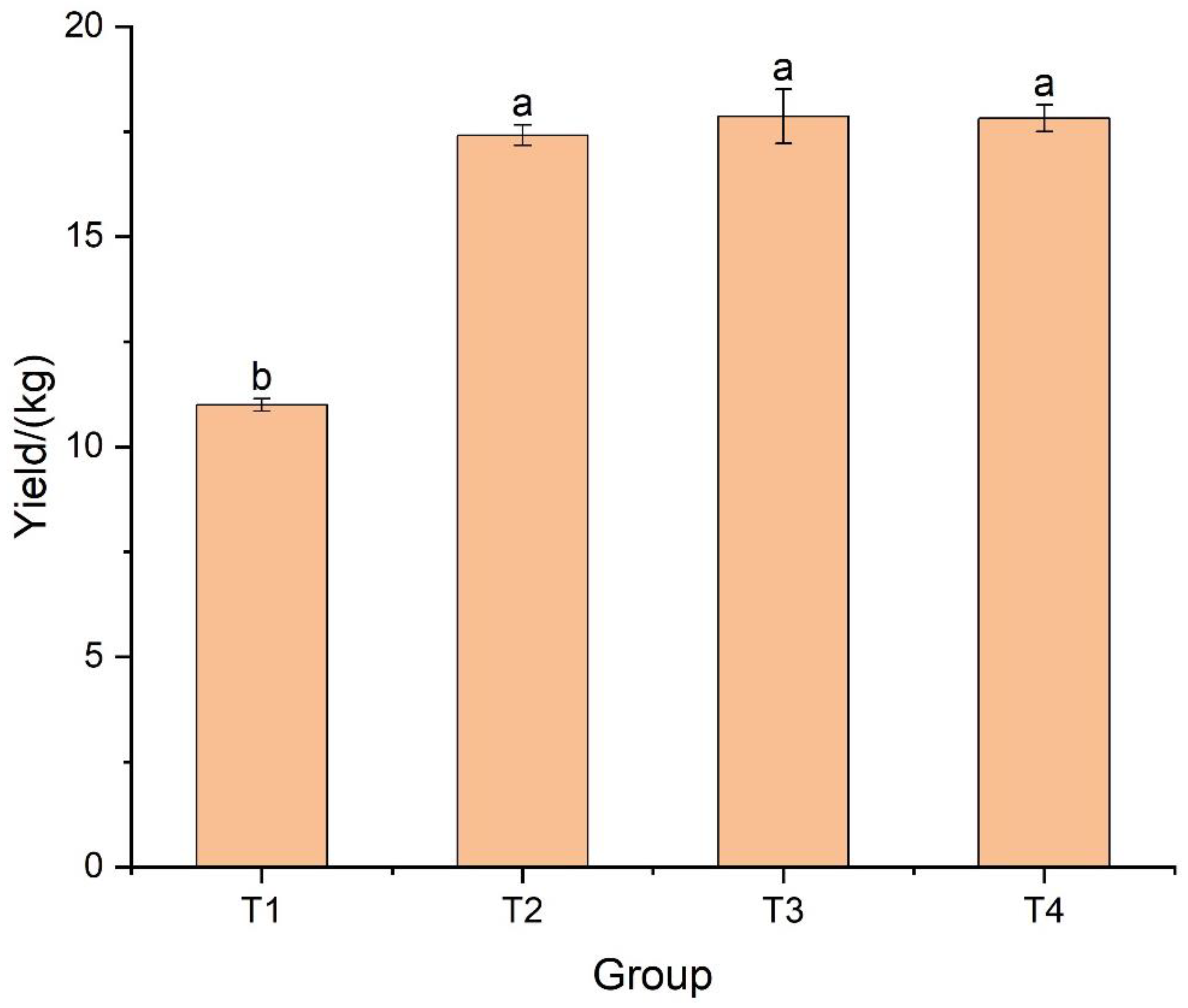
3.4. Improvement in Productivity and Quality of Japonica Rice by Seaweed Extract
4. Discussion
4.1. Rhizosphere Bacteria Dynamics in the Tillering and Heading Stage of Rice
4.2. Effect of Seaweed Extract Supply on Nutrient Absorption in Rice Rhizosphere Soil
4.3. Effects of Seaweed Extract Supply on Rice Yield and Quality
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sasse, J.; Martinoia, E.; Northen, T. Feed your friends: Do plant exudates shape the root microbiome? Trends Plant Sci. 2018, 23, 25–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pascale, A.; Proietti, S.; Pantelides, I.S.; Stringlis, I.A. Modulation of the root microbiome by plant molecules: The basis for targeted disease suppression and plant growth promotion. Front. Plant Sci. 2019, 10, 1741. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Chen, R.; Hu, J.; Zhao, F.; Wang, J.; Chu, H.; Zhang, J.; Dolfing, J.; Lin, X. Bacillus asahii comes to the fore in organic manure fertilized alkaline soils. Soil Biol. Biochem. 2015, 81, 186–194. [Google Scholar] [CrossRef] [Green Version]
- Habibi, S.; Djedidi, S.; Prongjunthuek, K.; Mortuza, M.F.; Ohkama-Ohtsu, N.; Sekimoto, H.; Yokoyoma, T. Physiological and genetic characterization of rice nitrogen fixer PGPR isolated from rhizosphere soils of different crops. Plant. Soil 2014, 379, 51–66. [Google Scholar] [CrossRef]
- Maeder, P.; Fliessbach, A.; Dubois, D.; Gunst, L.; Fried, P.; Niggli, U. Soil fertility and biodiversity in organic farming. Science 2002, 296, 1694–1697. [Google Scholar] [CrossRef] [Green Version]
- Molina-Santiago, C.; Matilla, M.A. Chemical fertilization: A short-term solution for plant productivity? Microb. Biotechnol. 2020, 13, 1311–1313. [Google Scholar] [CrossRef] [Green Version]
- Berg, G.; Smalla, K. Plant species and soil type cooperatively shape the structure and function of microbial communities in the rhizosphere. FEMS Microbiol. Ecol. 2009, 68, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Huang, R.; McGrath, S.P.; Hirsch, P.R.; Clark, I.M.; Storkey, J.; Wu, L.; Zhou, J.; Liang, Y. Plant-microbe networks in soil are weakened by century-long use of inorganic fertilizers. Microb. Biotechnol. 2019, 12, 1464–1475. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.-C.; Shamsi, I.H.; Xu, D.-T.; Wang, G.-H.; Lin, X.-Y.; Jilani, G.; Hussain, N.; Chaudhry, A.N. Chemical fertilizer and organic manure inputs in soil exhibit a vice versa pattern of microbial community structure. Appl. Soil Ecol. 2012, 57, 1–8. [Google Scholar] [CrossRef]
- Wang, B.; Gao, B.; Zimmerman, A.R.; Zheng, Y.; Lyu, H. Novel biochar-impregnated calcium alginate beads with improved water holding and nutrient retention properties. J. Environ. Manag. 2018, 209, 105–111. [Google Scholar] [CrossRef]
- Wei, M.; Hu, G.; Wang, H.; Bai, E.; Lou, Y.; Zhang, A.; Zhuge, Y. 35 years of manure and chemical fertilizer application alters soil microbial community composition in a Fluvo-aquic soil in Northern China. Eur. J. Soil Biol. 2017, 82, 27–34. [Google Scholar] [CrossRef]
- Ning, C.-C.; Gao, P.-D.; Wang, B.-Q.; Lin, W.-P.; Jiang, N.-H.; Cai, K.-Z. Impacts of chemical fertilizer reduction and organic amendments supplementation on soil nutrient, enzyme activity and heavy metal content. J. Integr. Agric. 2017, 16, 1819–1831. [Google Scholar] [CrossRef] [Green Version]
- Hussain, H.I.; Kasinadhuni, N.; Arioli, T. The effect of seaweed extract on tomato plant growth, productivity and soil. J. Appl. Phycol. 2021, 33, 1305–1314. [Google Scholar] [CrossRef]
- Peng, C.; Lai, S.; Luo, X.; Lu, J.; Huang, Q.; Chen, W. Effects of long term rice straw application on the microbial communities of rapeseed rhizosphere in a paddy-upland rotation system. Sci. Total Environ. 2016, 557-558, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Ashour, M.; Hassan, S.M.; Elshobary, M.E.; Ammar, G.A.G.; Gaber, A.; Alsanie, W.F.; Mansour, A.T.; El-Shenody, R. Impact of Commercial Seaweed Liquid Extract (TAM®) Biostimulant and its bioactive molecules on growth and antioxidant activities of hot pepper (Capsicum annuum). Plants 2021, 10, 1045. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Stirk, W.A.; Plackova, L.; Kulkarni, M.G.; Dolezal, K.; Van Staden, J. Interactive effects of plant growth-promoting rhizobacteria and a seaweed extract on the growth and physiology of Allium cepa L. (onion). J. Plant Physiol. 2021, 262, 153437. [Google Scholar] [CrossRef] [PubMed]
- Thiele-Bruhn, S.; Bloem, J.; de Vries, F.T.; Kalbitz, K.; Wagg, C. Linking soil biodiversity and agricultural soil management. Curr. Opin. Environ. Sustain. 2012, 4, 523–528. [Google Scholar] [CrossRef]
- Battacharyya, D.; Babgohari, M.Z.; Rathor, P.; Prithiviraj, B. Seaweed extracts as biostimulants in horticulture. Sci. Hortic-Amestersam 2015, 196, 39–48. [Google Scholar] [CrossRef]
- Colla, G.; Cardarelli, M.; Bonini, P.; Rouphael, Y. Foliar applications of protein hydrolysate, plant and seaweed extracts increase yield but differentially modulate fruit quality of greenhouse tomato. HortScience 2017, 52, 1214–1220. [Google Scholar] [CrossRef]
- De Saeger, J.; Van Praet, S.; Vereecke, D.; Park, J.; Jacques, S.; Han, T.; Depuydt, S. Toward the molecular understanding of the action mechanism of Ascophyllum nodosum extracts on plants. J. Appl. Phycol. 2020, 32, 573–597. [Google Scholar] [CrossRef] [Green Version]
- Khan, W.; Rayirath, U.P.; Subramanian, S.; Jithesh, M.N.; Rayorath, P.; Hodges, D.M.; Critchley, A.T.; Craigie, J.S.; Norrie, J.; Prithiviraj, B. Seaweed extracts as biostimulants of plant growth and development. J. Plant Growth Regul. 2009, 28, 386–399. [Google Scholar] [CrossRef]
- Tejada, M.; Benítez, C.; Gómez, I.; Parrado, J. Use of biostimulants on soil restoration: Effects on soil biochemical properties and microbial community. Appl. Soil Ecol. 2011, 49, 11–17. [Google Scholar] [CrossRef]
- Hassan, S.M.; Ashour, M.; Sakai, N.; Zhang, L.; Hassanien, H.A.; Gaber, A.; Ammar, G. Impact of seaweed liquid extract biostimulant on growth, yield, and chemical composition of cucumber (Cucumis sativus). Agriculture 2021, 11, 320. [Google Scholar] [CrossRef]
- Wang, M.; Chen, L.; Li, Y.; Chen, L.; Liu, Z.; Wang, X.; Yan, P.; Qin, S. Responses of soil microbial communities to a short-term application of seaweed fertilizer revealed by deep amplicon sequencing. Appl. Soil Ecol. 2018, 125, 288–296. [Google Scholar] [CrossRef]
- Mukherjee, A.; Patel, J.S. Seaweed extract: Biostimulator of plant defense and plant productivity. Int. J. Environ. Sci. Technol. 2020, 17, 553–558. [Google Scholar] [CrossRef]
- Al-Taweel, L.S.; Al-Budairy, Z.J. Influence of Vermicompost, Seaweed Extract and Nitrogen Fertilisers on Maize (Zea mays L.) Soil Rhizosphere Microbes. Asian J. Water Environ. 2021, 18, 79–85. [Google Scholar] [CrossRef]
- Tawfeeq, A.; Culham, A.; Davis, F.; Reeves, M. Does fertilizer type and method of application cause significant differences in essential oil yield and composition in rosemary (Rosmarinus officinalis L.)? Ind. Crop. Prod. 2016, 88, 17–22. [Google Scholar] [CrossRef]
- Chen, Y.; Li, J.; Huang, Z.; Su, G.; Li, X.; Sun, Z.; Qin, Y. Impact of short-term application of seaweed fertilizer on bacterial diversity and community structure, soil nitrogen contents, and plant growth in maize rhizosphere soil. Folia Microbiol. 2020, 65, 591–603. [Google Scholar] [CrossRef]
- Arioli, T.; Mattner, S.W.; Hepworth, G.; McClintock, D.; McClinock, R. Effect of seaweed extract application on wine grape yield in Australia. J. Appl. Phycol. 2021, 33, 1883–1891. [Google Scholar] [CrossRef]
- Shen, Y.; Wang, H.; Li, W.; Liu, Z.; Liu, Y.; Wei, H.; Li, J. Synthesis and characterization of double-network hydrogels based on sodium alginate and halloysite for slow release fertilizers. Int. J. Biol. Macromol. 2020, 164, 557–565. [Google Scholar] [CrossRef]
- Edwards, J.; Johnson, C.; Santos-Medellin, C.; Lurie, E.; Podishetty, N.K.; Bhatnagar, S.; Eisen, J.A.; Sundaresan, V. Structure, variation, and assembly of the root-associated microbiomes of rice. Proc. Natl. Acad. Sci. USA 2015, 112, E911–E920. [Google Scholar] [CrossRef] [Green Version]
- Sun, W.; Xiao, E.; Pu, Z.; Krumins, V.; Dong, Y.; Li, B.; Hu, M. Paddy soil microbial communities driven by environment- and microbe-microbe interactions: A case study of elevation-resolved microbial communities in a rice terrace. Sci. Total Environ. 2018, 612, 884–893. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, Y.X.; Zhang, N.; Hu, B.; Jin, T.; Xu, H.; Qin, Y.; Yan, P.; Zhang, X.; Guo, X. NRT1.1B is associated with root microbiota composition and nitrogen use in field-grown rice. Nat. Biotechnol. 2019, 37, 676–684. [Google Scholar] [CrossRef]
- Vafa, Z.N.; Sohrabi, Y.; Sayyed, R.Z.; Luh Suriani, N.; Datta, R. Effects of the combinations of rhizobacteria, mycorrhizae, and seaweed, and supplementary irrigation on growth and yield in wheat cultivars. Plants 2021, 10, 811. [Google Scholar] [CrossRef]
- Hasnain, M.; Chen, J.; Ahmed, N.; Memon, S.; Wang, L.; Wang, Y.; Wang, P. The Effects of fertilizer type and application time on soil properties, plant traits, yield and quality of tomato. Sustainability 2020, 12, 9065. [Google Scholar] [CrossRef]
- Hernández, T.; Chocano, C.; Moreno, J.-L.; García, C. Towards a more sustainable fertilization: Combined use of compost and inorganic fertilization for tomato cultivation. Agric. Ecosyst. Environ. 2014, 196, 178–184. [Google Scholar] [CrossRef]
- Norman, R.J.; Edberg, J.C.; Stucki, J.W. Determination of nitrate in soil extracts by dual-wavelength ultraviolet spectrophotometry. Soil Sci. Soc. Am. J. 1985, 49, 1182–1185. [Google Scholar] [CrossRef]
- Bray, R.H.; Kurtz, L.T. Determination of total, organic and available forms of phosphorus in soils. Soil Sci. 1945, 59, 39–45. [Google Scholar] [CrossRef]
- Carson, P.L. Recommended potassium test. North Dak. Agric. Exp. Stn. Bull. 1980, 17–18. [Google Scholar]
- Wang, M.; Chen, L.; Liu, Z.; Zhang, Z.; Qin, S.; Yan, P. Isolation of a novel alginate lyase-producing Bacillus litoralis strain and its potential to ferment Sargassum horneri for biofertilizer. Microbiologyopen 2016, 5, 1038–1049. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahe, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glockner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Su, J.Q.; Yang, X.R.; Zhu, Y.G. Distinct rhizosphere effect on active and total bacterial communities in paddy soils. Sci. Total Environ. 2019, 649, 422–430. [Google Scholar] [CrossRef]
- Frindte, K.; Zoche, S.A.; Knief, C. Development of a distinct microbial community upon first season crop change in soils of long-term managed maize and rice fields. Front. Microbiol. 2020, 11, 588198. [Google Scholar] [CrossRef]
- Zhou, X.; Wang, J.T.; Zhang, Z.F.; Li, W.; Chen, W.; Cai, L. Microbiota in the rhizosphere and seed of rice from China, With Reference to Their Transmission and Biogeography. Front. Microbiol. 2020, 11, 995. [Google Scholar] [CrossRef]
- Muller, D.B.; Vogel, C.; Bai, Y.; Vorholt, J.A. The plant microbiota: Systems-level insights and perspectives. Annu. Rev. Genet. 2016, 50, 211–234. [Google Scholar] [CrossRef] [Green Version]
- Doi, T.; Abe, J.; Shiotsu, F.; Morita, S. Study on rhizosphere bacterial community in lowland rice grown with organic fertilizers by using PCR-denaturing gradient gel electrophoresis. Plant. Root 2011, 5, 5–16. [Google Scholar] [CrossRef]
- Kristensen, J.M.; Singleton, C.; Clegg, L.A.; Petriglieri, F.; Nielsen, P.H. High diversity and functional potential of undescribed “Acidobacteriota” in Danish wastewater treatment plants. Front. Microbiol. 2021, 12, 643950. [Google Scholar] [CrossRef]
- Wu, T.; Qin, Y.; Li, M. Intercropping of tea (Camellia sinensis L.) and Chinese chestnut: Variation in the structure of rhizosphere bacterial communities. J. Soil Sci. Plant Nutr. 2021, 21, 2178–2190. [Google Scholar] [CrossRef]
- Bloom, A.J. The increasing importance of distinguishing among plant nitrogen sources. Curr. Opin. Plant Biol. 2015, 25, 10–16. [Google Scholar] [CrossRef]
- Mehnaz, S. Microbes—Friends and foes of sugarcane. J. Basic Microbiol. 2013, 53, 954–971. [Google Scholar] [CrossRef]
- Bender, S.F.; Wagg, C.; van der Heijden, M.G.A. An underground revolution: Biodiversity and soil ecological engineering for agricultural sustainability. Trends Ecol. Evol. 2016, 31, 440–452. [Google Scholar] [CrossRef]
- Alam, M.Z.; Braun, G.; Norrie, J.; Mark Hodges, D. Ascophyllum extract application can promote plant growth and root yield in carrot associated with increased root-zone soil microbial activity. Can. J. Plant Sci. 2014, 94, 337–348. [Google Scholar] [CrossRef] [Green Version]
- Natsume, M.; Kamo, Y.; Hirayama, M.; Adachi, T. Isolation and characterization of alginate-derived oligosaccharides with root growth-promoting activities. Carbohydr. Res. 1994, 258, 187–197. [Google Scholar] [CrossRef]
- Nielsen, U.N.; Ayres, E.; Wall, D.H.; Bardgett, R.D. Soil biodiversity and carbon cycling: A review and synthesis of studies examining diversity-function relationships. Eur. J. Soil. Sci. 2011, 62, 105–116. [Google Scholar] [CrossRef]
- Feng, J.; Franks, A.E.; Lu, Z.; Xu, J.; He, Y. Assembly and variation of root-associated microbiota of rice during their vegetative growth phase with and without lindane pollutant. SEL 2020, 3, 207–219. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, N.; Liu, Y.X.; Zhang, X.; Hu, B.; Qin, Y.; Xu, H.; Wang, H.; Guo, X.; Qian, J. Root microbiota shift in rice correlates with resident time in the field and developmental stage. Sci. China Life Sci. 2018, 61, 613–621. [Google Scholar] [CrossRef]
- Jacobs, J.L.; Sundin, G.W. Effect of solar UV-B radiation on a phyllosphere bacterial community. Appl. Environ. Microbiol. 2001, 67, 5488–5496. [Google Scholar] [CrossRef] [Green Version]
- Nabti, E.; Jha, B.; Hartmann, A. Impact of seaweeds on agricultural crop production as biofertilizer. Int. J. Environ. Sci. Technol. 2016, 14, 1119–1134. [Google Scholar] [CrossRef]
- Nedzarek, A. Decomposition of macroalgae and the release of nutrient in Admiralty Bay, King George Island, Antarctica. Polar Biosci. 2004, 17, 26–35. [Google Scholar]
- Yang, S.H.; Seo, J.; Koo, Y. Alginate and fucoidan changes the bacterial community in different directions and the alginate or fucoidan degrading bacteria isolated from paddy soil promotes the plant growth. Arch. Microbiol. 2021, 203, 5183–5192. [Google Scholar] [CrossRef]
- Kumar, D.; Singh, A.P. Efficacy of potassium humate and chemical fertilizers on yield and nutrient availability patterns in soil at different growth stages of rice. Commun. Soil Sci. Plan. 2017, 48, 245–261. [Google Scholar] [CrossRef]
- Yang, L.; Elisa, N.; Eliot, N. Privacy and security aspects of E-Government in smart cities. In Smart Cities Cybersecurity and Privacy; Elsevier: Amsterdam, The Netherlands, 2019; pp. 89–102. [Google Scholar]
- Wang, C.-X.; Zhu, C.-C.; Lu, C.-Y.; Yang, Y.; Li, Q.-F.; Liu, Q.-Q.; Zhang, C.-Q. Grain quality and starch physicochemical properties of chalky rice mutant. Agronomy 2021, 11, 1575. [Google Scholar] [CrossRef]
- Cheng, F.M.; Zhong, L.J.; Wang, F.; Zhang, G.P. Differences in cooking and eating properties between chalky and translucent parts in rice grains. Food Chem. 2005, 90, 39–46. [Google Scholar] [CrossRef]
- Zhu, A.; Zhang, Y.; Zhang, Z.; Wang, B.; Xue, P.; Cao, Y.; Chen, Y.; Li, Z.; Liu, Q.; Cheng, S. Genetic dissection of qPCG1 for a quantitative trait locus for percentage of chalky grain in rice (Oryza sativa L.). Front. Plant. Sci. 2018, 9, 1173. [Google Scholar] [CrossRef]
- Yun-hong, Z.; Li-shu, W.U.; Ming-jian, G.; Hong-qing, H.U.; Shan-xue, Z. Effects of Several Oligosaccharides on the Yield and Quality of Brassica chinensis. J. Huazhong Agric. Univ. 2009, 28, 164–168. [Google Scholar]
- Laporte, D.; Vera, J.; Chandía, N.P.; Zúñiga, E.A.; Matsuhiro, B.; Moenne, A. Structurally unrelated algal oligosaccharides differentially stimulate growth and defense against tobacco mosaic virus in tobacco plants. J. Appl. Phycol. 2006, 19, 79–88. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, W.; Zhao, X.; Wang, H.; Yin, H. Preparation of alginate oligosaccharides and their biological activities in plants: A review. Carbohydr. Res. 2020, 494, 108056. [Google Scholar] [CrossRef]
- Zhang, Y.; Yin, H.; Zhao, X.; Wang, W.; Du, Y.; He, A.; Sun, K. The promoting effects of alginate oligosaccharides on root development in Oryza sativa L. mediated by auxin signaling. Carbohydr. Polym. 2014, 113, 446–454. [Google Scholar] [CrossRef]
- Zhang, Y.H.; Yin, H.; Liu, H.; Wang, W.X.; Wu, L.S.; Zhao, X.M.; Du, Y.G. Alginate oligosaccharides regulate nitrogen metabolism via calcium in Brassica campestris L. var. utilis Tsen et Lee. J. Hortic. Sci. Biotechnol. 2015, 88, 502–508. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | pH | Nitrate Nitrogen | Phosphorus | Potassium |
|---|---|---|---|---|
| (mg/kg) | ||||
| T1 | 8.03 ± 0.06 | 19.15 ± 3.83 | 31.30 ± 3.80 | 195.30 ± 47.06 |
| T2 | 7.96 ± 0.07 | 21.16 ± 6.30 | 25.83 ± 0.31 | 173.56 ± 12.76 |
| T3 | 8.01 ± 0.10 | 18.85 ± 5.86 | 33.23 ± 4.45 | 187.82 ± 15.47 |
| T4 | 8.01 ± 0.04 | 16.64 ± 3.14 | 30.78 ± 7.85 | 202.79 ± 11.43 |
| Group | Base Fertilizer/g | Total Topdressing/g |
|---|---|---|
| T1: negative control | 0 | 0 |
| T2: chemical fertilizer (N-P2O5-K2O:18-20-8) | 720 | 1440 |
| T3: chemical fertilizer with 5‰ S. horneri extract powder | 720 (contained 3.6 g S. horneri extract powder) | 1440 (contained 7.2 g S. horneri extract powder) |
| T4: less chemical fertilizer with 5‰ S. horneri extract powder (80% of that of T3) | 576 (contained 2.88 g S. horneri extract powder) | 1152 (contained 5.76 g S. horneri extract powder) |
| Group | Brown Rice Rate (%) | Milled Rice Rate (%) | Chalkiness Degree (%) | Chalky Rice Rate (%) | Protein Content (%) | Adhesive Strength (mm) | Amylose Content (%) |
|---|---|---|---|---|---|---|---|
| T1 | 84.9 | 76.6 | 1.2 | 8 | 7.12 | 74 | 10.6 |
| T2 | 86.6 | 76.9 | 3.4 | 22 | 7.89 | 79 | 10.3 |
| T3 | 86.1 | 77.4 | 1.8 | 10 | 8.76 | 78 | 10.1 |
| T4 | 85.5 | 75.6 | 2.4 | 13 | 7.54 | 70 | 10.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, C.-L.; Song, W.-L.; Sun, L.; Qin, S.; Ren, C.-G.; Yang, J.-C.; Feng, D.-W.; Liu, N.; Yan, J.; Cui, B.-B.; et al. Effect of Seaweed Extract Supplement on Rice Rhizosphere Bacterial Community in Tillering and Heading Stages. Agronomy 2022, 12, 342. https://doi.org/10.3390/agronomy12020342
Chen C-L, Song W-L, Sun L, Qin S, Ren C-G, Yang J-C, Feng D-W, Liu N, Yan J, Cui B-B, et al. Effect of Seaweed Extract Supplement on Rice Rhizosphere Bacterial Community in Tillering and Heading Stages. Agronomy. 2022; 12(2):342. https://doi.org/10.3390/agronomy12020342
Chicago/Turabian StyleChen, Chun-Lin, Wan-Lin Song, Lin Sun, Song Qin, Cheng-Gang Ren, Jian-Chao Yang, Da-Wei Feng, Ning Liu, Jun Yan, Bi-Bo Cui, and et al. 2022. "Effect of Seaweed Extract Supplement on Rice Rhizosphere Bacterial Community in Tillering and Heading Stages" Agronomy 12, no. 2: 342. https://doi.org/10.3390/agronomy12020342