
Root Growth and Architecture of Wheat and Brachypodium Vary in Response to Algal Fertilizer in Soil and Solution

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
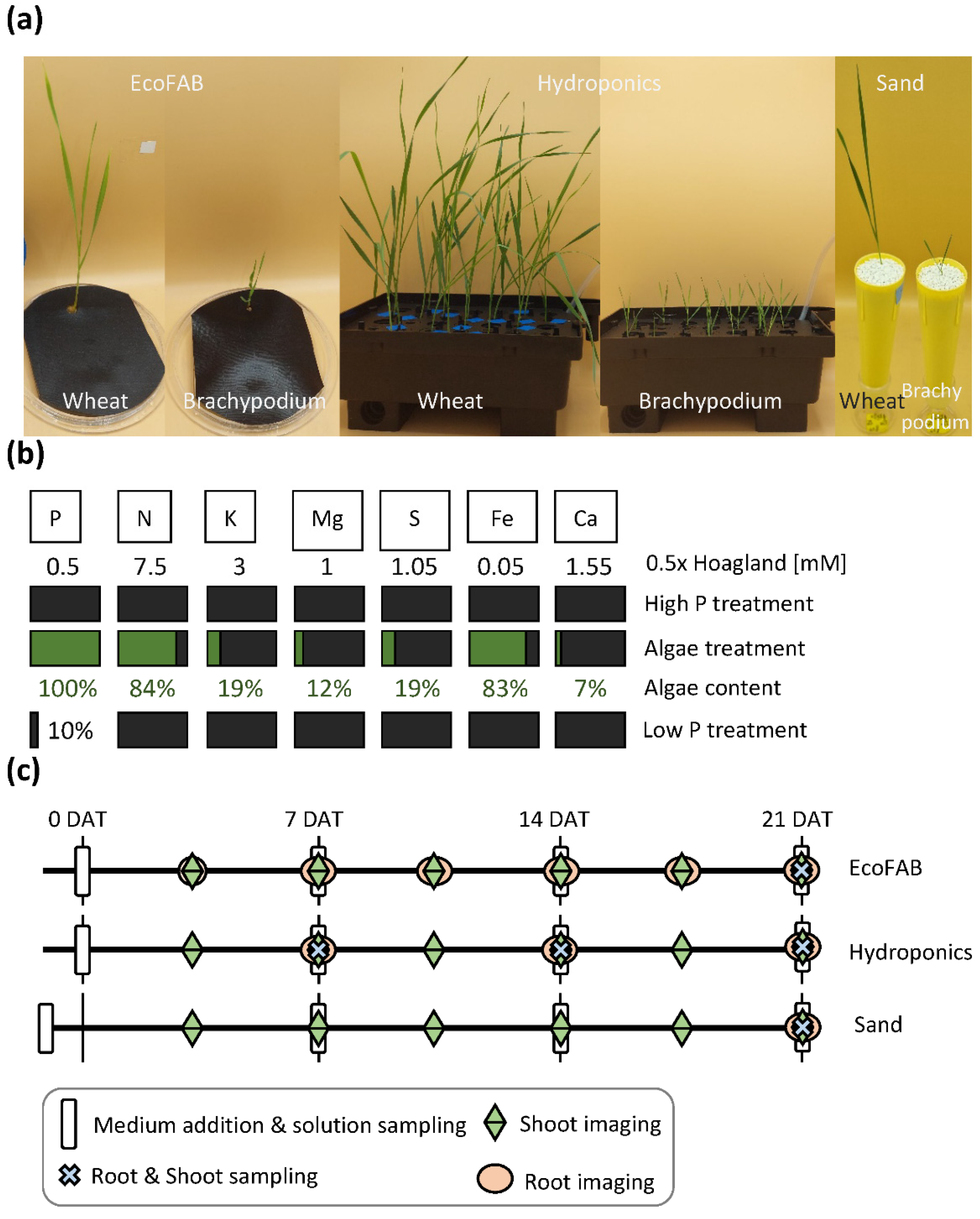
2.1. Plant Material
2.2. Experimental Setups and Growth Conditions
2.2.1. Nutrient Treatments
2.2.2. EcoFAB
2.2.3. Hydroponics
2.2.4. Sand
2.3. Measurements
2.3.1. Non-Invasive Phenotyping
2.3.2. Invasive Phenotyping
2.3.3. Nutrient Content Determination
2.4. Statistical Analyses
3. Results
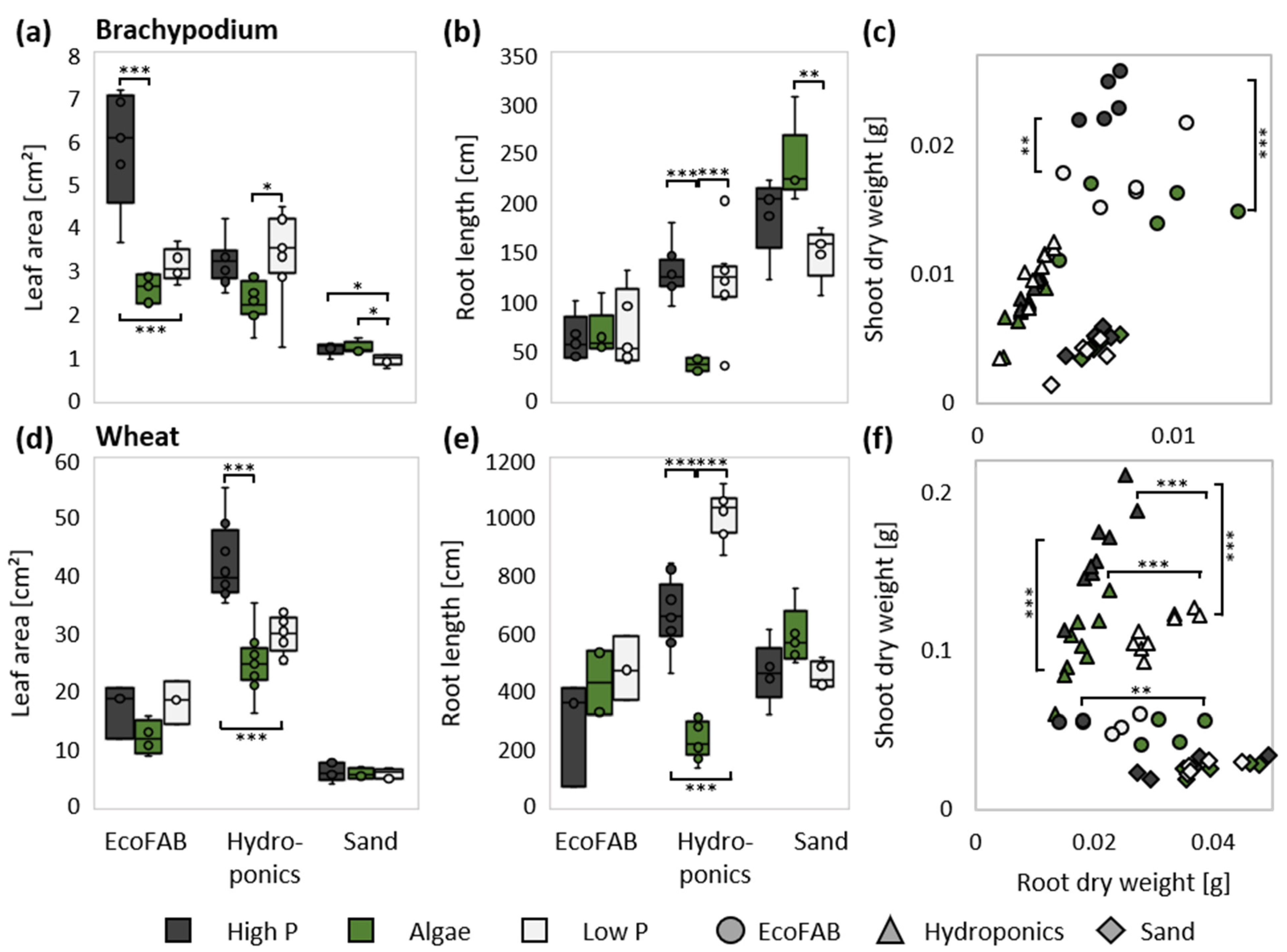
3.1. Wheat and Brachypodium Took Up P from Algal Biomass
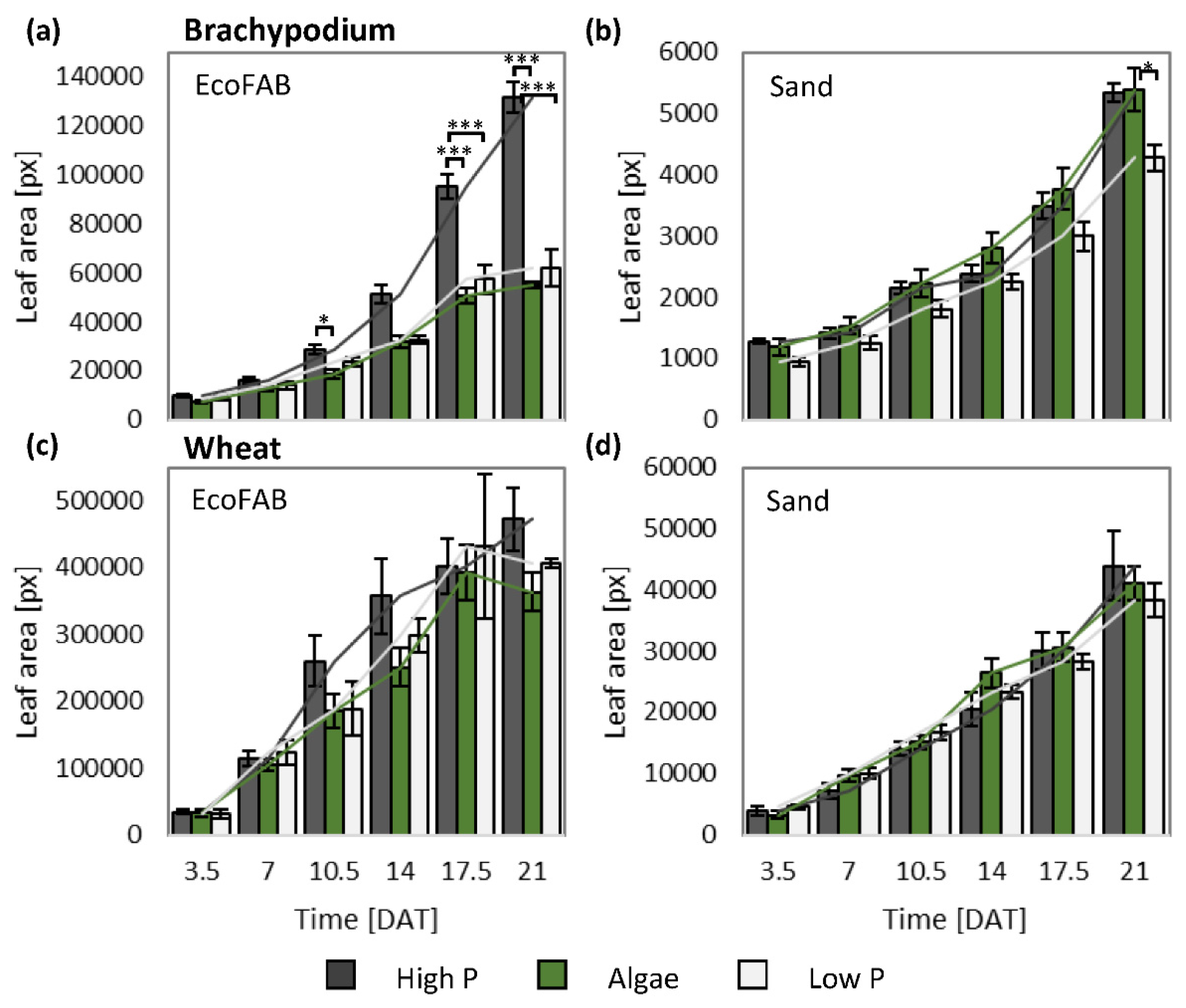
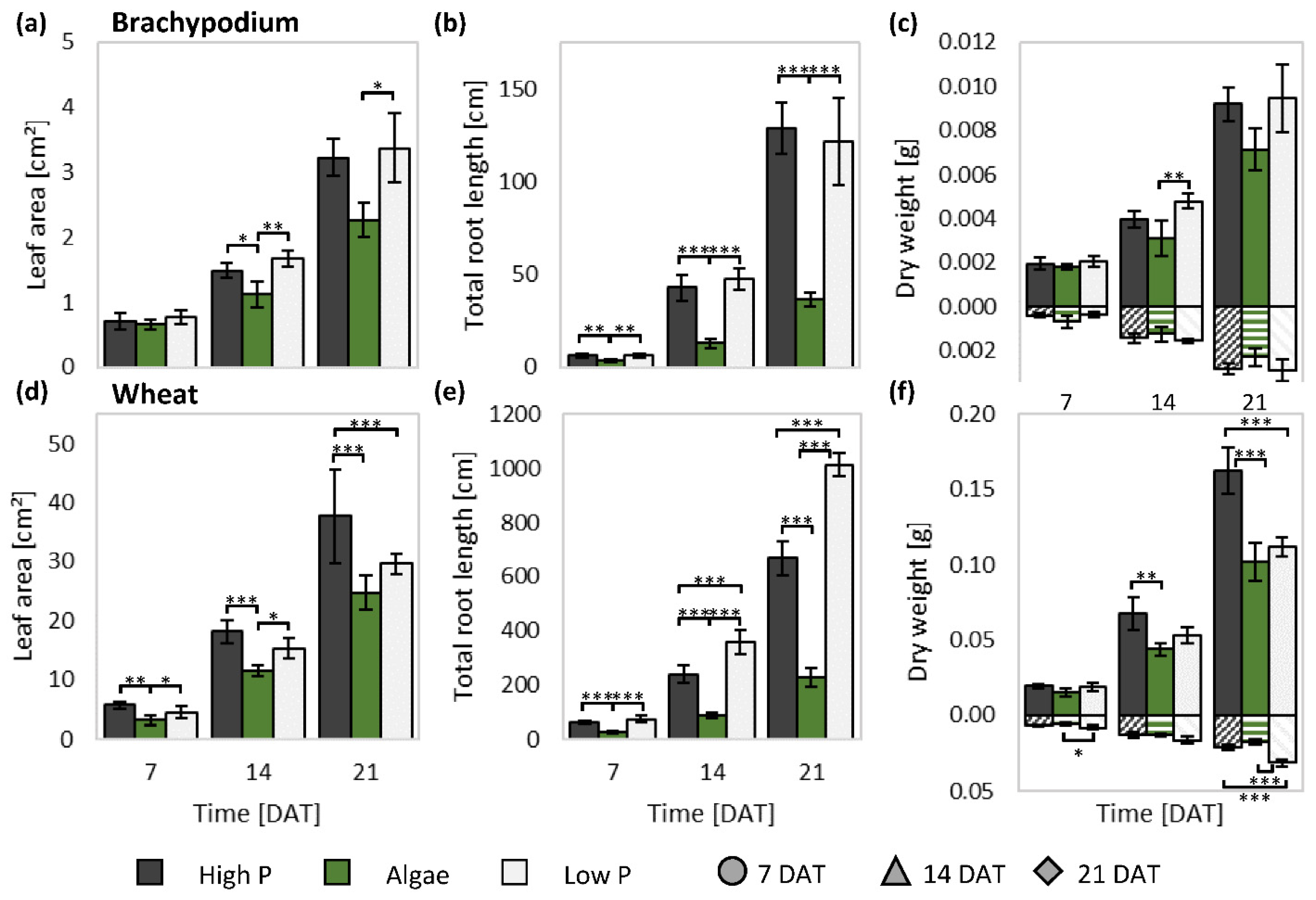
3.2. Non-Invasive and Invasive Measurements Allowed Dynamic Phenotyping
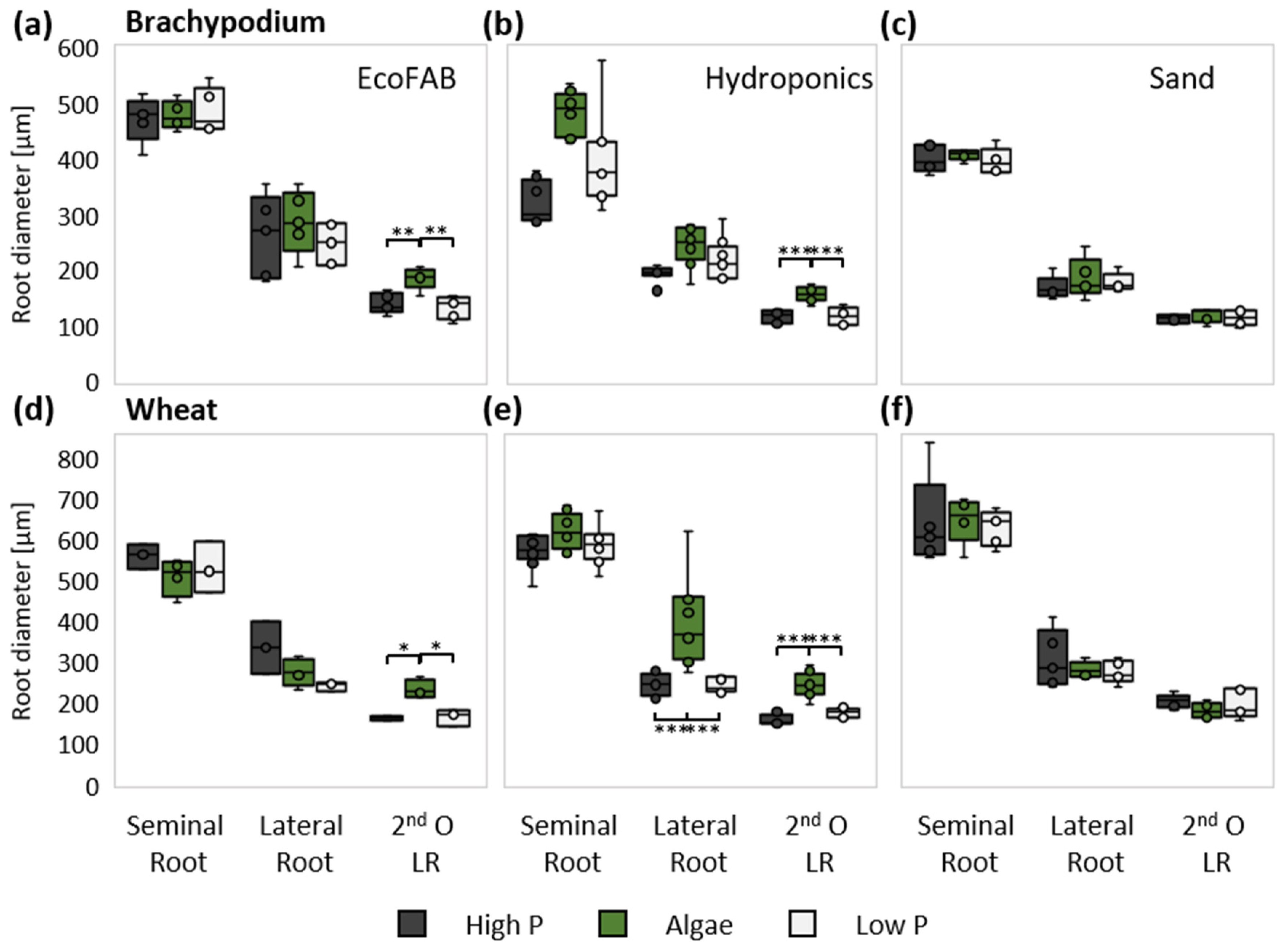
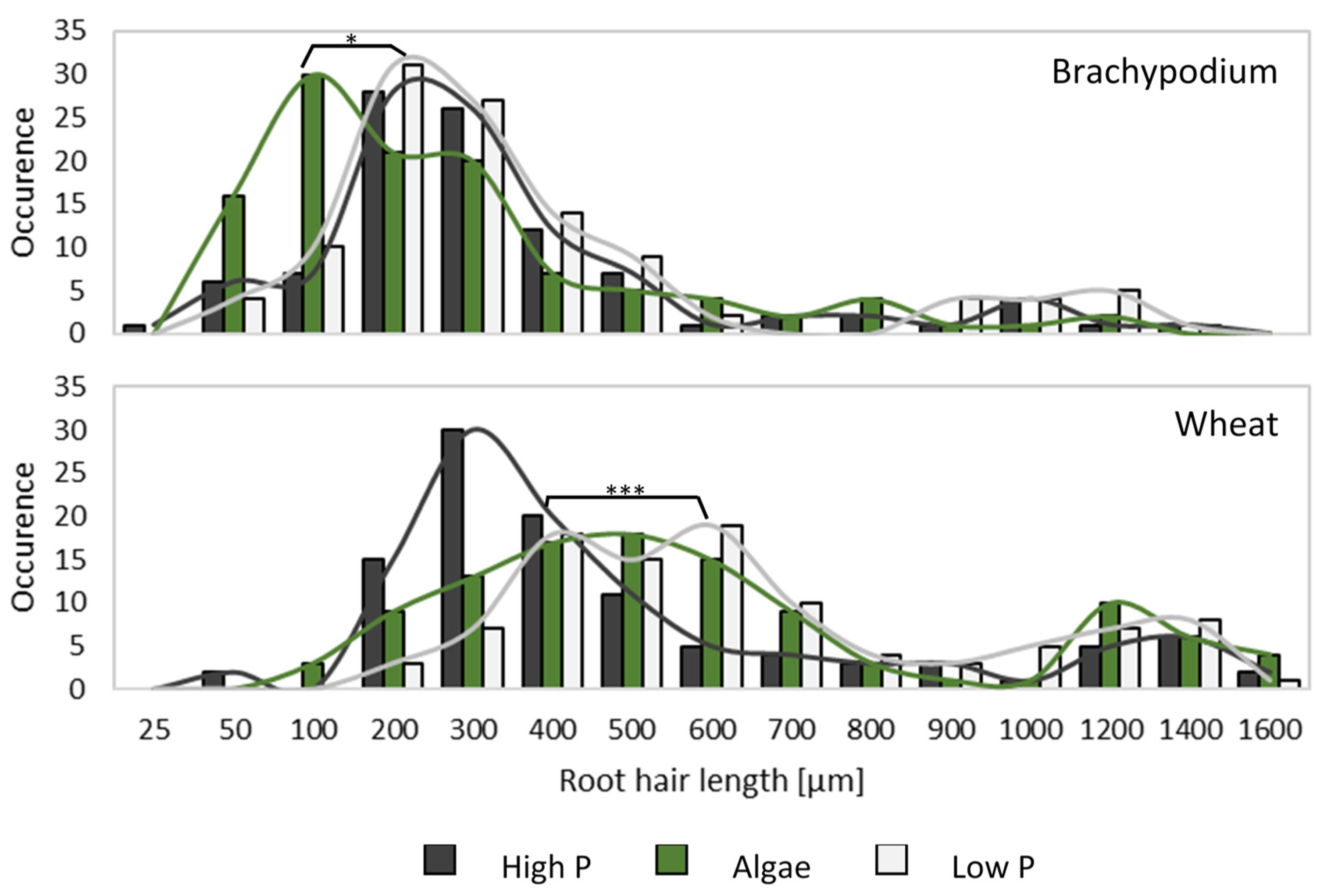
3.3. Presence of Algae Changed Root Morphology in Wheat and Brachypodium
4. Discussion
4.1. Wheat and Brachypodium Are Taking Up P from Algal Biomass
4.2. P Efficiency in Three Inherently Different Systems
4.3. Algal Biomass Impacts Root Development and Medium pH
4.4. Nutrient Supply by Algae Is Complementing Mineral Nutrition
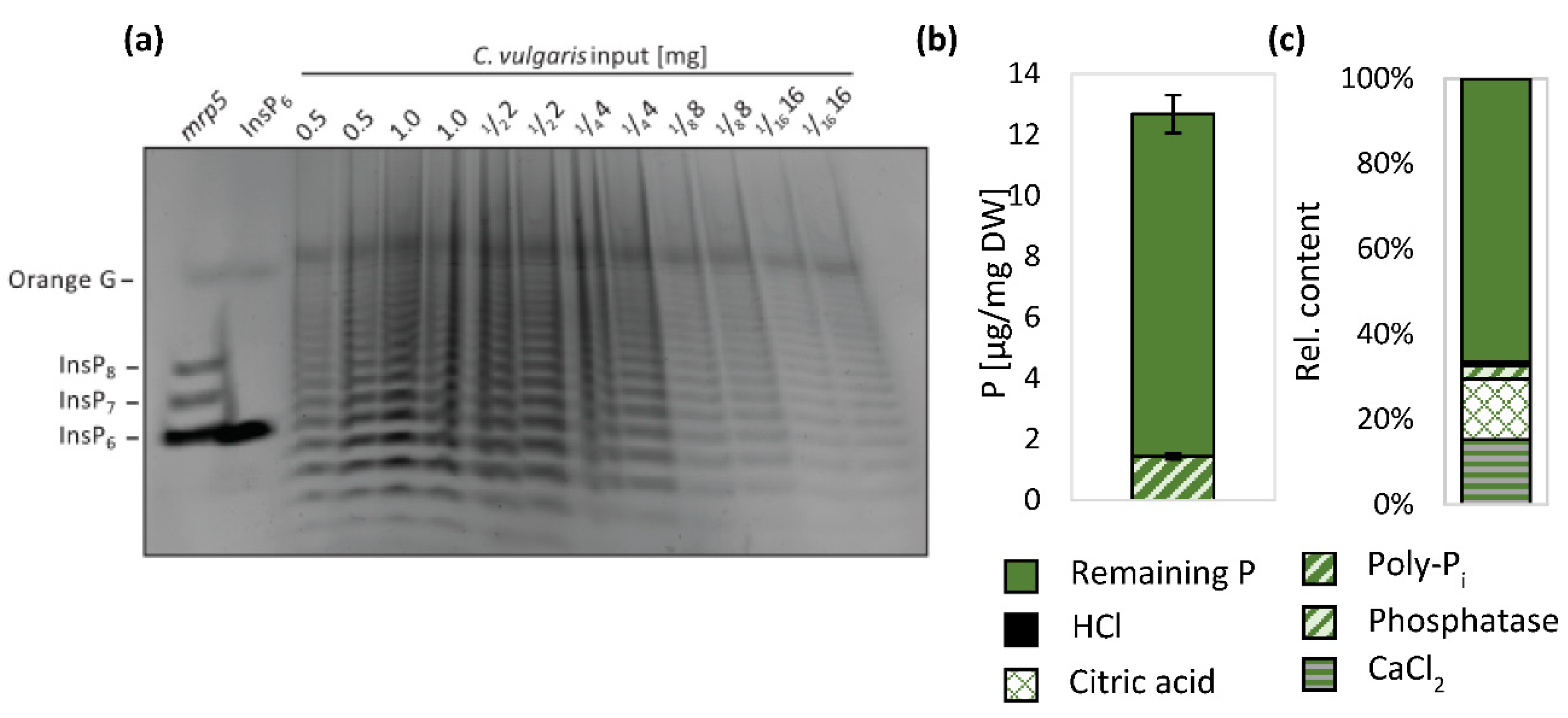
4.5. P-Form Complexity within Algae Biomass Offers Potential for Slow Release P Fertilizer
4.6. Wheat and Brachypodium Have a Divergent Shoot, but Similar Root Response to Algal Biomass
4.7. Future Research for Renewable Fertilizer Use by Crop
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Solovchenko, A.; Verschoor, A.M.; Jablonowski, N.D.; Nedbal, L. Phosphorus from wastewater to crops: An alternative path involving microalgae. Biotechnol. Adv. 2016, 34, 550–564. [Google Scholar] [CrossRef]
- Ismagulova, T.; Shebanova, A.; Gorelova, O.; Baulina, O.; Solovchenko, A. A new simple method for quantification and locating P and N reserves in microalgal cells based on energy-filtered transmission electron microscopy (EFTEM) elemental maps. PLoS ONE 2018, 13, e0208830. [Google Scholar] [CrossRef] [PubMed]
- Moudříková, Š.; Nedbal, L.; Solovchenko, A.; Mojzeš, P. Raman microscopy shows that nitrogen-rich cellular inclusions in microalgae are microcrystalline guanine. Algal Res. 2017, 23, 216–222. [Google Scholar] [CrossRef]
- Mojzeš, P.; Gao, L.; Ismagulova, T.; Pilátová, J.; Moudříková, Š.; Gorelova, O.; Solovchenko, A.; Nedbal, L.; Salih, A. Guanine, a high-capacity and rapid-turnover nitrogen reserve in microalgal cells. Proc. Natl. Acad. Sci. USA 2020, 117, 32722–32730. [Google Scholar] [CrossRef] [PubMed]
- Miyachi, S.; Kanai, R.; Mihara, S.; Aoki, S. Metabolic roles of inorganic polyphosphates in chlorella cells. Biochim. Biophys. Acta 1964, 93, 625–634. [Google Scholar] [CrossRef]
- Brown, N.; Shilton, A. Luxury uptake of phosphorus by microalgae in waste stabilisation ponds: Current understanding and future direction. Rev. Environ. Sci. Bio/Technol. 2014, 13, 321–328. [Google Scholar] [CrossRef]
- Moudříková, Š.; Ivanov, I.N.; Vítová, M.; Nedbal, L.; Zachleder, V.; Mojzeš, P.; Bišová, K. Comparing Biochemical and Raman Microscopy Analyses of Starch, Lipids, Polyphosphate, and Guanine Pools during the Cell Cycle of Desmodesmus quadricauda. Cells 2021, 10, 62. [Google Scholar] [CrossRef]
- Couso, I.; Evans, B.S.; Li, J.; Liu, Y.; Ma, F.; Diamond, S.; Allen, D.K.; Umen, J.G. Synergism between Inositol Polyphosphates and TOR Kinase Signaling in Nutrient Sensing, Growth Control, and Lipid Metabolism in Chlamydomonas. Plant Cell 2016, 28, 2026–2042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raboy, V. myo-Inositol-1,2,3,4,5,6-hexakisphosphate. Phytochemistry 2003, 64, 1033–1043. [Google Scholar] [CrossRef]
- Zhu, J.; Lau, K.; Puschmann, R.; Harmel, R.K.; Zhang, Y.; Pries, V.; Gaugler, P.; Broger, L.; Dutta, A.K.; Jessen, H.J.; et al. Two bifunctional inositol pyrophosphate kinases/phosphatases control plant phosphate homeostasis. eLife 2019, 8, e43582. [Google Scholar] [CrossRef]
- Riemer, E.; Qiu, D.; Laha, D.; Harmel, R.K.; Gaugler, P.; Gaugler, V.; Frei, M.; Hajirezaei, M.-R.; Laha, N.P.; Krusenbaum, L.; et al. ITPK1 is an InsP6/ADP phosphotransferase that controls phosphate signaling in Arabidopsis. Mol. Plant 2021, 14, 1864–1880. [Google Scholar] [CrossRef]
- Laha, D.; Johnen, P.; Azevedo, C.; Dynowski, M.; Weiß, M.; Capolicchio, S.; Mao, H.; Iven, T.; Steenbergen, M.; Freyer, M.; et al. VIH2 Regulates the Synthesis of Inositol Pyrophosphate InsP8 and Jasmonate-Dependent Defenses in Arabidopsis. Plant Cell 2015, 27, 1082–1097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blüher, D.; Laha, D.; Thieme, S.; Hofer, A.; Eschen-Lippold, L.; Masch, A.; Balcke, G.; Pavlovic, I.; Nagel, O.; Schonsky, A.; et al. A 1-phytase type III effector interferes with plant hormone signaling. Nat. Commun. 2017, 8, 2159. [Google Scholar] [CrossRef]
- Laha, N.P.; Dhir, Y.W.; Giehl, R.F.H.; Schäfer, E.M.; Gaugler, P.; Shishavan, Z.H.; Gulabani, H.; Mao, H.; Zheng, N.; von Wirén, N.; et al. ITPK1-Dependent Inositol Polyphosphates Regulate Auxin Responses in Arabidopsis thaliana. bioRXiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Gulabani, H.; Goswami, K.; Walia, Y.; Roy, A.; Noor, J.J.; Ingole, K.D.; Kasera, M.; Laha, D.; Giehl, R.F.H.; Schaaf, G.; et al. Arabidopsis inositol polyphosphate kinases IPK1 and ITPK1 modulate crosstalk between SA-dependent immunity and phosphate-starvation responses. Plant Cell Rep 2021. [Google Scholar] [CrossRef] [PubMed]
- Mau, L.; Kant, J.; Walker, R.; Kuchendorf, C.M.; Schrey, S.D.; Roessner, U.; Watt, M. Wheat Can Access Phosphorus From Algal Biomass as Quickly and Continuously as From Mineral Fertilizer. Front. Plant Sci. 2021, 12, 631314. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, C.; Schiedung, H.; Harrison, L.; Briese, C.; Ackermann, B.; Kant, J.; Schrey, S.D.; Hofmann, D.; Singh, D.; Ebenhöh, O.; et al. Evaluating potential of green alga Chlorella vulgaris to accumulate phosphorus and to fertilize nutrient-poor soil substrates for crop plants. J. Appl. Phycol. 2018, 30, 2827–2836. [Google Scholar] [CrossRef]
- Siebers, N.; Hofmann, D.; Schiedung, H.; Landsrath, A.; Ackermann, B.; Gao, L.; Mojzeš, P.; Jablonowski, N.D.; Nedbal, L.; Amelung, W. Towards phosphorus recycling for agriculture by algae: Soil incubation and rhizotron studies using 33P-labeled microalgal biomass. Algal Res. 2019, 43, 101634. [Google Scholar] [CrossRef]
- Richardson, A.E.; Hadobas, P.A.; Hayes, J.E. Acid phosphomonoesterase and phytase activities of wheat (Triticum aestivum L.) roots and utilization of organic phosphorus substrates by seedlings grown in sterile culture. Plant Cell Environ. 2000, 23, 397–405. [Google Scholar] [CrossRef]
- Torres-Dorante, L.O.; Claassen, N.; Steingrobe, B.; Olfs, H.-W. Hydrolysis rates of inorganic polyphosphates in aqueous solution as well as in soils and effects on P availability. J. Plant Nutr. Soil Sci. 2005, 168, 352–358. [Google Scholar] [CrossRef]
- Mukherjee, C.; Chowdhury, R.; Ray, K. Phosphorus Recycling from an Unexplored Source by Polyphosphate Accumulating Microalgae and Cyanobacteria—A Step to Phosphorus Security in Agriculture. Front. Microbiol. 2015, 6, 1421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FAOstat. Food and Agriculture Organization of the United Nations, 2019. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 11 December 2021).
- Appels, R.; Eversole, K.; Stein, N.; Feuillet, C.; Keller, B.; Rogers, J.; Pozniak, C.J.; Choulet, F.; Distelfeld, A.; Poland, J.; et al. Shifting the limits in wheat research and breeding using a fully annotated reference genome. Science 2018, 361, eaar7191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watt, M.; Magee, L.J.; McCully, M.E. Types, structure and potential for axial water flow in the deepest roots of field-grown cereals. New Phytol. 2008, 178, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Vogel, J.P.; Garvin, D.F.; Mockler, T.C.; Schmutz, J.; Rokhsar, D.; Bevan, M.W.; Barry, K.; Lucas, S.; Harmon-Smith, M.; Lail, K.; et al. Genome sequencing and analysis of the model grass Brachypodium distachyon. Nature 2010, 463, 763–768. [Google Scholar] [CrossRef]
- Watt, M.; Schneebeli, K.; Dong, P.; Wilson, I.W. The shoot and root growth of Brachypodium and its potential as a model for wheat and other cereal crops. Funct. Plant Biol. 2009, 36, 960–969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawasaki, A.; Donn, S.; Ryan, P.R.; Mathesius, U.; Devilla, R.; Jones, A.; Watt, M. Microbiome and Exudates of the Root and Rhizosphere of Brachypodium distachyon, a Model for Wheat. PLoS ONE 2016, 11, e0164533. [Google Scholar] [CrossRef] [Green Version]
- Chochois, V.; Vogel, J.P.; Watt, M. Application of Brachypodium to the genetic improvement of wheat roots. J. Exp. Bot. 2012, 63, 3467–3474. [Google Scholar] [CrossRef] [Green Version]
- Poiré, R.; Chochois, V.; Sirault, X.R.R.; Vogel, J.P.; Watt, M.; Furbank, R.T. Digital imaging approaches for phenotyping whole plant nitrogen and phosphorus response in Brachypodium distachyon. J. Integr. Plant Biol. 2014, 56, 781–796. [Google Scholar] [CrossRef]
- Chochois, V.; Vogel, J.P.; Rebetzke, G.J.; Watt, M. Variation in Adult Plant Phenotypes and Partitioning among Seed and Stem-Borne Roots across Brachypodium distachyon Accessions to Exploit in Breeding Cereals for Well-Watered and Drought Environments. Plant Physiol. 2015, 168, 953–967. [Google Scholar] [CrossRef] [Green Version]
- Hoagland, D.R.; Arnon, D.I. The water-culture method for growing plants without soil. Circular. Calif. Agric. Exp. Stn. 1950, 347, 32. [Google Scholar]
- Gao, J.; Sasse, J.; Lewald, K.M.; Zhalnina, K.; Cornmesser, L.T.; Duncombe, T.A.; Yoshikuni, Y.; Vogel, J.P.; Firestone, M.K.; Northen, T.R. Ecosystem Fabrication (EcoFAB) Protocols for The Construction of Laboratory Ecosystems Designed to Study Plant-microbe Interactions. JoVE 2018, 134, e57170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasse, J.; Kant, J.; Cole, B.J.; Klein, A.P.; Arsova, B.; Schlaepfer, P.; Gao, J.; Lewald, K.; Zhalnina, K.; Kosina, S.; et al. Multilab EcoFAB study shows highly reproducible physiology and depletion of soil metabolites by a model grass. New Phytol. 2019, 222, 1149–1160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller-Linow, M.; Wilhelm, J.; Briese, C.; Wojciechowski, T.; Schurr, U.; Fiorani, F. Plant Screen Mobile: An open-source mobile device app for plant trait analysis. Plant Methods 2019, 15, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nestler, J.; Keyes, S.D.; Wissuwa, M. Root hair formation in rice (Oryza sativa L.) differs between root types and is altered in artificial growth conditions. J. Exp. Bot. 2016, 67, 3699–3708. [Google Scholar] [CrossRef] [Green Version]
- Wilson, M.S.C.; Bulley, S.J.; Pisani, F.; Irvine, R.F.; Saiardi, A. A novel method for the purification of inositol phosphates from biological samples reveals that no phytate is present in human plasma or urine. Open Biol. 2015, 5, 150014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Losito, O.; Szijgyarto, Z.; Resnick, A.C.; Saiardi, A. Inositol Pyrophosphates and Their Unique Metabolic Complexity: Analysis by Gel Electrophoresis. PLoS ONE 2009, 4, e5580. [Google Scholar] [CrossRef]
- DeLuca, T.H.; Glanville, H.C.; Harris, M.; Emmett, B.A.; Pingree, M.R.A.; de Sosa, L.L.; Cerdá-Moreno, C.; Jones, D.L. A novel biologically-based approach to evaluating soil phosphorus availability across complex landscapes. Soil Biol. Biochem. 2015, 88, 110–119. [Google Scholar] [CrossRef] [Green Version]
- McKercher, R.B.; Tollefson, T.S. Barley response to phosphorus from phospholipids and nucleic acids. Can. J. Soil Sci. 1978, 58, 103–105. [Google Scholar] [CrossRef]
- Nguyen, T.D.P.; Frappart, M.; Jaouen, P.; Pruvost, J.; Bourseau, P. Harvesting Chlorella vulgaris by natural increase in pH: Effect of medium composition. Environ. Technol. 2014, 35, 1378–1388. [Google Scholar] [CrossRef]
- Otsuki, A.; Hanya, T. Production of dissolved organic matter from dead green algal cells. II. Anaerobic microbial decomposition. Limnol. Oceanogr. 1972, 17, 258–264. [Google Scholar] [CrossRef]
- Blossfeld, S.; Schreiber, C.M.; Liebsch, G.; Kuhn, A.J.; Hinsinger, P. Quantitative imaging of rhizosphere pH and CO2 dynamics with planar optodes. Ann. Bot. 2013, 112, 267–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talboys, P.J.; Healey, J.R.; Withers, P.J.A.; Jones, D.L. Phosphate depletion modulates auxin transport in Triticum aestivum leading to altered root branching. J. Exp. Bot. 2014, 65, 5023–5032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jirásková, D.; Poulíčková, A.; Novák, O.; Sedláková, K.; Hradecká, V.; Strnad, M. High-throughput screening technology for monitoring phytohormone production in microalgae. J. Phycol. 2009, 45, 108–118. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Krogstad, T.; Clarke, J.L.; Hallama, M.; Øgaard, A.F.; Eich-Greatorex, S.; Kandeler, E.; Clarke, N. Rhizosphere Organic Anions Play a Minor Role in Improving Crop Species’ Ability to Take Up Residual Phosphorus (P) in Agricultural Soils Low in P Availability. Front. Plant Sci. 2016, 7, 1664. [Google Scholar] [CrossRef] [Green Version]
- Raven, J.A.; Lambers, H.; Smith, S.E.; Westoby, M. Costs of acquiring phosphorus by vascular land plants: Patterns and implications for plant coexistence. New Phytol. 2018, 217, 1420–1427. [Google Scholar] [CrossRef] [Green Version]
- Nabel, M.; Schrey, S.D.; Poorter, H.; Koller, R.; Nagel, K.A.; Temperton, V.M.; Dietrich, C.C.; Briese, C.; Jablonowski, N.D. Coming Late for Dinner: Localized Digestate Depot Fertilization for Extensive Cultivation of Marginal Soil with Sida hermaphrodita. Front. Plant Sci. 2018, 9, 1095. [Google Scholar] [CrossRef]
- Khan, Z.; Karamahmutoğlu, H.; Elitaş, M.; Yüce, M.; Budak, H. Through the Looking Glass: Real-Time Imaging in Brachypodium Roots and Osmotic Stress Analysis. Plants 2019, 8, 14. [Google Scholar] [CrossRef] [Green Version]
- Lennard, W.; Goddek, S. Aquaponics: The Basics. In Aquaponics Food Production Systems: Combined Aquaculture and Hydroponic Production Technologies for the Future; Goddek, S., Joyce, A., Kotzen, B., Burnell, G.M., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 113–143. [Google Scholar]
- Tang, C.; Kuo, J.; Longnecker, N.E.; Thomson, C.J.; Robson, A.D. High pH Causes Disintegration of the Root Surface in Lupinus angustifolius L. Ann. Bot. 1993, 71, 201–207. [Google Scholar] [CrossRef]
- Robles-Aguilar, A.A.; Pang, J.; Postma, J.A.; Schrey, S.D.; Lambers, H.; Jablonowski, N.D. The effect of pH on morphological and physiological root traits of Lupinus angustifolius treated with struvite as a recycled phosphorus source. Plant Soil 2019, 434, 65–78. [Google Scholar] [CrossRef] [Green Version]
- Camargo, C.E.D.O.; Ferreira Filho, A.W.P.; Salomon, M.V. Temperature and pH of the nutrient solution on wheat primary root growth. Sci. Agric. 2004, 61, 313–318. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tissue | Treatment | EcoFAB | Hydroponics | Sand | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| P Conc. | Total P Uptake | N Conc. | Total N Uptake | P Conc. | Total P Uptake | N Conc. | Total N Uptake | P Conc. | Total P Uptake | N Conc. | Total N Uptake | ||
| Shoot | High P | 3.62 | 0.09 | 41.9 | 0.99 | nd | nd | nd | nd | nd | nd | nd | nd |
| Algae | 2.63 | 0.04 | 23.6 | 0.35 | nd | nd | nd | nd | nd | nd | nd | nd | |
| Low P | 0.77 | 0.01 | 35.0 | 0.62 | nd | nd | nd | nd | nd | nd | nd | nd | |
| Root | High P | 3.61 | 0.02 | 29.9 | 0.20 | nd | nd | nd | nd | nd | nd | nd | nd |
| Algae | 3.72 | 0.03 | 20.2 | 0.17 | nd | nd | nd | nd | nd | nd | nd | nd | |
| Low P | 0.70 | 0.01 | 30.8 | 0.23 | nd | nd | nd | nd | nd | nd | nd | nd | |
| Whole | High P | 3.62 | 0.11 | 39.28 | 1.18 | 3.98 | 0.05 | 50.6 | 0.61 | 1.78 | 0.02 | 26.5 | 0.29 |
| plant | Algae | 3.03 | 0.07 | 22.35 | 0.52 | 5.87 | 0.06 | 56.8 | 0.54 | 1.35 | 0.01 | 23.7 | 0.25 |
| Low P | 0.75 | 0.02 | 33.74 | 0.85 | 5.96 | 0.08 | 51.0 | 0.68 | 0.90 | 0.01 | 22.0 | 0.20 | |
| Tissue | Treatment | EcoFAB | Hydroponic | Sand | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| P Conc. | Total P Uptake | N Conc. | Total N Uptake | P Conc. | Total P Uptake | N Conc. | Total N Uptake | P Conc. | Total P Uptake | N Conc. | Total N Uptake | ||
| Shoot | High P | 5.07 | 0.28 | 44.20 | 2.46 | 9.69 a ± 0.14 | 1.58a ± 0.11 | 63.50 a ± 0.61 | 10.33 a ± 0.75 | 3.06 A ± 0.21 | 0.08 A ± 0.01 | 34.37 A ± 1.17 | 0.96 A ± 0.08 |
| Algae | 4.12 | 0.21 | 24.70 | 1.28 | 8.27 b ± 0.15 | 0.84 b ± 0.11 | 59.40 b ± 0.86 | 6.04 b ± 0.71 | 2.12 A ± 0.41 | 0.06 A ± 0.01 | 27.37 B ± 1.34 | 0.72 A ± 0.05 | |
| Low P | 2.20 | 0.12 | 47.50 | 2.52 | 2.76 c ± 0.38 | 0.30 c ± 0.03 | 64.23 a ± 0.64 | 7.20 b ± 0.48 | 2.16 A ± 0.07 | 0.06 A ± 0.00 | 32.2 A,B ± 1.11 | 0.88 A ± 0.06 | |
| Root | High P | 2.27 | 0.04 | 18.90 | 0.32 | 4.32 a ± 0.43 | 0.09 a ± 0.00 | 28.07 a ± 1.75 | 0.58 a ± 0.01 | 1.36 A ± 0.21 | 0.05 A ± 0.01 | 12.53 A ± 0.41 | 0.42 A ± 0.03 |
| Algae | 1.65 | 0.06 | 11.10 | 0.38 | 3.68 a ± 0.38 | 0.06 a ± 0.01 | 36.10 b ± 0.47 | 0.63 a ± 0.05 | 1.08 A ± 0.15 | 0.04 A ± 0.01 | 9.47 B ± 0.24 | 0.39 A ± 0.04 | |
| Low P | 0.65 | 0.02 | 16.20 | 0.41 | 1.05 b ± 0.11 | 0.03 b ± 0.01 | 20.73 c ± 1.29 | 0.65 a ± 0.08 | 1.12 A ± 0.26 | 0.04 A ± 0.01 | 10.23 B ± 0.34 | 0.40 A ± 0.02 | |
| Whole plant | High P | 4.42 | 0.32 | 38.33 | 2.78 | 9.08 a ± 0.14 | 1.67 a ± 0.10 | 59.45 a ± 0.55 | 10.92 a ± 0.10 | 2.12 A ± 0.03 | 0.13 A ± 0.01 | 29.83 A ± 1.05 | 1.84 A ± 0.01 |
| Algae | 3.13 | 0.27 | 19.24 | 1.66 | 7.60 b ± 0.12 | 0.91 b ± 0.11 | 55.94 b ± 0.54 | 6.67 b ± 0.11 | 1.48 A ± 0.22 | 0.10 A ± 0.02 | 17.18 B ± 1.08 | 1.15 B ± 0.02 | |
| Low P | 1.70 | 0.13 | 37.45 | 2.93 | 2.39 c ± 0.29 | 0.34 c ± 0.02 | 54.75 b ± 0.59 | 7.85 b ± 0.02 | 1.54 A ± 0.15 | 0.10 A ± 0.01 | 24.27 C ± 0.99 | 1.60 A ± 0.01 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mau, L.; Junker, S.; Bochmann, H.; Mihiret, Y.E.; Kelm, J.M.; Schrey, S.D.; Roessner, U.; Schaaf, G.; Watt, M.; Kant, J.; et al. Root Growth and Architecture of Wheat and Brachypodium Vary in Response to Algal Fertilizer in Soil and Solution. Agronomy 2022, 12, 285. https://doi.org/10.3390/agronomy12020285
Mau L, Junker S, Bochmann H, Mihiret YE, Kelm JM, Schrey SD, Roessner U, Schaaf G, Watt M, Kant J, et al. Root Growth and Architecture of Wheat and Brachypodium Vary in Response to Algal Fertilizer in Soil and Solution. Agronomy. 2022; 12(2):285. https://doi.org/10.3390/agronomy12020285
Chicago/Turabian StyleMau, Lisa, Simone Junker, Helena Bochmann, Yeshambel E. Mihiret, Jana M. Kelm, Silvia D. Schrey, Ute Roessner, Gabriel Schaaf, Michelle Watt, Josefine Kant, and et al. 2022. "Root Growth and Architecture of Wheat and Brachypodium Vary in Response to Algal Fertilizer in Soil and Solution" Agronomy 12, no. 2: 285. https://doi.org/10.3390/agronomy12020285