
An Overview of the Mechanisms Involved in Coffee-Hemileia vastatrix Interactions: Plant and Pathogen Perspectives

 ,
,  , , , , ,
, , , , ,
Abstract
:1. Introduction
2. CLR Causal Agent: Life Cycle and Biotrophic Infection Process
3. Rust Variability
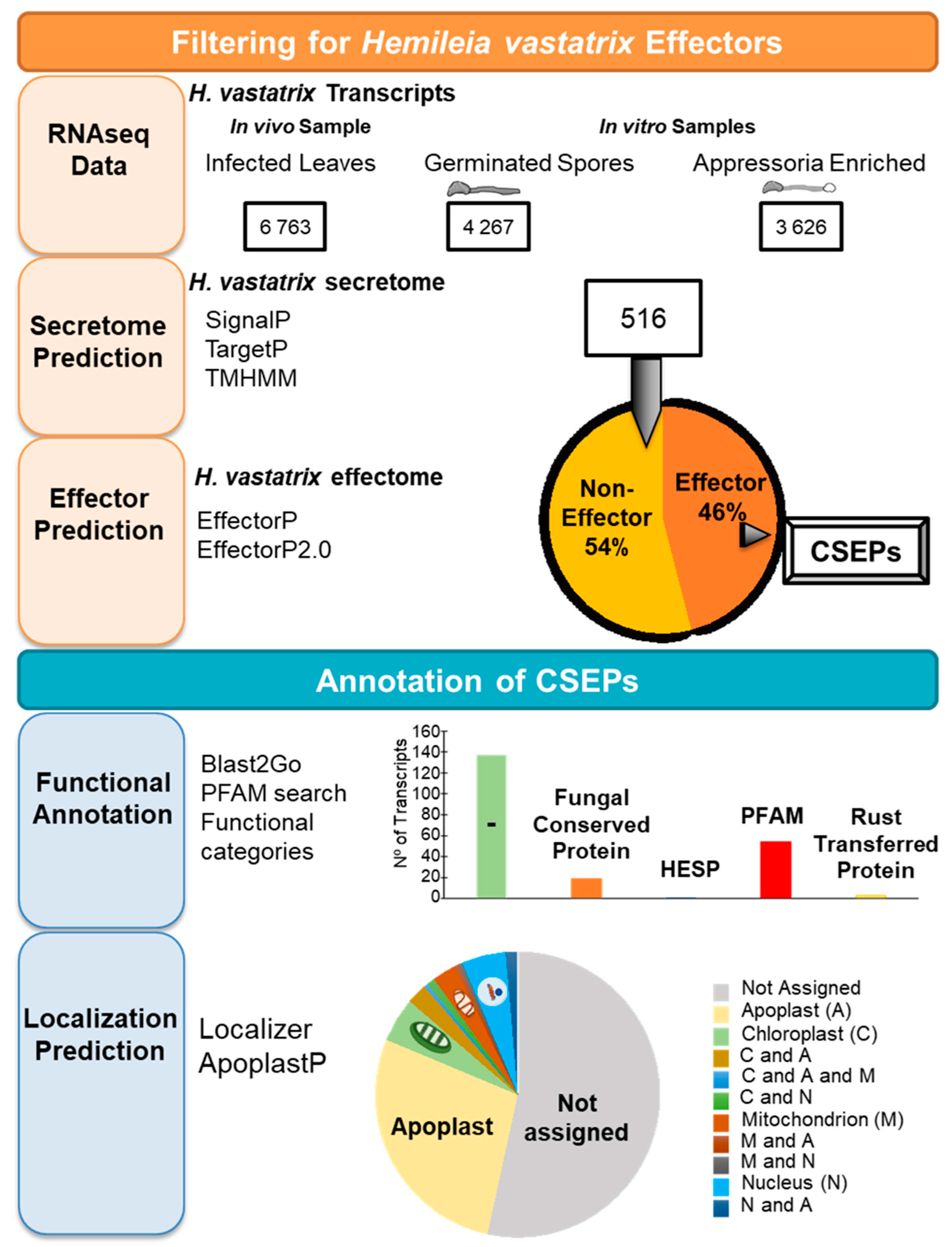
4. Mechanisms of Pathogenicity: The Search for H. vastatrix Effectors
5. Disease Resistance Breeding
5.1. Sources of Resistance to CLR
5.1.1. Spontaneous Interspecific Hybrids
5.1.2. Timor Hybrid (HDT)
5.1.3. HDT Germplasm
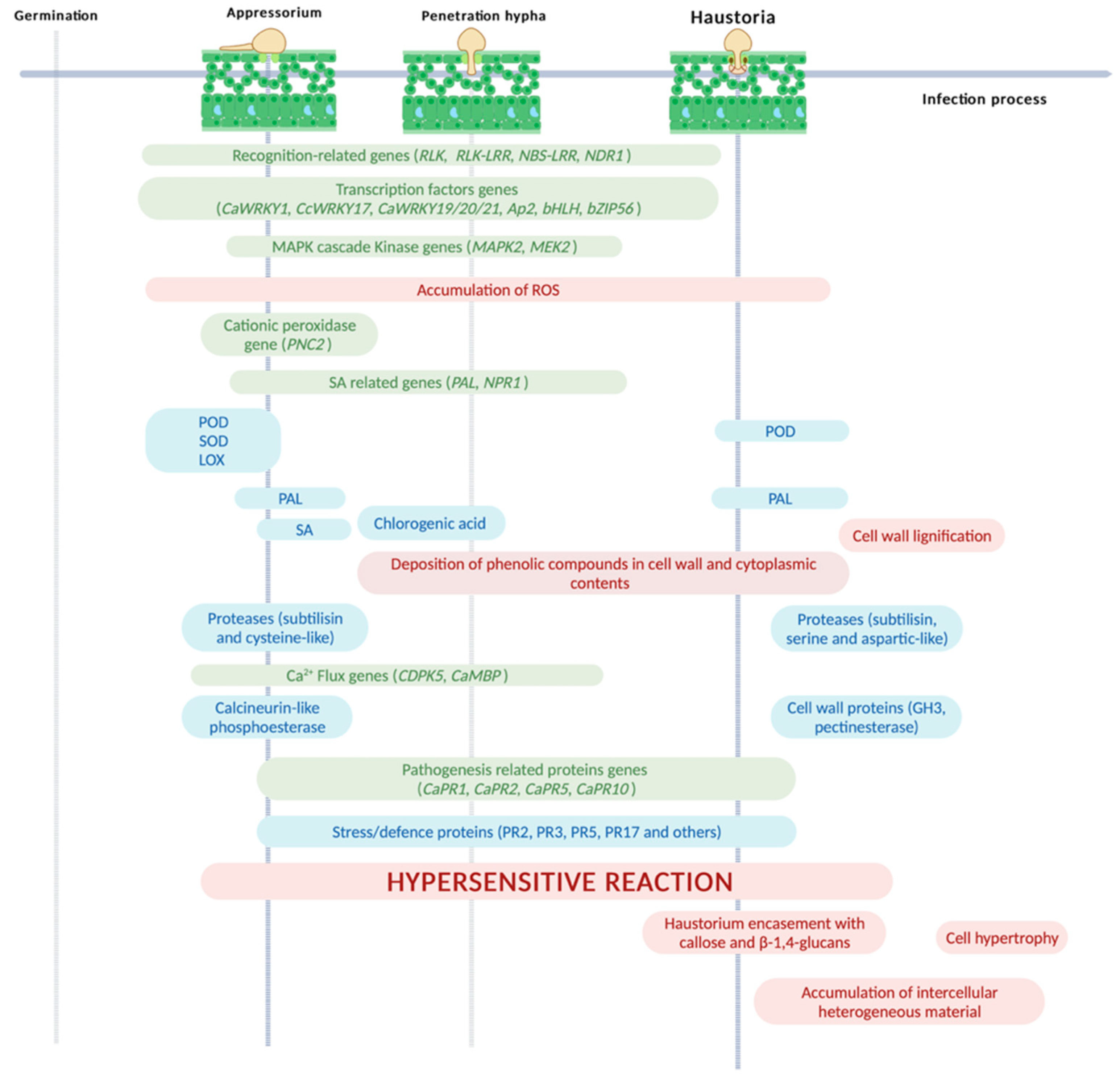
6. Coffee Defense Mechanisms
7. Systemic-Acquired Resistance (SAR) in Coffee
8. Conclusions and Perspectives
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- International Coffee Organization (ICO). Coffee Development Report 2019 Overview; International Coffee Organization: London, UK, 2019. [Google Scholar]
- Davis, A.P.; Tosh, J.; Ruch, N.; Fay, M. Growing coffee: Psilanthus (Rubiaceae) subsumed on the basis of plastid and nuclear DNA sequences; implication for the size, morphology, distribution and evolutionary history of Coffea. Bot. J. Linn. Soc. 2011, 167, 357–377. [Google Scholar] [CrossRef] [Green Version]
- Charrier, A.; Berthaud, J. Botanical classification of coffee. In Coffee: Botany, Biochemistry and Production of Beans and Beverage; Clifford, M.N., Wilson, K.C., Eds.; Croom Helm: London, UK; Sydney, Australia, 1985; ISBN 978-1-4615-6659-5. [Google Scholar]
- International Coffee Organization (ICO). ICO Coffee Production 2020. Available online: https://www.ico.org/prices/po-production.pdf (accessed on 29 December 2021).
- Davis, A.P.; Chadburn, H.; Moat, J.; O’Sullivan, R.; Hargreaves, S.; Lughadha, E.N. High extinction risk for wild coffee species and implications for coffee sector sustainability. Sci. Adv. 2019, 5, eaav3473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Vossen, H.; Bertrand, B.; Charrier, A. Next generation variety development for sustainable production of arabica coffee (Coffea arabica L.): A review. Euphytica 2015, 204, 243–256. [Google Scholar] [CrossRef]
- Talhinhas, P.; Batista, D.; Diniz, I.; Vieira, A.; Silva, D.N.; Loureiro, A.; Tavares, S.; Pereira, A.P.; Azinheira, H.G.; Guerra-Guimarães, L.; et al. Pathogen profile—The coffee leaf rust pathogen Hemileia vastatrix: One and a half centuries around the tropics. Mol. Plant Pathol. 2017, 18, 1039–1051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avelino, J.; Allinne, C.; Cerda, R.; Willocquet, L.; Savary, S. Multiple-disease system in coffee: From crop loss assessment to sustainable management. Annu. Rev. Phytopathol. 2018, 56, 611–635. [Google Scholar] [CrossRef] [PubMed]
- Maghuly, F.; Jankowicz-Cieslak, J.; Bado, S. Improving coffee species for pathogen resistance. CAB Rev. 2020, 15, 1–18. [Google Scholar] [CrossRef]
- Kahn, L.H. Quantitative framework for coffee leaf rust (Hemileia vastatrix), production and futures. Int. J. Agric. Ext. 2019, 7, 77–87. [Google Scholar] [CrossRef]
- Bettencourt, A.J.; Rodrigues, C.J. Principles and practice of coffee breeding for resistance to rust and other diseases. In Coffee Agronomy; Clarke, R.J., Macrae, R., Eds.; Elsevier Applied Science Publishers LTD: London, UK; New York, NY, USA, 1988; Volume 4, pp. 199–234. [Google Scholar]
- McCook, S.; Peterson, P.D. The geopolitics of plant pathology: Frederick Wellman, coffee leaf rust, and cold war networks of science. Annu. Rev. Phytopathol. 2020, 58, 181–199. [Google Scholar] [CrossRef]
- Wellman, F. Peligro de introducción de la Hemileia del café a las Américas. Turrialba 1952, 2, 47–50. [Google Scholar]
- Keith, L.; Sugiyama, L.; Brill, E.; Adams, B.-L.; Fukada, M.; Hoffman, K.; Ocenar, J.; Kawabata, A.; Kong, A.; McKemy, J.; et al. First report of coffee leaf rust caused by Hemileia vastatrix on coffee (Coffea arabica) in Hawaii. Plant Dis. 2021, 34, 2–4. [Google Scholar] [CrossRef]
- Baker, P. The ‘Big Rust’: An update on the coffee leaf rust situation. Coffee Cocoa Int. 2014, 40, 37–39. [Google Scholar]
- Avelino, J.; Cristancho, M.; Georgiou, S.; Imbach, P.; Aguilar, L.; Bornemann, G.; Läderach, P.; Anzueto, F.; Hruska, A.J.; Morales, C. The coffee rust crises in Colombia and Central America (2008–2013): Impacts, plausible causes and proposed solutions. Food Secur. 2015, 7, 303–321. [Google Scholar] [CrossRef] [Green Version]
- McCook, S.; Vandermeer, J. The Big Rust and the Red Queen: Long-term perspectives on coffee rust research. Phytopathology 2015, 105, 1164–1173. [Google Scholar] [CrossRef] [Green Version]
- Rhiney, K.; Guido, Z.; Knudson, C.; Avelino, J.; Bacon, C.M.; Leclerc, G.; Aime, M.C.; Bebber, D.P. Epidemics and the future of coffee production. Proc. Natl. Acad. Sci. USA 2021, 118, e2023212118. [Google Scholar] [CrossRef]
- Noronha-Wagner, M.; Bettencourt, A.J. Genetic study of the resistance of Coffea spp. to leaf rust. Can. J. Bot. 1967, 45, 2021–2031. [Google Scholar] [CrossRef]
- Rodrigues, C.J.; Bettencourt, A.J.; Rijo, L. Races of the pathogen and resistance to coffee rust. Annu. Rev. Phytopathol. 1975, 13, 49–70. [Google Scholar] [CrossRef]
- Várzea, V.M.P.; Marques, D.V. Population variability of Hemileia vastatrix vs. coffee durable resistance. In Durable Resistance to Coffee Leaf Rust; Zambolim, L., Zambolim, E., Várzea, V.M.P., Eds.; Universidade Federal de Viçosa: Viçosa, Brasil, 2005; pp. 53–74. [Google Scholar]
- De Almeida, D.P.; Samila, I.; Castro, L.; Antônio, T.; Mendes, D.O.; Alves, D.R. Receptor-Like Kinase (RLK) as a candidate gene conferring resistance to Hemileia vastatrix in coffee. Sci. Agric. 2021, 78, e20200023. [Google Scholar] [CrossRef]
- Bettgenhaeuser, J.; Gilbert, B.; Ayliffe, M.; Moscou, M.J. Nonhost resistance to rust pathogens—A continuation of continua. Front. Plant Sci. 2014, 5, 664. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Lu, Y.J.; Chen, H.; Day, B. The lifecycle of the plant immune system. Crit. Rev. Plant Sci. 2020, 39, 72–100. [Google Scholar] [CrossRef]
- Nishad, R.; Ahmed, T.; Rahman, V.J.; Kareem, A. Modulation of plant defense system in response to microbial interactions. Front. Microbiol. 2020, 11, 1298. [Google Scholar] [CrossRef]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Upson, J.L.; Zess, E.K.; Białas, A.; Wu, C.H.; Kamoun, S. The coming of age of EvoMPMI: Evolutionary molecular plant–microbe interactions across multiple timescales. Curr. Opin. Plant Biol. 2018, 44, 108–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Wit, P.J.G.M.; Mehrabi, R.; Van Den Burg, H.A.; Stergiopoulos, I. Fungal effector proteins: Past, present and future: Review. Mol. Plant Pathol. 2009, 10, 735–747. [Google Scholar] [CrossRef] [PubMed]
- Zipfel, C. Plant pattern-recognition receptors. Trends Immunol. 2014, 35, 345–351. [Google Scholar] [CrossRef]
- Han, G.Z. Origin and evolution of the plant immune system. New Phytol. 2019, 222, 70–83. [Google Scholar] [CrossRef] [Green Version]
- Flor, H.H. Current status of the gene-for-gene concept. Annu. Rev. Phytopathol. 1971, 9, 275–297. [Google Scholar] [CrossRef]
- Lang, J.; Colcombet, J. Sustained incompatibility between MAPK signaling and pathogen effectors. Int. J. Mol. Sci. 2020, 21, 7954. [Google Scholar] [CrossRef]
- Ravensdale, M.; Bernoux, M.; Ve, T.; Kobe, B.; Thrall, P.H.; Ellis, J.G.; Dodds, P.N. Intramolecular interaction influences binding of the flax L5 and L6 resistance proteins to their AvrL567 ligands. PLoS Pathog. 2012, 8, e1003004. [Google Scholar] [CrossRef]
- Peng, Y.; Van Wersch, R.; Zhang, Y. Convergent and divergent signaling in PAMP-triggered immunity and effector-triggered immunity. Mol. Plant-Microbe Interact. 2018, 31, 403–409. [Google Scholar] [CrossRef] [Green Version]
- Balint-Kurti, P. The plant hypersensitive response: Concepts, control and consequences. Mol. Plant Pathol. 2019, 20, 1163–1178. [Google Scholar] [CrossRef] [Green Version]
- Heath, M.C. Hypersensitive response-related death. Plant Mol. Biol. 2000, 44, 321–334. [Google Scholar] [CrossRef]
- Andersen, E.J.; Ali, S.; Byamukama, E.; Yen, Y.; Nepal, M.P. Disease resistance mechanisms in plants. Genes 2018, 9, 339. [Google Scholar] [CrossRef] [Green Version]
- Periyannan, S.; Milne, R.J.; Figueroa, M.; Lagudah, E.S.; Dodds, P.N. An overview of genetic rust resistance: From broad to specific mechanisms. PLoS Pathog. 2017, 13, e1006380. [Google Scholar] [CrossRef] [Green Version]
- Martins, D.; Araújo, S.D.; Rubiales, D.; Patto, M.C.V. Legume crops and biotrophic pathogen interactions: A continuous cross-talk of a multilayered array of defense mechanisms. Plants 2020, 9, 1460. [Google Scholar] [CrossRef]
- Metraux, J. Systemic acquired resistance and salicylic acid: Current state of knowledge. Eur. J. Plant Pathol. 2001, 107, 13–18. [Google Scholar] [CrossRef]
- Cavalcanti, F.R.; Resende, M.L.V.; Carvalho, C.P.S.; Silveira, J.A.G.; Oliveira, J.T.A. Induced defence responses and protective effects on tomato against Xanthomonas vesicatoria by an aqueous extract from Solanum lycocarpum infected with Crinipellis perniciosa. Biol. Control 2006, 39, 408–417. [Google Scholar] [CrossRef]
- Balmer, A.; Pastor, V.; Gamir, J.; Flors, V.; Mauch-Mani, B. The “prime-ome”: Towards a holistic approach to priming. Trends Plant Sci. 2015, 20, 443–452. [Google Scholar] [CrossRef]
- Gozzo, F.; Faoro, F. Systemic acquired resistance (50 years after discovery): Moving from the lab to the field. J. Agric. Food Chem. 2013, 61, 12473–12491. [Google Scholar] [CrossRef]
- De Resende, M.L.V.; Pozza, E.A.; Reichel, T.; Botelho, D.M.S. Strategies for coffee leaf rust management in organic crop systems. Agronomy 2021, 11, 1865. [Google Scholar] [CrossRef]
- Heath, M.C. Signalling between pathogenic rust fungi and resistant or susceptible host plants. Ann. Bot. 1997, 80, 713–720. [Google Scholar] [CrossRef] [Green Version]
- Voegele, R.T.; Mendgen, K.W. Nutrient uptake in rust fungi: How sweet is parasitic life? Euphytica 2011, 179, 41–55. [Google Scholar] [CrossRef]
- Catanzariti, A.M.; Dodds, P.N.; Lawrence, G.J.; Ayliffe, M.A.; Ellis, J.G. Haustorially expressed secreted proteins from flax rust are highly enriched for avirulence elicitors. Plant Cell 2006, 18, 243–256. [Google Scholar] [CrossRef] [Green Version]
- Garnica, D.P.; Nemri, A.; Upadhyaya, N.M.; Rathjen, J.P.; Dodds, P.N. The ins and outs of rust haustoria. PLoS Pathog. 2014, 10, e1004329. [Google Scholar] [CrossRef] [Green Version]
- Mendgen, K.; Voegele, R. Biology of rusts and mechanisms of infection. In Durable Resistance to Coffee Leave Rust; Zambolim, L., Zambolim, E.M., Várzea, V., Eds.; Universidade Federal de Viçosa: Viçosa, Brasil, 2005; pp. 233–248. [Google Scholar]
- Azinheira, H.G.; Guerra-Guimarães, L.; Silva, M.C.; Várzea, V.; Ricardo, C.P. Esterase activity and adhesion during the early stages of Hemileia vastatrix differentiation. In Proceedings of the 21st International Conference on Coffee Science (ASIC), France, Montpellier, 11–15 September 2006; pp. 1325–1329. [Google Scholar]
- Silva, M.C.; Nicole, M.; Rijo, L.; Geiger, J.P.; Rodrigues, C.J. Cytochemistry of plant- rust fungus interface during the compatible interaction Coffea arabica (cv. Caturra)-Hemileia vastatrix (race III). Int. J. Plant Sci. 1999, 160, 79–91. [Google Scholar] [CrossRef]
- Silva, M.C.; Várzea, V.; Guerra-Guimarães, L.; Azinheira, H.G.; Fernandez, D.; Petitot, A.S.; Bertrand, B.; Lashermes, P.; Nicole, M. Coffee resistance to the main diseases: Leaf rust and coffee berry disease. Braz. J. Plant Physiol. 2006, 18, 119–147. [Google Scholar] [CrossRef] [Green Version]
- Cramer, P.J.S. Review of literature of coffee research in Indonesia. Inter-Am. Inst. Agric. Sci. Misc. Publ. 1952, 15, 262. [Google Scholar]
- Narasimhaswamy, R.L. Coffee leaf disease (Hemileia) in India. Indian Coffee 1961, 25, 382–388. [Google Scholar]
- Mayne, W.W. Physiological specializalion of Hemileia vastatrix. B. & Br. Nature 1932, 129, 510. [Google Scholar] [CrossRef]
- Mayne, W.W. Annual report of the coffee scientific officer 1938–39. Mys. Coffee Exp. Stn. 1939, 19, 1–16. [Google Scholar]
- Zhang, H.; Li, J.; Zhou, H.; Chen, Z.; Pereira, A.P.; Silva, M.C.; Várzea, V. Arabica coffee production in the Yunnan province of China. In Proceedings of the 24th International Conference on Coffee Science (ASIC), San José, Costa Rica, 12–16 November 2012; pp. 679–684. [Google Scholar]
- Noppakoonwong, U.; Khomarwut, C.; Hanthewee, M.; Jarintorn, S.; Hassarungsee, S.; Meesook, S.; Daoruang, C.; Nakai, P.; Lertwatanakiat, S.; Satayawut, K.; et al. Research and Development of Arabica Coffee in Thailand. In Proceedings of the 25th International Conference on Coffee Science (ASIC), Armenia, Colombia, 8–13 September 2014; pp. 42–49. [Google Scholar]
- Gichuru, E.K.; Ithiru, J.M.; Silva, M.C.; Pereira, A.P.; Várzea, V.M.P. Additional physiological races of coffee leaf rust (Hemileia vastatrix) identified in Kenya. Trop. Plant Pathol. 2012, 37, 424–427. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, B.; Bettencourt, A.J.; Rodrigues, C.J.; Rijo, L.; Azinheira, H.; Talhinhas, P.; Silva, M.C.; Várzea, V. Data Records of CIFC’s Research, 1955–2021. Unpublished work.
- Li, L.; Várzea, V.M.P.; Xia, Q.; Xiang, W.; Tang, T.; Zhu, M.; He, C.; Pereira, A.P.; Silva, M.C.; Wu, W.; et al. First report of Hemileia vastatrix (Coffee Leaf Rust) physiological races emergent in coffee germplasm collections in the coffee—Cropping regions of China. Plant Dis. 2021, 105, 4162. [Google Scholar] [CrossRef]
- Tavares, S.; Ramos, A.P.; Pires, A.S.; Azinheira, H.G.; Caldeirinha, P.; Link, T.; Abranches, R.; Silva, M.D.; Voegele, R.T.; Loureiro, J.; et al. Genome size analyses of Pucciniales reveal the largest fungal genomes. Front. Plant Sci. 2014, 5, 422. [Google Scholar] [CrossRef] [Green Version]
- Porto, B.N.; Caixeta, E.T.; Mathioni, S.M.; Vidigal, P.M.P.; Zambolim, L.; Zambolim, E.M.; Donofrio, N.; Polson, S.W.; Maia, T.A.; Chen, C.; et al. Genome sequencing and transcript analysis of Hemileia vastatrix reveal expression dynamics of candidate effectors dependent on host compatibility. PLoS ONE 2019, 14, e0215598. [Google Scholar] [CrossRef]
- Cristancho, M.A.; Botero-Rozo, D.O.; Giraldo, W.; Tabima, J.; Riaño-Pachón, D.M.; Escobar, C.; Rozo, Y.; Rivera, L.F.; Durán, A.; Restrepo, S.; et al. Annotation of a hybrid partial genome of the coffee rust (Hemileia vastatrix) contributes to the gene repertoire catalog of the Pucciniales. Front. Plant Sci. 2014, 5, 594. [Google Scholar] [CrossRef] [Green Version]
- Silva, D.N.; Várzea, V.; Paulo, O.S.; Batista, D. Population genomic footprints of host adaptation, introgression and recombination in coffee leaf rust. Mol. Plant Pathol. 2018, 19, 1742–1753. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, A.S.; Silva, D.N.; Várzea, V.; Paulo, O.S. Scaled-up genomic data uncovers a higher level of population genetic structuring in Hemileia vastatrix. In Proceedings of the 28th International Conference on Coffee Science (ASIC), Montpellier, France, 28 June–1 July 2021; p. 40. [Google Scholar]
- Guerra-Guimarães, L.; Pinheiro, C.; Leclercq, C.; Resende, M.L.V.; Jenny, J.; Ricardo, C.P.; Várzea, V. The first insight on the Hemileia vastatrix urediniospores proteome. In Proceedings of the 28th International Conference on Coffee Science (ASIC), Montpellier, France, 28 June–1 July 2021; p. 223. [Google Scholar]
- Boufleur, T.R.; Massola Júnior, N.S.; Tikami, Í.; Sukno, S.A.; Thon, M.R.; Baroncelli, R. Identification and comparison of Colletotrichum secreted effector candidates reveal two independent lineages pathogenic to soybean. Pathogens 2021, 10, 1520. [Google Scholar] [CrossRef]
- Vleeshouwers, V.G.A.A.; Oliver, R.P. Effectors as tools in disease resistance breeding against biotrophic, hemibiotrophic, and necrotrophic plant pathogens. Mol. Plant-Microbe Interact. 2014, 27, 196–206. [Google Scholar] [CrossRef] [Green Version]
- Gorash, A.; Armonienė, R.; Kazan, K. Can effectoromics and loss-of-susceptibility be exploited for improving Fusarium head blight resistance in wheat? Crop J. 2021, 9, 1–16. [Google Scholar] [CrossRef]
- Fernandez, D.; Tisserant, E.; Talhinhas, P.; Azinheira, H.; Vieira, A.; Petitot, A.S.; Loureiro, A.; Poulain, J.; da Silva, C.; Silva, M.C.; et al. 454-pyrosequencing of Coffea arabica leaves infected by the rust fungus Hemileia vastatrix reveals in planta-expressed pathogen-secreted proteins and plant functions in a late compatible plant-rust interaction. Mol. Plant Pathol. 2012, 13, 17–37. [Google Scholar] [CrossRef]
- Talhinhas, P.; Azinheira, H.G.; Vieira, B.; Loureiro, A.; Tavares, S.; Batista, D.; Morin, E.; Petitot, A.-S.; Paulo, O.S.; Poulain, J.; et al. Overview of the functional virulent genome of the coffee leaf rust pathogen Hemileia vastatrix with an emphasis on early stages of infection. Front. Plant Sci. 2014, 5, 88. [Google Scholar] [CrossRef] [Green Version]
- Sperschneider, J.; Gardiner, D.M.; Dodds, P.N.; Tini, F.; Covarelli, L.; Singh, K.B.; Manners, J.M.; Taylor, J.M. EffectorP: Predicting fungal effector proteins from secretomes using machine learning. New Phytol. 2016, 210, 743–761. [Google Scholar] [CrossRef] [Green Version]
- Sperschneider, J.; Dodds, P.N.; Gardiner, D.M.; Singh, K.B.; Taylor, J.M. Improved prediction of fungal effector proteins from secretomes with EffectorP 2.0. Mol. Plant Pathol. 2018, 19, 2094–2110. [Google Scholar] [CrossRef] [Green Version]
- Loureiro, A.; Tavares, S.; Azinheira, H.G.; Talhinhas, P.; Pereira, A.P.; Fernandez, D.; Silva, M.C. Identification and characterisation of Hemileia vastatrix effectors. In Proceedings of the 25th International Conference on Coffee Science (ASIC), Armenia, Colombia, 8–13 September 2014; p. 250. [Google Scholar]
- Silva, M.C.; Guerra-Guimarães, L.; Azinheira, H.G.; Talhinhas, P.; Tavares, S.; Loureiro, A.; Diniz, I.; Vieira, A.; Pereira, A.P.; Oliveira, H.; et al. Mechanisms involved in coffee—Hemileia vastatrix interactions: Plant and pathogen perspectives. In Proceedings of the 26th International Conference on Coffee Science (ASIC), Kunming, China, 13–19 November 2016; p. 14. [Google Scholar]
- Tavares, S.; Azinheira, H.G.; Voegele, R.; Thordal-Christensen, H.; Silva, M.; Talhinhas, P. Identification of Hemileia vastatrix candidate effectors reveals new ways of promoting pathogen variability through alternative splicing. In Proceedings of the 28th International Conference on Coffee Science (ASIC), Montpellier, France, 28 June–1 July 2021; p. 42. [Google Scholar]
- Maia, T.; Badel, J.L.; Marin-Ramirez, G.; Rocha, C.d.M.; Fernandes, M.B.; da Silva, J.C.F.; de Azevedo-Junior, G.M.; Brommonschenkel, S.H. The Hemileia vastatrix effector HvEC-016 suppresses bacterial blight symptoms in coffee genotypes with the SH1 rust resistance gene. New Phytol. 2017, 213, 1315–1329. [Google Scholar] [CrossRef]
- Sharma, A.; Jones, J.B.; White, F.F. Recent advances in developing disease resistance in plants. F1000Research 2019, 8, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Alvarado, G.; Combes, M.C.; Romero, G.; Lashermes, P. Genetic analysis of partial resistance to coffee leaf rust (Hemileia vastatrix Berk & Br.) introgressed into the cultivated Coffea arabica L. from the diploid C. canephora species. Euphytica 2009, 167, 57–67. [Google Scholar] [CrossRef]
- Romero, G.; Alvarado, G.; Ligarreto, G.; Galeano, N.F. Partial resistance to leaf rust (Hemileia vastatrix) in coffee (Coffea arabica L.): Genetic analysis and molecular characterization of putative candidate genes. Mol. Breed. 2010, 25, 685–697. [Google Scholar] [CrossRef]
- Castillo, Z.P.; Alvarado, A.G. Resistencia incompleta de genotipos de café a la roya bajo condiciones de campo en la región central de Colombia. Cenicafé 1997, 48, 40–58. [Google Scholar]
- Toniutti, L.; Breitler, J.C.; Etienne, H.; Campa, C.; Doulbeau, S.; Urban, L.; Lambot, C.; Pinilla, J.C.H.; Bertrand, B. Influence of environmental conditions and genetic background of Arabica coffee (C. arabica L) on leaf rust (Hemileia vastatrix) pathogenesis. Front. Plant Sci. 2017, 8, 2025. [Google Scholar] [CrossRef] [Green Version]
- Lashermes, P.; Combes, M.C.; Ribas, A.; Cenci, A.; Mahé, L.; Etienne, H. Genetic and physical mapping of the SH3 region that confers resistance to leaf rust in coffee tree (Coffea arabica L.). Tree Genet. Genomes 2010, 6, 973–980. [Google Scholar] [CrossRef]
- Ribas, A.F.; Cenci, A.; Combes, M.C.; Etienne, H.; Lashermes, P. Organization and molecular evolution of a disease-resistance gene cluster in coffee trees. BMC Genom. 2011, 12, 240. [Google Scholar] [CrossRef] [Green Version]
- Prakash, N.S.; Marques, D.V.; Varzea, V.M.P.; Silva, M.C.; Combes, M.C.; Lashermes, P. Introgression molecular analysis of a leaf rust resistance gene from Coffea liberica into C. arabica L. Theor. Appl. Genet. 2004, 109, 1311–1317. [Google Scholar] [CrossRef] [PubMed]
- Mahé, L.; Combes, M.C.; Várzea, V.M.P.; Guilhaumon, C.; Lashermes, P. Development of sequence characterized DNA markers linked to leaf rust (Hemileia vastatrix) resistance in coffee (Coffea arabica L.). Mol. Breed. 2008, 21, 105–113. [Google Scholar] [CrossRef]
- Romero, G.; Vásquez, L.M.; Lashermes, P.; Herrera, J.C. Identification of a major QTL for adult plant resistance to coffee leaf rust (Hemileia vastatrix) in the natural Timor hybrid (Coffea arabica × C. canephora). Plant Breed. 2014, 133, 121–129. [Google Scholar] [CrossRef]
- de Brito, G.G.; Caixeta, E.T.; Gallina, A.P.; Zambolim, E.M.; Zambolim, L.; Diola, V.; Loureiro, M.E. Inheritance of coffee leaf rust resistance and identification of AFLP markers linked to the resistance gene. Euphytica 2010, 173, 255–264. [Google Scholar] [CrossRef] [Green Version]
- Diola, V.; de Brito, G.G.; Caixeta, E.T.; Maciel-Zambolim, E.; Sakiyama, N.S.; Loureiro, M.E. High-density genetic mapping for coffee leaf rust resistance. Tree Genet. Genomes 2011, 7, 1199–1208. [Google Scholar] [CrossRef] [Green Version]
- Pestana, K.N.; Capucho, A.S.; Caixeta, E.T.; de Almeida, D.P.; Zambolim, E.M.; Cruz, C.D.; Zambolim, L.; Pereira, A.A.; de Oliveira, A.C.B.; Sakiyama, N.S. Inheritance study and linkage mapping of resistance loci to Hemileia vastatrix in Híbrido de Timor UFV 443-03. Tree Genet. Genomes 2015, 11, 72. [Google Scholar] [CrossRef] [Green Version]
- Alkimim, E.R.; Caixeta, E.T.; Sousa, T.V.; Pereira, A.A.; de Oliveira, A.C.B.; Zambolim, L.; Sakiyama, N.S. Marker-assisted selection provides arabica coffee with genes from other Coffea species targeting on multiple resistance to rust and coffee berry disease. Mol. Breed. 2017, 37, 6. [Google Scholar] [CrossRef] [Green Version]
- Almeida, D.P.; Caixeta, E.T.; Moreira, K.F.; de Oliveira, A.C.B.; de Freitas, K.N.P.; Pereira, A.A.; Rosado, R.D.S.; Zambolim, L.; Cruz, C.D. Marker-assisted pyramiding of multiple disease resistance genes in coffee genotypes (Coffea arabica). Agronomy 2021, 11, 1763. [Google Scholar] [CrossRef]
- Sousa, T.V.; Caixeta, E.T.; Alkimim, E.R.; Oliveira, A.C.B.; Pereira, A.A.; Sakiyama, N.S.; Zambolim, L.; Resende, M.D.V. Early selection enabled by the implementation of genomic selection in Coffea arabica breeding. Front. Plant Sci. 2019, 9, 1934. [Google Scholar] [CrossRef] [Green Version]
- Cui, L.; Hanika, K.; Visser, R.G.F.; Bai, Y. Improving pathogen resistance by exploiting plant susceptibility genes in coffee (Coffea spp.). Agronomy 2020, 10, 1928. [Google Scholar] [CrossRef]
- Narasimhaswamy, R.L. Arabica selection S. 795. Its origin and performance: A study. Indian Coffee 1960, 24, 197–204. [Google Scholar]
- Srinivasan, K.H.; Narasimhaswamy, R.L. A review of coffee breeding work done at the Government coffee experiment station, Balehonnur. Indian Coffee 1975, 34, 311–321. [Google Scholar]
- Bettencourt, A.J.; Rodrigues, C.J.; Lopes, J. Study of the physiologic specialization of the coffee rust Hemileia vastatrix B.&Br. and selection of coffee clones for the establishment of a standard range of differential hosts for this rust. In Progress Report 1960–1965; Instituto de Investigação Científica Tropical (IICT): Oeiras, Portugal, 1965; pp. 28–46. [Google Scholar]
- Capot, J.L. L’ amélioration du caféir en Côte dÍvoire. Les Lybrides “Arabustas.” Café Cacao Thé 1972, 19, 13–18. [Google Scholar]
- Carvalho, A.; Eskes, A.B.; Fazuoli, L.C. Breeding coffee for rust resistance in Bazil. In Coffee Rust: Epidemiology, Resistance and Management; Kushalappa, A.C., Eskes, A.B., Eds.; CRC Press: Boca Raton, FL, USA, 1989; pp. 295–307. [Google Scholar]
- Cambrony, H. Arabusta and other interspecific fertile hybrids. In Coffee Agronomy; Clarke, R.J., Macrae, R., Eds.; Elsevier Applied Science Publishers LTD: London, UK, 1988; Volume 4, pp. 263–291. [Google Scholar]
- Sreenivasan, M.S. Breeding coffee for leaf rust resistance in India. In Coffee Rust: Epidemiology, Resistance and Management; Kushalappa, A., Eskes, A.B., Eds.; CRC Press: Boca Raton, FL, USA, 1989; pp. 316–323. [Google Scholar]
- Mahé, L.; Várzea, V.; Le Pierres, D.; Combes, M.C.; Lashermes, P. A new source of resistance against coffee leaf rust from New-Caledonian natural interspecific hybrids between Coffea arabica and Coffea canephora. Plant Breed. 2007, 126, 638–641. [Google Scholar] [CrossRef]
- Krug, C.; DePoerck, R. World coffee survey. In FAO Agricultural Studies No76; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 1968. [Google Scholar]
- Bettencourt, A.J. Considerações gerais sobre o ‘Híbrido de Timor’. Circular 1973, 23, 1–20. [Google Scholar]
- Gonçalves, M.M.; Rodrigues, M.L.; Daehnhardt, E. Estudos sobre o café de Timor I. A Hemileia vastatrix B. & Br. no território e o melhoramento da cafeicultura face à doença. Comum. Miss. Est. Agron. Ultramar 1976, 86, 1–29. [Google Scholar]
- Bettencourt, A.J. Melhoramento genético do cafeeiro. Transferência de factores de resistência à H. vastatrix Berk & Br. para as principais cultivares de Coffea arabica L. Ph.D. Thesis, Universidade de Lisboa, Lisboa, Portugal, 1981. [Google Scholar]
- Carvalho, A. Principles and practice of coffee plant breeding for productivity and quality factors: Coffea arabica. In Coffee Agronomy; Clarke, R.J., Macrae, R., Eds.; Elsevier Applied Science Publishers LTD: London, UK, 1988; Volume 4, pp. 129–165. [Google Scholar]
- Sera, G.; Sera, T.; Fonseca, I.C.B.; Ito, D.S. Resistance to leaf rust in coffee cultivars. Coffee Sci. 2010, 5, 59–66. [Google Scholar]
- Carvalho, C.H. Cultivares de Café; Carvalho, C.H., Ed.; Embrapa: Brasília, Brasil, 2007. [Google Scholar]
- Arabica Coffee Varieties. Available online: https://varieties.worldcoffeeresearch.org/varieties (accessed on 20 December 2021).
- Sera, G.H.; Carvalho, C.H.S.; Abrahão, J.C.; Pozza, E.A.; Matiello, J.B.; Almeida, S.R.; Bartelega, L. Coffee leaf rust in Brazil: Historical events, current situation, and control measures. Agronomy 2021. submitted. [Google Scholar]
- Jayarama, D. Chandragiri-farmer friendly new Arabica plant variety. Indian Coffee 2007, 71, 21–25. [Google Scholar]
- Bertrand, B.; Etienne, H.; Cilas, C.; Charrier, A.; Baradat, P. Coffea arabica hybrid performance for yield, fertility and bean weight. Euphytica 2005, 141, 255–262. [Google Scholar] [CrossRef]
- Marie, L.; Abdallah, C.; Campa, C.; Courtel, P.; Bordeaux, M.; Navarini, L.; Lonzarich, V.; Bosselmann, A.S.; Turreira-García, N.; Alpizar, E.; et al. G × E interactions on yield and quality in Coffea arabica: New F1 hybrids outperform American cultivars. Euphytica 2020, 216, 78. [Google Scholar] [CrossRef] [Green Version]
- Alvarado, A.G.; Posada-Suárez, H.E.; Cortina-Guerrero, H.A. Castillo: Nueva variedad de café con resistencia a la roya. Av. Tec. Cenicafé 2005, 337, 1–8. [Google Scholar]
- Castillo, Z.; Moreno, L. La Variedad Colombia: Selección de un Cultivar Compuesto Resistente a la Roya del Cafeto; Cenicafé: Chinchiná, Colombia, 1987; 169p. [Google Scholar]
- Florez, C.; Londoño, C.E.; Guerrero, H.A.; Moncada, M.; Restrepo, E.C.; Ruales, L.N.; Muñoz, C.A.; Rendón, J.R.; Orrego, H. Nueva variedad de porte bajo, altamente productiva, resistente à la roya y al CBD con mayor calidad física del grano. Av. Tec. Cenicafé 2016, 469, 1–8. [Google Scholar]
- Gichimu, B.; Gichuru, E.; Mamati, G.; Nyende, A. Occurrence of Ck-1 gene conferring resistance to Coffee Berry Disease in Coffea arabica cv. Ruiru 11 and its parental genotypes. J. Agric. Crop Res. 2014, 2, 51–61. [Google Scholar]
- Opile, W.; Agwanda, C. Propagation and distribution of cultivar Ruiru 11. Kenya Coffee 1993, 58, 1496–1508. [Google Scholar]
- Pereira, A.A.; Carvalho, G.; Moura, W.M.; Botelho, C.E.; Rezende, J.C.; Oliveira, A.C.B.; Silva, F.L. Cultivares, origens e suas características. In Café Arábica do Plantio à Colheita; Reis, P.R., Cunha, R., Eds.; EPAMIG: Lavras, Brasil, 2010; Volume 1, pp. 163–221. ISBN 978-85-99764-14-5. [Google Scholar]
- Eskes, A.B.; Leroy, T. Coffee Selection and Breeding; Wintgens, J.N., Ed.; Wiley-VCH Verlag GmbH & Co. KGaA.: Weinheim, Germany, 2004. [Google Scholar] [CrossRef]
- Chaves, G.M. Melhoramento do cafeeiro visando a obtenção de cultivares resistentes a Hemileia vastatrix Berk. et Br. Rev. Ceres 1976, 23, 321–332. [Google Scholar]
- Bertrand, B.; Aguillar, G.; Santacreo, R.; Anzuelo, F. El Mejoramiento Genético en América Central. In Desafios de la Cafeicultura Centroamérica; Bertrand, B., Rapidel, B., Eds.; PROMECAFÉ, CIRAD: Paris, France; IRD, CCCR, IICA: San José, Costa Rica, 1999; pp. 407–456. [Google Scholar]
- Bettencourt, A.J. Características agronómicas de selecções derivados de cruzamentos entre Híbrido de Timor e as variedades Caturra, Villa Sarchi e Catuaí. In Proceedings of the Comunicações Simpósio sobre Ferrugens do Cafeeiro; Centro de Investigação das Ferrugens do Cafeeiro: Oeiras, Portugal, 1983; pp. 351–374. [Google Scholar]
- Bettencourt, A.J.; Fazuoli, L. Melhoramento genético de Coffea arabica L.: Transferência de genes de resistência a Hemileia vastatrix do Híbrido de Timor para a cultivar Villa Sarchí de Coffea. Inst. Agron. Camp. 2008, 84, 1–20. [Google Scholar]
- Matiello, J.; Almeida, S.; Ferreira, I.; Carvalho, C.H. Available online: http://sindicafe-mg.com.br/posts/variedades-de-cafe-resistentes-sao-vantajosas (accessed on 16 November 2021).
- van der Vossen, H.A.M.; Walyaro, D.J. Additional evidence for oligogenic inheritance of durable host resistance to coffee berry disease (Colletotrichum kahawae) in arabica coffee (Coffea arabica L.). Euphytica 2009, 165, 105–111. [Google Scholar] [CrossRef]
- Ito, D.; Sera, T.; Sera, G.; Del Grossi, L.; Kanayama, F. Resistance to bacterial blight in arabica coffee cultivars. Crop Breed. Appl. Biot. 2008, 8, 99–103. [Google Scholar] [CrossRef]
- Lashermes, P. Breeding tools for durable resistance to nematodes (Meloidogyne spp.) of coffee varieties. Plant Prot. Sci. 2002, 38, 717–720. [Google Scholar] [CrossRef]
- Petek, M.; Sera, T.; Sera, G.; Fonseca, I.C.; Ito, D. Selection of progenies of Coffea arabica with simultaneous resistance to bacterial blight and leaf rust. Bragantia 2006, 65, 65–73. [Google Scholar] [CrossRef]
- Rezende, R.; Salgado, S.M.; Rezende, J.; Carvalho, G.; Pereira, A.; Lima, R.; Ferreira, A. Resistência de progênies de Coffea arabica em área infestada por Meloidogyne exigua. Nematropica 2013, 43, 233–240. [Google Scholar]
- Bertrand, B.; Anthony, F.; Lashermes, P. Breeding for resistance to Meloidogyne exigua in Coffea arabica by introgression of resistance genes of Coffea canephora. Plant Pathol. 2001, 50, 637–643. [Google Scholar] [CrossRef]
- Melo, G.A.; Shimizu, M.M.; Mazzafera, P. Polyphenoloxidase activity in coffee leaves and its role in resistance against the coffee leaf miner and coffee leaf rust. Phytochemistry 2006, 67, 277–285. [Google Scholar] [CrossRef]
- Reichel, T.; de Resende, M.L.V.; Monteiro, A.C.A.; Freitas, N.C.; dos Santos Botelho, D.M. Constitutive defense strategy of coffee under field conditions: A comparative assessment of resistant and susceptible cultivars to rust. Mol. Biotechnol. 2021. [Google Scholar] [CrossRef]
- Silva, M.C.; Guerra-Guimarães, L.; Loureiro, A.; Nicole, M.R. Involvement of peroxidases in the coffee resistance to orange rust (Hemileia vastatrix). Physiol. Mol. Plant Pathol. 2008, 72, 29–38. [Google Scholar] [CrossRef]
- Silva, M.C.; Nicole, M.; Guerra-Guimarães, L.; Rodrigues, C.J. Hypersensitive cell death and post-haustorial defence responses arrest the orange rust (Hemileia vastatrix) growth in resistant coffee leaves. Physiol. Mol. Plant Pathol. 2002, 60, 169–183. [Google Scholar] [CrossRef]
- Ganesh, D.; Petitot, A.S.; Silva, M.C.; Alary, R.; Lecouls, A.C.; Fernandez, D. Monitoring of the early molecular resistance responses of coffee (Coffea arabica L.) to the rust fungus (Hemileia vastatrix) using real-time quantitative RT-PCR. Plant Sci. 2006, 170, 1045–1051. [Google Scholar] [CrossRef]
- Guerra-Guimarães, L.; Cardoso, S.; Martins, I.; Loureiro, A.; Bernardes, S.A.; Várzea, V.; Silva, M.C. Differential induction of superoxide dismutase in Coffea arabica—Hemileia vastatrix interactions. In Proceedings of the 22th International Conference on Coffee Science (ASIC), Campinas, Brazil, 14–19 September 2008; pp. 1036–1039. [Google Scholar]
- Diniz, I.; Talhinhas, P.; Azinheira, H.G.; Várzea, V.; Medeira, C.; Maia, I.; Petitot, A.-S.; Nicole, M.; Fernandez, D.; Silva, M.C. Cellular and molecular analyses of coffee resistance to Hemileia vastatrix and nonhost resistance to Uromyces vignae in the resistance-donor genotype HDT832/2. Eur. J. Plant Pathol. 2012, 133, 141–157. [Google Scholar] [CrossRef]
- Guerra-Guimarães, L.; Tenente, R.; Pinheiro, C.; Chaves, I.; Silva, M.D.; Cardoso, F.M.H.; Planchon, S.; Barros, D.R.; Renaut, J.; Ricardo, C.P. Proteomic analysis of apoplastic fluid of Coffea arabica leaves highlights novel biomarkers for resistance against Hemileia vastatrix. Front. Plant Sci. 2015, 6, 478. [Google Scholar] [CrossRef] [Green Version]
- Diniz, I.; Figueiredo, A.; Sebastiana, M.; Munoz-Pajarez, A.J.; Valverde, J.; Azevedo, H.; Rodrigues, A.S.; Prakash, R.S.; Pereira, A.P.; Guerra-Guimarães, L.; et al. First steps on the resistance profiling of Kawisari coffee hybrid through cytological and gene expression analyses. In Proceedings of the 28th International Conference on Coffee Science (ASIC), Montpellier, France, 28 June–1 July 2021; p. 60. [Google Scholar]
- Ramiro, D.; Escoute, J.; Petitot, A.S.; Nicole, M.; Maluf, M.P.; Fernandez, D. Biphasic haustorial differentiation of coffee rust (Hemileia vastatrix race II) associated with defence responses in resistant and susceptible coffee cultivars. Plant Pathol. 2009, 58, 944–955. [Google Scholar] [CrossRef]
- Silva, M.C.; Loureiro, A.; Guerra-Guimarães, L.; Nicole, M.; Valente, P.; Rodrigues, C.J., Jr. Active oxygen metabolism in the hypersensitive response of coffee-rust interaction. In Proceedings of the 11th Congress of the Mediterranean Phytopathological Union and 3rd Congress of the Sociedade Portuguesa de Fitopatologia, Évora, Portugal, 17–20 September 2001; pp. 346–348. [Google Scholar]
- Sá, M.; Ferreira, J.P.; Queiroz, V.T.; Vilas-Boas, L.; Silva, M.C.; Almeida, M.H.; Guerra-Guimarães, L.; Bronze, M.R. A liquid chromatography/electrospray ionisation tandem mass spectrometry method for the simultaneous quantification of salicylic, jasmonic and abscisic acids in Coffea arabica leaves. J. Sci. Food Agric. 2014, 94, 529–536. [Google Scholar] [CrossRef]
- Meyer, D.; Pajonk, S.; Micali, C.; O’Connell, R.; Schulze-Lefert, P. Extracellular transport and integration of plant secretory proteins into pathogen-induced cell wall compartments. Plant J. 2009, 57, 986–999. [Google Scholar] [CrossRef]
- Guerra-Guimarães, L.; Silva, M.C.; Struck, C.; Loureiro, A.; Nicole, M.; Rodrigues, C.J.; Ricardo, C.P.P. Chitinases of Coffea arabica genotypes resistant to orange rust Hemileia vastatrix. Biol. Plant. 2009, 53, 702–706. [Google Scholar] [CrossRef]
- Maxemiuc-Naccache, V.; Braga, M.R.; Dietrich, S.M.C. Chitinase and beta-1,3-glucanase changes in compatible and incompatible combinations between coffee leaf disks and coffee rust (Hemileia vastatrix). Rev. Bras. Bot. 1992, 15, 145–150. [Google Scholar]
- Slusarenko, A.J. The role of lipoxygenase in plant resistance to infection. In Lipoxygenase and Lipoxygenase Pathway Enzymes; AOCS Press: Champaign, IL, USA, 1996; pp. 176–197. [Google Scholar]
- Fernandez, D.; Santos, P.; Agostini, C.; Bon, M.C.; Petitot, A.S.; Silva, M.C.; Guerra-Guimarães, L.; Ribeiro, A.; Argout, X.; Nicole, M. Coffee (Coffea arabica L.) genes early expressed during infection by the rust fungus (Hemileia vastatrix). Mol. Plant Pathol. 2004, 5, 527–536. [Google Scholar] [CrossRef] [Green Version]
- Florez, J.C.; Mofatto, L.S.; do Livramento Freitas-Lopes, R.; Ferreira, S.S.; Zambolim, E.M.; Carazzolle, M.F.; Zambolim, L.; Caixeta, E.T. High throughput transcriptome analysis of coffee reveals prehaustorial resistance in response to Hemileia vastatrix infection. Plant Mol. Biol. 2017, 95, 607–623. [Google Scholar] [CrossRef] [Green Version]
- Diola, V.; Brito, G.G.; Caixeta, E.T.; Pereira, L.F.P.; Loureiro, M.E. A new set of differentially expressed signaling genes is early expressed in coffee leaf rust race II incompatible interaction. Funct. Integr. Genom. 2013, 13, 379–389. [Google Scholar] [CrossRef]
- Guzzo, S.D.; Harakava, R.; Tsai, S.M. Identification of coffee genes expressed during systemic acquired resistance and incompatible interaction with Hemileia vastatrix. J. Phytopathol. 2009, 157, 625–638. [Google Scholar] [CrossRef]
- Couttolenc-Brenis, E.; Carrión, G.L.; Villain, L.; Ortega-Escalona, F.; Ramírez-Martínez, D.; Mata-Rosas, M.; Méndez-Bravo, A. Prehaustorial local resistance to coffee leaf rust in a Mexican cultivar involves expression of salicylic acid-responsive genes. PeerJ 2020, 2020, e8345. [Google Scholar] [CrossRef] [Green Version]
- Ramiro, D.; Jalloul, A.; Petitot, A.S.; Grossi de Sá, M.F.; Maluf, M.P.; Fernandez, D. Identification of coffee WRKY transcription factor genes and expression profiling in resistance responses to pathogens. Tree Genet. Genomes 2010, 6, 767–781. [Google Scholar] [CrossRef]
- Petitot, A.S.; Lecouls, A.C.; Fernandez, D. Sub-genomic origin and regulation patterns of a duplicated WRKY gene in the allotetraploid species Coffea arabica. Tree Genet. Genomes 2008, 4, 379–390. [Google Scholar] [CrossRef]
- Petitot, A.S.; Barsalobres-Cavallari, C.; Ramiro, D.; Albuquerque Freire, E.; Etienne, H.; Fernandez, D. Promoter analysis of the WRKY transcription factors CaWRKY1a and CaWRKY1b homoeologous genes in coffee (Coffea arabica). Plant Cell Rep. 2013, 32, 1263–1276. [Google Scholar] [CrossRef]
- Niks, R.E.; Rubiales, D. Potentially durable resistance mechanisms in plants to specialised fungal pathogens. Euphytica 2002, 124, 201–216. [Google Scholar] [CrossRef]
- Vaz-Patto, M.C.; Rubiales, D. Unveiling common responses of Medicago truncatula to appropriate and inappropriate rust species. Front. Plant Sci. 2014, 5, 618. [Google Scholar] [CrossRef] [Green Version]
- Barka, G.D.; Caixeta, E.T.; de Almeida, R.F.; Alvarenga, S.M.; Zambolim, L. Differential expression of molecular rust resistance components have distinctive profiles in Coffea arabica—Hemileia vastatrix interactions. Eur. J. Plant Pathol. 2017, 149, 543–561. [Google Scholar] [CrossRef]
- Santos, D. Comparative Analysis of 454 Pyrosequencing Data from Coffee Transcriptomes. Master’s Thesis, Universidade de Lisboa, Lisboa, Portugal, 2011. [Google Scholar]
- Tang, D.; Wang, G.; Zhou, J.M. Receptor kinases in plant-pathogen interactions: More than pattern recognition. Plant Cell 2017, 29, 618–637. [Google Scholar] [CrossRef] [Green Version]
- Knepper, C.; Savory, E.A.; Day, B. The role of NDR1 in pathogen perception and plant defense signaling. Plant Signal. Behav. 2011, 6, 1114–1116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cacas, J.L.; Petitot, A.S.; Bernier, L.; Estevan, J.; Conejero, G.; Mongrand, S.; Fernandez, D. Identification and characterization of the Non-race specific Disease Resistance 1 (NDR1) orthologous protein in coffee. BMC Plant Biol. 2011, 11, 144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNeece, B.T.; Pant, S.R.; Sharma, K.; Niruala, P.; Lawrence, G.W.; Klink, V.P. A Glycine max homolog of Non-race specific Disease Resistance 1 (NDR1) alters defense gene expression while functioning during a resistance response to different root pathogens in different genetic backgrounds. Plant Physiol. Biochem. 2017, 114, 60–71. [Google Scholar] [CrossRef] [PubMed]
- Bouché, N.; Yellin, A.; Snedden, W.A.; Fromm, H. Plant-specific calmodulin-binding proteins. Annu. Rev. Plant Biol. 2005, 56, 435–466. [Google Scholar] [CrossRef]
- Aldon, D.; Mbengue, M.; Mazars, C.; Galaud, J.P. Calcium signalling in plant biotic interactions. Int. J. Mol. Sci. 2018, 19, 665. [Google Scholar] [CrossRef] [Green Version]
- Jagodzik, P.; Tajdel-Zielinska, M.; Ciesla, A.; Marczak, M.; Ludwikow, A. Mitogen-activated protein kinase cascades in plant hormone signaling. Front. Plant Sci. 2018, 9, 1387. [Google Scholar] [CrossRef]
- Krysan, P.J.; Colcombet, J. Cellular complexity in MAPK signaling in plants: Questions and emerging tools to answer them. Front. Plant Sci. 2018, 871, 1674. [Google Scholar] [CrossRef]
- Phukan, U.J.; Jeena, G.S.; Shukla, R.K. WRKY transcription factors: Molecular regulation and stress responses in plants. Front. Plant Sci. 2016, 7, 760. [Google Scholar] [CrossRef] [Green Version]
- Wani, S.H.; Anand, S.; Singh, B.; Bohra, A.; Joshi, R. WRKY transcription factors and plant defense responses: Latest discoveries and future prospects. Plant Cell Rep. 2021, 40, 1071–1085. [Google Scholar] [CrossRef]
- Bari, R.; Jones, J.D.G. Role of plant hormones in plant defence responses. Plant Mol. Biol. 2009, 69, 473–488. [Google Scholar] [CrossRef]
- Barsalobre-Cavallari, C.F.; Petitot, A.S.; Severino, F.E.; Maia, I.G.; Fernandez, D. Host response profiling to fungal infection: Molecular cloning, characterization and expression analysis of NPR1 gene from coffee (Coffea arabica). In Microbial pathogens and strategies for combating them: Science, technology and education; Mendez-Vilas, A., Ed.; Formatex Research Center: Badajoz, Spain, 2013; pp. 411–418. [Google Scholar]
- Tang, B.; Li, X.; Pu, L.; Zhao, Q.; Cui, X.; Ge, F.; Liu, D. A pathogenesis-related protein 10 gene PnPR10–3 was involved in molecular interaction between Panax notoginseng and Fusarium solani. Australas. Plant Pathol. 2019, 48, 447–456. [Google Scholar] [CrossRef]
- Van Loon, L.C.; Rep, M.; Pieterse, C.M.J. Significance of inducible defense-related proteins in infected plants. Annu. Rev. Phytopathol. 2006, 44, 135–162. [Google Scholar] [CrossRef] [Green Version]
- Breen, S.; Williams, S.J.; Outram, M.; Kobe, B.; Solomon, P.S. Emerging Insights into the Functions of Pathogenesis-Related Protein 1. Trends Plant Sci. 2017, 22, 871–879. [Google Scholar] [CrossRef]
- Daudi, A.; Cheng, Z.; O’Brien, J.A.; Mammarella, N.; Khan, S.; Ausubel, F.M.; Paul Bolwell, G. The apoplastic oxidative burst peroxidase in Arabidopsis is a major component of pattern-triggered immunity. Plant Cell 2012, 24, 275–287. [Google Scholar] [CrossRef] [Green Version]
- Jwa, N.S.; Hwang, B.K. Convergent evolution of pathogen effectors toward reactive oxygen species signaling networks in plants. Front. Plant Sci. 2017, 8, 1687. [Google Scholar] [CrossRef] [Green Version]
- Mellersh, D.G.; Foulds, I.V.; Higgins, V.J.; Heath, M.C. H2O2 plays different roles in determining penetration failure in three diverse plant-fungal interactions. Plant J. 2002, 29, 257–268. [Google Scholar] [CrossRef]
- Torres, M.A.; Jones, J.D.G.; Dangl, J.L. Reactive oxygen species signaling in response to pathogens. Plant Physiol. 2006, 141, 373–378. [Google Scholar] [CrossRef] [Green Version]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef] [Green Version]
- Hwang, I.S.; Hwang, B.K. The pepper 9-lipoxygenase gene CaLOX1 functions in defense and cell death responses to microbial pathogens. Plant Physiol. 2010, 152, 948–967. [Google Scholar] [CrossRef] [Green Version]
- Rojas, M.L.; Montes de Gómez, V.; Ocampo, C.A. Stimulation of lipoxygenase activity in colyledonary leaves of coffee reacting hypersensitively to the coffee leaf rust. Physiol. Mol. Plant Pathol. 1993, 43, 209–219. [Google Scholar] [CrossRef]
- Göbel, C.; Feussner, I.; Rosahl, S. Lipid Peroxidation during the hypersensitive response in potato in the absence of 9-Lipoxygenases. J. Biol. Chem. 2003, 278, 52834–52840. [Google Scholar] [CrossRef] [Green Version]
- Vermerris, W.; Nicholson, R. Phenolic Compound Biochemistry; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2006; ISBN 9781402051630. [Google Scholar]
- Bagal, U.R.; Leebens-Mack, J.H.; Lorenz, W.W.; Dean, J.F.D. The phenylalanine ammonia lyase (PAL) gene family shows a gymnosperm-specific lineage. BMC Genom. 2012, 13, S1. [Google Scholar] [CrossRef]
- Cesarino, I. Structural features and regulation of lignin deposited upon biotic and abiotic stresses. Curr. Opin. Biotechnol. 2019, 56, 209–214. [Google Scholar] [CrossRef]
- Campa, C.; Doulbeau, S.; Dussert, S.; Hamon, S.; Noirot, M. Qualitative relationship between caffeine and chlorogenic acid contents among wild Coffea species. Food Chem. 2005, 93, 135–139. [Google Scholar] [CrossRef]
- Leitão, S.; Guerra-Guimarães, L.; Bronze, M.R.; Vilas Boas, L.; Sá, M.; Almeida, M.H.G.; Silva, M.C. Chlorogenic acid content in coffee leaves: Possible role in coffee leaf rust resistance. In Proceedings of the 23rd International Conference on Coffee Science (ASIC), Bali, Indonesia, 3–8 October 2010; pp. 743–747. [Google Scholar]
- Caillaud, M.C.; Wirthmueller, L.; Sklenar, J.; Findlay, K.; Piquerez, S.J.M.; Jones, A.M.E.; Robatzek, S.; Jones, J.D.G.; Faulkner, C. The plasmodesmal protein PDLP1 localises to haustoria-associated membranes during downy mildew infection and regulates callose deposition. PLoS Pathog. 2014, 10, e1004496. [Google Scholar] [CrossRef] [PubMed]
- Conrath, U. Chapter 9 Priming of Induced Plant Defense Responses, 1st ed.; Elsevier Ltd.: Amsterdam, The Netherlands, 2009; Volume 51. [Google Scholar] [CrossRef]
- Conrath, U.; Beckers, G.J.M.; Langenbach, C.J.G.; Jaskiewicz, M.R. Priming for Enhanced Defense. Annu. Rev. Phytopathol. 2015, 53, 97–119. [Google Scholar] [CrossRef] [PubMed]
- Martins, E.M.F.; Moraes, W.B.C. Development of Hemileia vastatrix in Coffee Plants with Genetic or Induced Resistance. J. Phytopathol. 1996, 144, 519–526. [Google Scholar] [CrossRef]
- Guzzo, S.D.; Martins, E.M.F. Local and systemic induction of β-1,3-glucanase and chitinase in coffee leaves protected against Hemileia vastatrix by Bacillus thuringiensis. J. Phytopathol. 1996, 144, 449–454. [Google Scholar] [CrossRef]
- Roveratni, D.S.; Teixeira, A.R.R.; Moraes, W.B.C. Bacillus thuringiensis—A new perspective for an Induced Protection to Coffee Leaf Rust. J. Phytopathol. 1989, 126, 149–159. [Google Scholar] [CrossRef]
- Guzzo, S.D.; Bach, E.E.; Martins, E.F.; Moraes, W.C. Crude Exopolysaccharides (EPS) from Xanthomonas campestris pv. manihotis, Xanthomonas campestris pv. Campestris and Commercial Xanthan Gum as Inducers of Protection in Coffee Plants against Hemileia vastatrix. J. Phytopathol. 1993, 139, 119–128. [Google Scholar] [CrossRef]
- Henrique, L.; Fernandes, M.; Lúcio, M.; Resende, V.; Pereira, R.; Costa, B.; Monteiro, A.; Júnior, P. Acidenzolar-S-methyl in rust and blotch control in field conditions. Coffee Sci. 2013, 8, 21–29. [Google Scholar]
- Carré-Missio, V.; Rodrigues, F.A.; Schurt, D.A.; Resende, R.S.; Souza, N.F.A.; Rezende, D.C.; Moreira, W.R.; Zambolim, L. Effect of foliar-applied potassium silicate on coffee leaf infection by Hemileia vastatrix. Ann. Appl. Biol. 2014, 164, 396–403. [Google Scholar] [CrossRef]
- Monteiro, A.C.A.; de Resende, M.L.V.; Valente, T.C.T.; Ribeiro Junior, P.M.; Pereira, V.F.; da Costa, J.R.; da Silva, J.A.G. Manganese phosphite in coffee defence against Hemileia vastatrix, the coffee rust fungus: Biochemical and molecular analyses. J. Phytopathol. 2016, 164, 1043–1053. [Google Scholar] [CrossRef] [Green Version]
- Costa, B.H.G.; de Resende, M.L.V.; Ribeiro Júnior, P.M.; Mathioni, S.M.; Pádua, M.A.; da Silva Júnior, M.B. Suppression of rust and brown eye spot diseases on coffee by phosphites and by-products of coffee and citrus industries. J. Phytopathol. 2014, 162, 635–642. [Google Scholar] [CrossRef]
- Silva, J.A.G.; Resende, M.L.V.; Monteiro, A.C.A.; Pádua, M.A.; Guerra-Guimarães, L.; Medeiros, F.L.; Martins, S.A.; Botelho, D.M.S. Resistance inducers applied alone or in association with fungicide for the management of leaf rust and brown eye spot of coffee under field conditions. J. Phytopathol. 2019, 167, 430–439. [Google Scholar] [CrossRef]
- Possa, K.F.; Silva, J.A.G.; Resende, M.L.V.; Tenente, R.; Pinheiro, C.; Chaves, I.; Planchon, S.; Monteiro, A.C.A.; Renaut, J.; Carvalho, M.A.F.; et al. Primary metabolism is distinctly modulated by plant resistance inducers in Coffea arabica leaves infected by Hemileia vastatrix. Front. Plant Sci. 2020, 11, 309. [Google Scholar] [CrossRef]
- De Nardi, B.; Dreos, R.; Del Terra, L.; Martellossi, C.; Asquini, E.; Tornincasa, P.; Gasperini, D.; Pacchioni, B.; Rathinavelu, R.; Pallavicini, A.; et al. Differential responses of Coffea arabica L. leaves and roots to chemically induced systemic acquired resistance. Genome 2006, 49, 1594–1605. [Google Scholar] [CrossRef]
- Fernandes, L.H.M.; de Oliveira Silveira, H.R.; de Souza, K.R.; de Resende, M.L.; Alves, J.D. Inductors of resistance and their role in photosynthesis and antioxidant system activity of coffee seedlings. Am. J. Plant Sci. 2014, 5, 3710–3716. [Google Scholar] [CrossRef] [Green Version]
- Resende, M.L.V.; Nojosa, G.B.A.; Cavalcanti, L.S.; Aguilar, M.A.G.; Silva, L.H.C.P.; Perez, J.O.; Andrade, G.C.G.; Carvalho, G.A.; Castro, R.M. Induction of resistance in cocoa against Crinipellis perniciosa and Verticillium dahliae by acibenzolar-S-methyl (ASM). Plant Pathol. 2002, 51, 621–628. [Google Scholar] [CrossRef]
- Chaves, I.; Barros, D.R.; Rodrigues, A.; Silva, M.C.; Ricardo, C.P.; Miguel, C.; Guerra-Guimarães, L. Identification of miRNAs as mediators of Coffea arabica resistance to Hemileia vastatrix. In Proceedings of the 25th International Conference on Coffee Science (ASIC), Armenia, Colombia, 8–13 September 2014; p. 219. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Research Fields | Coffee Defense Responses | References |
|---|---|---|
| Cytology | ROS accumulation in the cell walls and cytoplasmic contents | [136,143,144] |
| Deployment of hypersensitive response (HR) | [52,136,137,140,141,142] | |
| Accumulation of phenolic-like compounds (Cell walls and cytoplasmic contents) | [52,136,137,140] | |
| Lignification of cell walls | [52,137] | |
| Haustorium encasement with callose and β-1,4-glucans | [52,137] | |
| Cell hypertrophy | [20,52,137] | |
| Accumulation of a material partially crystallised (containing pectins, polysaccharides and phenolic-like compounds) in the intercellular spaces | [52,137] | |
| Analytical chemistry | Increase in salicylic acid (SA) levels | [145] |
| Increase of chlorogenic acids (4,5-diCQA) content | [146] | |
| Biochemistry | Increase in chitinases and glucanases activity | [147,148] |
| Immunodetection of class I chitinases | [147] | |
| Increase in peroxidase (POD) activity | [136] | |
| Increase in superoxide dismutase (SOD) activity | [139] | |
| Increase in lipoxygenase (LOX) activity | [149] | |
| Increase in phenylalanine ammonia lyase (PAL) activity | [137] |
| Gene | Annotation | Accession Number | Reference |
|---|---|---|---|
| CaRLK | Receptor-like kinase | DSS6; CF589181 | [140,150] |
| NB-ARC domain | NB-ARC domain-containing disease resistance protein | n.a. | [151] |
| SD1-8 | Receptor-like serine/threonine-protein kinase SD1-8 | n.a. | [151] |
| NBS-LRR | Nucleotide-binding site–leucine-rich repeat | DQ123968; GT030058.1 | [152,153] |
| At5g39020 | Putative probable receptor-like protein kinase At5g39020 | n.a. | [151] |
| LRR_RLK2 | Leucine-rich repeat receptor-like protein kinase | Contig_37_g342.t1 | [22] |
| RGH1A | Resistance gene analogs 1A | GT029983.1 | [152] |
| NDR1 | Non-race specific Disease Resistance 1 | DSS12; CO773976 | [138,150,154] |
| CDPK5 | Calcium-dependent protein kinase 5 | GT030068.1 | [152] |
| CBP | Calmodulin-binding protein | GT030070.1 | [152] |
| CaWRKY1 | WRKY transcription factor 1 | DSS16 | [138,140,150,155,156] |
| CaWRKY1a | WRKY transcription factor 1a | n.a. | [156,157] |
| CaWRKY1b | WRKY transcription factor 1b | n.a. | [156,157] |
| CaWRKY3 | WRKY transcription factor 3 | GT715378; GT715379; GT669686; GT684003; GT675420; GT675421; GT675422; GT675423 | [155] |
| CaWRKY6 | WRKY transcription factor 6 | GW460548; GW439927; GT671542; GT700942; GT688469; GT722306 | [155] |
| CaWRKY7 | WRKY transcription factor 7 | n.a. | [155] |
| CaWRKY8 | WRKY transcription factor 8 | GT685360; GW436389 | [155] |
| CaWRKY10 | WRKY transcription factor 10 | DV693086 | [155] |
| CaWRKY11 | WRKY transcription factor 11 | GW489409; GW472658; GW489686; GW467855 | [155] |
| CaWRKY12 | WRKY transcription factor 12 | GW487300; GT715908 | [155] |
| CaWRKY13 | WRKY transcription factor 13 | GW428837; GW469454 | [155] |
| CaWRKY14 | WRKY transcription factor 14 | GT730216; GT688262; GT688263; GT733537 | [155] |
| CaWRKY15 | WRKY transcription factor 15 | GT725660; GT721476; GT721477; GT704531; GT707389; GT702040; GT702041; GW490838; GT695761; GT674787; GT704614 | [155] |
| CaWRKY17 | WRKY transcription factor 17 | DV678740 | [155] |
| CaWRKY19 | WRKY transcription factor 19 | GW490173; GW439047; GW438774; GW472647; GW481654 | [155] |
| CaWRKY20 | WRKY transcription factor 20 | DV706804 | [155] |
| CaWRKY21 | WRKY transcription factor 21 | DV709595.1 | [155] |
| CaWRKY22 | WRKY transcription factor 22 | GT685546; GT684702 | [155] |
| bHLH | Putative basic helix-loop-helix (bHLH) DNA-binding superfamily protein | n.a. | [151] |
| AP2-type transcription factor | AP2-type transcription factor | DSS17 | [150] |
| bZIP56 | Transcription factor bZIP56 | GT030080.1 | [152] |
| MAPK2 | Mitogen-activated protein kinase 2 | GT030062.1 | [152] |
| MEK2 | Dual specificity mitogen-activated protein kinase kinase 2 | GT030000.1 | [152] |
| 13-LOX | Lipoxygenase 13 | DV704189 | [140,142] |
| LOX | Lipoxygenase | DQ123948 | [153] |
| CaPAL | Phenylalanine ammonia lyase | DQ067599; JF838179 | [140] |
| NPR1 | Non-expressor of pathogenesis-related gene | n.a. | [154] |
| CaGT | Salicylic acid-glucosyltransferase | DSS22; CO773975 | [140,150] |
| ERF | Ethylene-responsive transcription factor 1B | n.a. | [151] |
| ABC | PDR-type ABC transporter 1 | DQ123937 | [153] |
| PR1 | Pathogenesis-related protein 1 | n.a. | [154] |
| CaPR1b | Pathogenesis-related protein 1b | DQ335594 | [140,142,143] |
| PR2 | Pathogenesis-related protein 2 (beta-1,3-glucanase) | DQ123927 | [153] |
| PR5 | Pathogen-related protein 5 (thaumatin-like gene) | GT030025.1 | [152] |
| CaPR10 | Pathogenesis-related protein 10 | CF589103 | [140,143] |
| PNC2 | Cationic peroxidase 2 | n.a. | [151] |
| R1A-6 | Putative late blight resistance protein homolog R1A-6 | n.a. | [151] |
| RGA1 | Putative disease resistance protein RGA1 | n.a. | [151] |
| PUB24 | Putative E3 ubiquitin-protein ligase PUB24 | n.a. | [151] |
| ITN1 | Putative ankyrin repeat-containing protein At3g12360 | n.a. | [151] |
| Dirigent-like protein | Putative disease resistance-responsive (dirigent- like protein) family protein | n.a. | [151] |
| Premnaspiridione oxygenase | Premnaspiridione oxygenase | n.a. | [151] |
| CyP450 | Cytochrome P450 | DSS10 | [150] |
| HSP70 | 70-kDa heat shock proteins | DSS11; DSS16; CO773974; DQ335599 | [150] |
| CaR111 | Unknown function | DSS23; CF589193 | [138,150] |
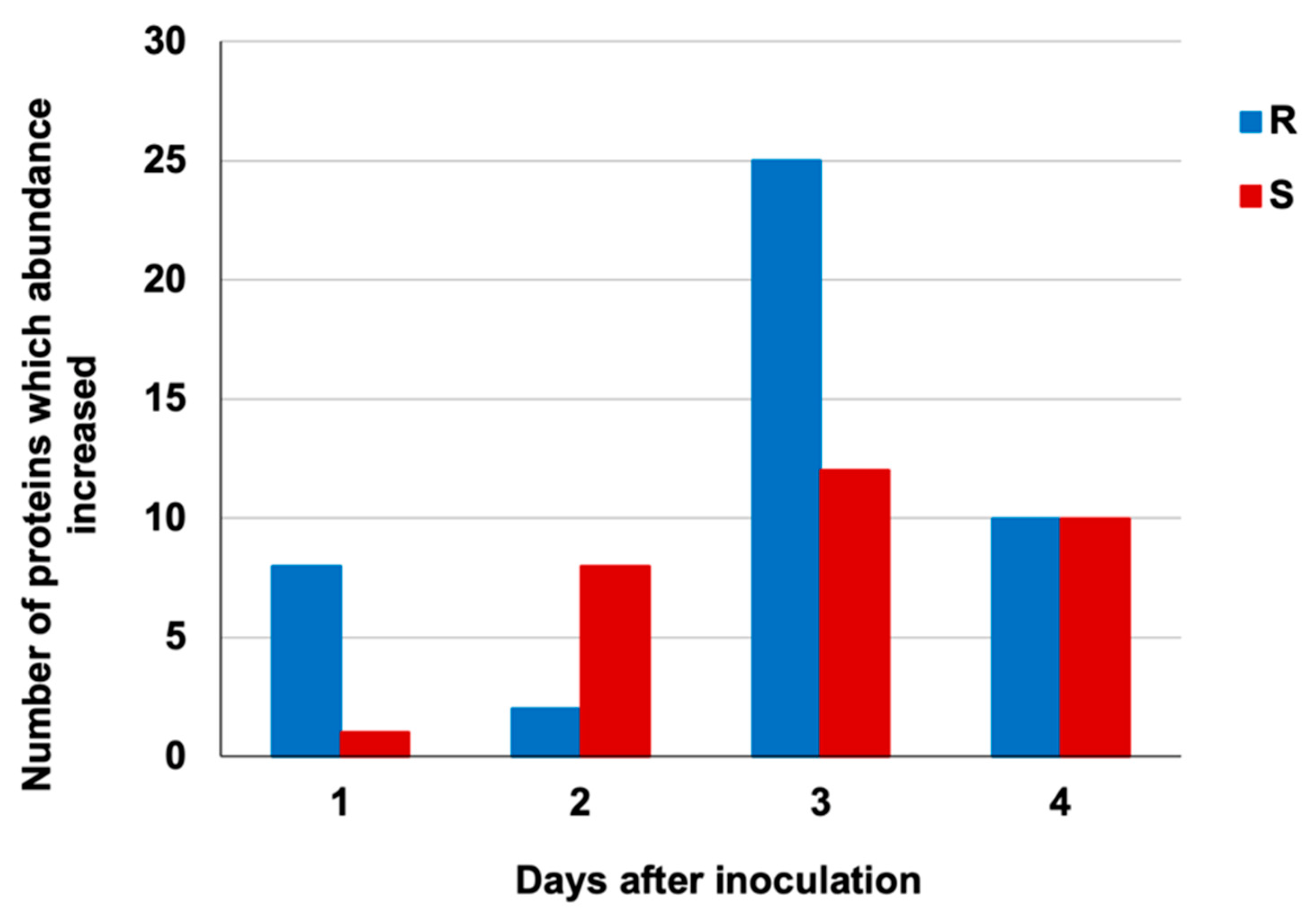
| d.a.i | Protein Identity a | Superfamily b | Functional Annotation c | Number of Spots |
|---|---|---|---|---|
| 1 | Calcineurin-like phosphoesterase | Metallophosphatases | Miscellaneous enzymes | 1 |
| Cysteine proteinase aleuran type | Peptidase_C1 | Protein degradation | 1 | |
| Subtilisin-like protease | Peptidases_S8_S54 | Protein degradation | 2 | |
| Berberine bridge enzyme | FAD_binding | Secondary metabolism | 1 | |
| Osmotin | GH64-Thaumatin-like | Stress/Defense | 1 | |
| Cysteine-rich repeat secretory protein 55-like | Stress-antifungal | Stress/Defense | 1 | |
| Chitinase-like protein | GH18_chitinase-like | Stress/Defense | 1 | |
| 2 | Chitinase-like protein | GH18_chitinase-like | Stress/Defense | 2 |
| 3 | Beta-xylosidase/alpha-L-Arabinofuranosidase | GH3 | Cell wall degradation | 7 |
| Pectin methylesterase | Pectinesterase | Cell wall | 1 | |
| Pectinesterase-like | Pectinesterase | Cell wall | 1 | |
| purple acid phosphatase | Metallophosphatases | Miscellaneous enzymes | 1 | |
| Aspartic proteinase nepenthesin-1-like | pepsin_retropepsin | Protein degradation | 2 | |
| Serine carboxypeptidase | Peptidase_S10 | Protein degradation | 4 | |
| Subtilisin-like protease | Peptidases_S8_S53 | Protein degradation | 6 | |
| Chitinase-like protein | GH18_chitinase-like | Stress/Defense | 1 | |
| Pathogenesis-related protein 5-like | GH64-Thaumatin-like | Stress/Defense | 1 | |
| 4 | Beta-xylosidase/alpha-L-Arabinofuranosidase | GH3 | Cell wall degradation | 1 |
| Alpha-L-fucosidase | Alpha-amylase | Cell wall degradation | 2 | |
| Non-cell-autonomous protein pathway | Aldose_epim | Minor CHO metabolism | 1 | |
| Lysosomal alpha-mannosidase-like | GH38 | Miscellaneous enzymes | 2 | |
| Alpha-xylosidase 1 | GH31 | Miscellaneous enzymes | 1 | |
| ASPARTIC PROTEASE IN GUARD CELL-like protein | pepsin_retropepsin | Protein degradation | 2 | |
| Germin-like protein | Cupin | Stress/Defense | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, M.d.C.; Guerra-Guimarães, L.; Diniz, I.; Loureiro, A.; Azinheira, H.; Pereira, A.P.; Tavares, S.; Batista, D.; Várzea, V. An Overview of the Mechanisms Involved in Coffee-Hemileia vastatrix Interactions: Plant and Pathogen Perspectives. Agronomy 2022, 12, 326. https://doi.org/10.3390/agronomy12020326
Silva MdC, Guerra-Guimarães L, Diniz I, Loureiro A, Azinheira H, Pereira AP, Tavares S, Batista D, Várzea V. An Overview of the Mechanisms Involved in Coffee-Hemileia vastatrix Interactions: Plant and Pathogen Perspectives. Agronomy. 2022; 12(2):326. https://doi.org/10.3390/agronomy12020326
Chicago/Turabian StyleSilva, Maria do Céu, Leonor Guerra-Guimarães, Inês Diniz, Andreia Loureiro, Helena Azinheira, Ana P. Pereira, Sílvia Tavares, Dora Batista, and Vítor Várzea. 2022. "An Overview of the Mechanisms Involved in Coffee-Hemileia vastatrix Interactions: Plant and Pathogen Perspectives" Agronomy 12, no. 2: 326. https://doi.org/10.3390/agronomy12020326