Decomposition Rate of Organic Residues and Soil Organisms’ Abundance in a Subtropical Pyrus pyrifolia Field


 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
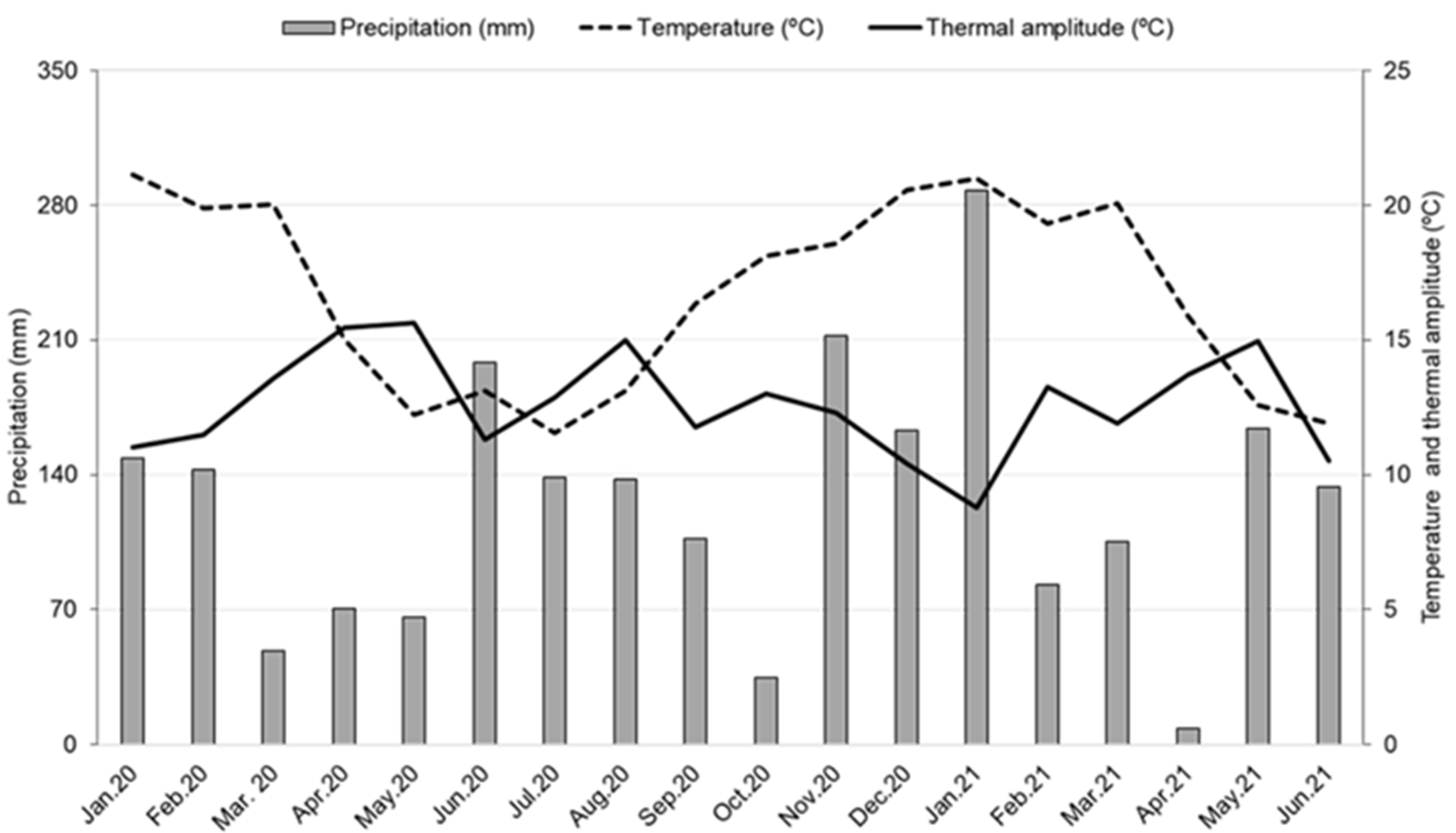
2.1. Pyrus Pyrifolia and Study Site
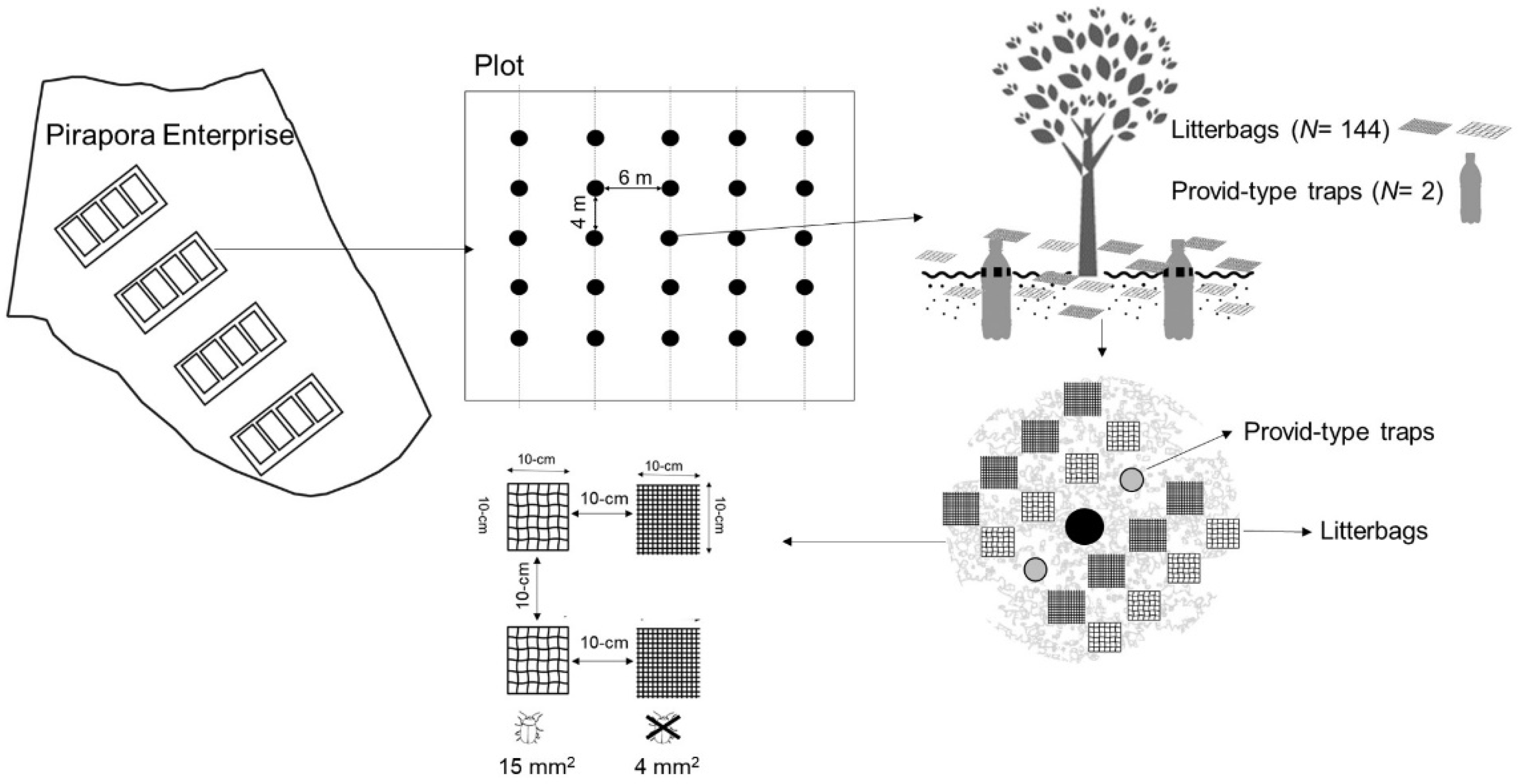
2.2. Experimental Design
2.3. Mulching and Compost Production
2.4. Soil Chemical Characterization
2.5. Organic Residues Decomposition Assay
2.6. Soil Organisms’ Collection
2.7. Statistical Analysis
3. Results
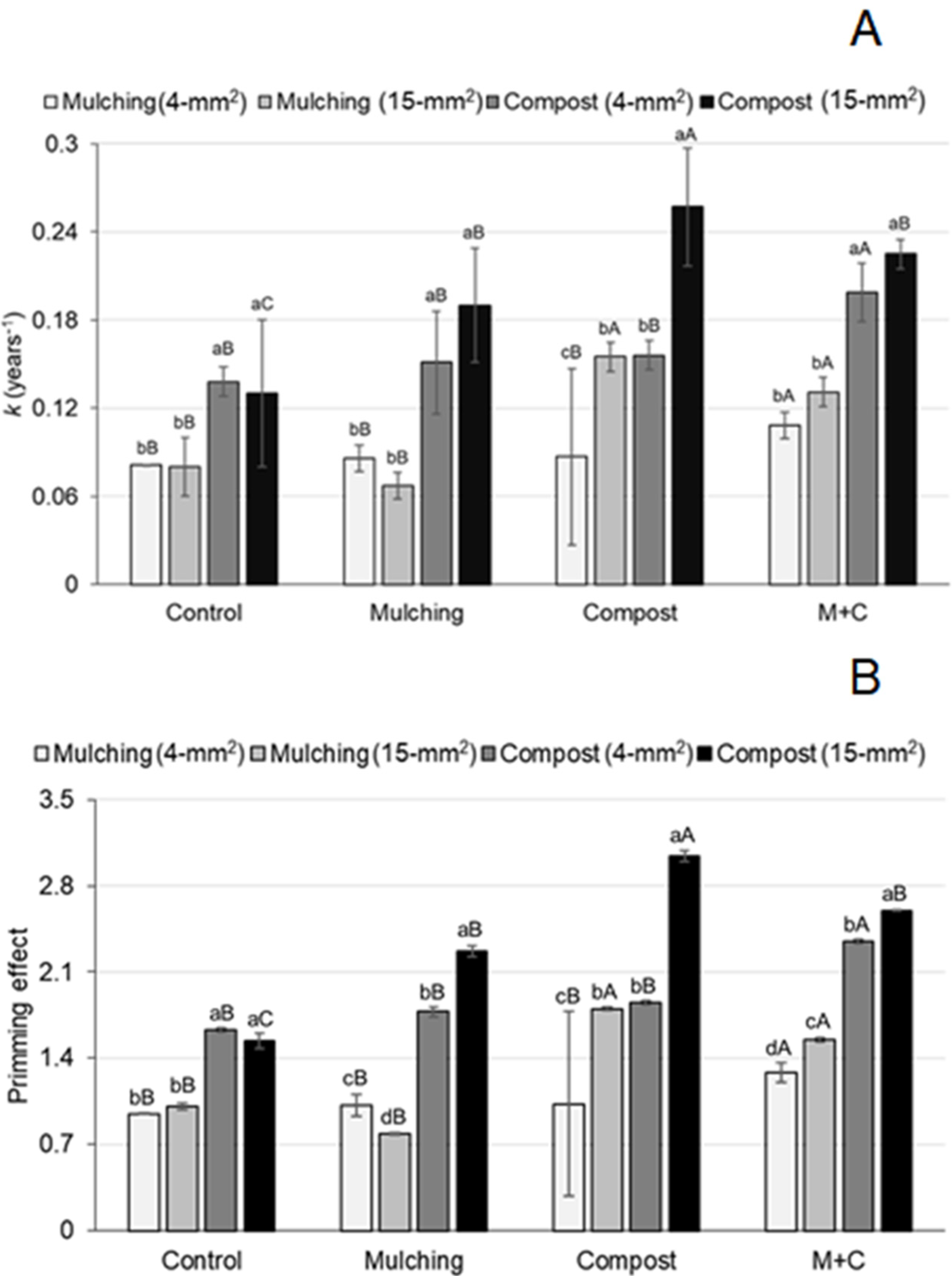
3.1. Influence of the Organic Residue Management and Soil Organisms’ Activity on Organic Residues Decomposition
3.2. Soil Organisms’ Collection in a 16-Year P. pyrifolia Field under Different Organic Residue Management
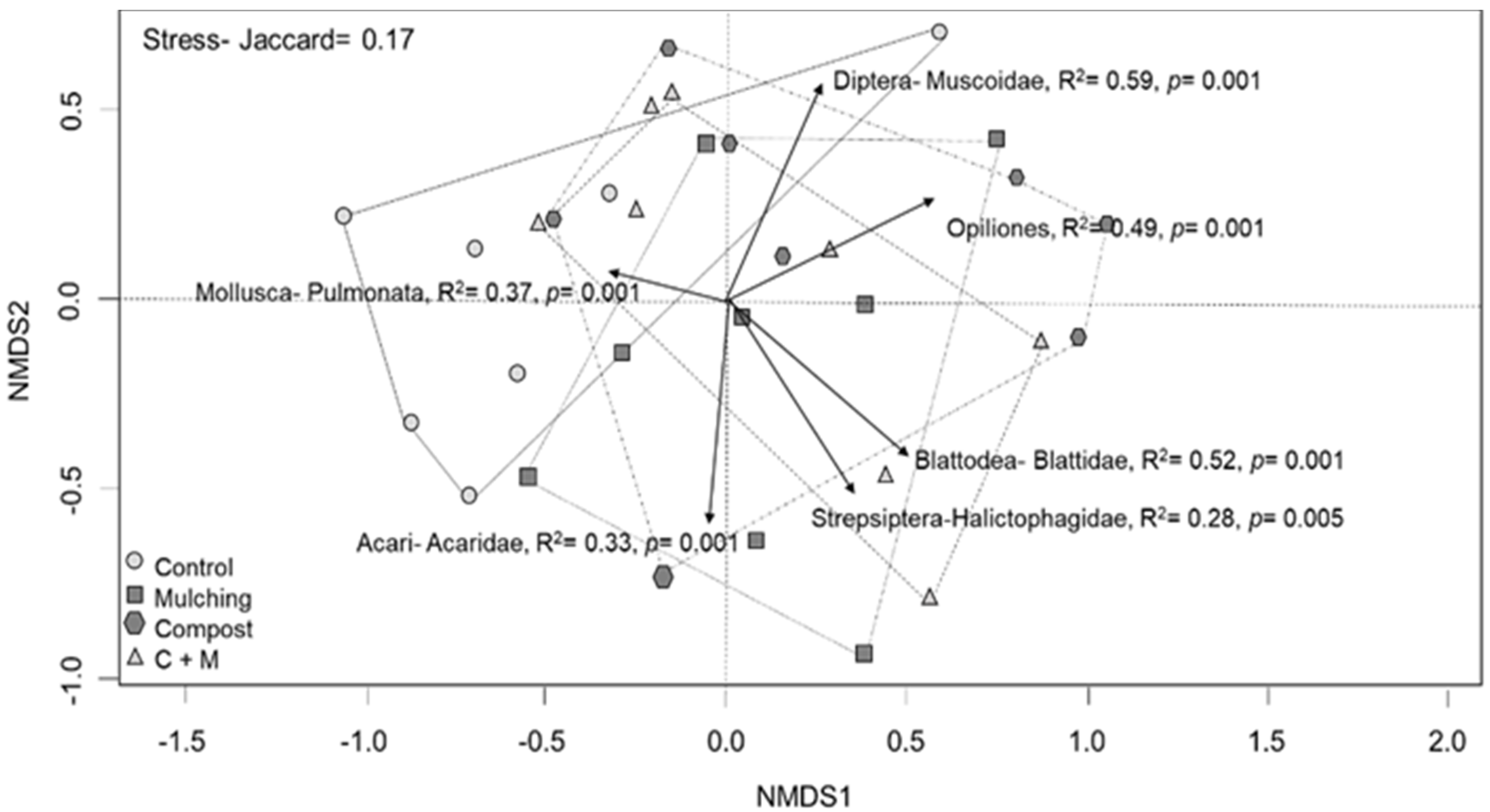
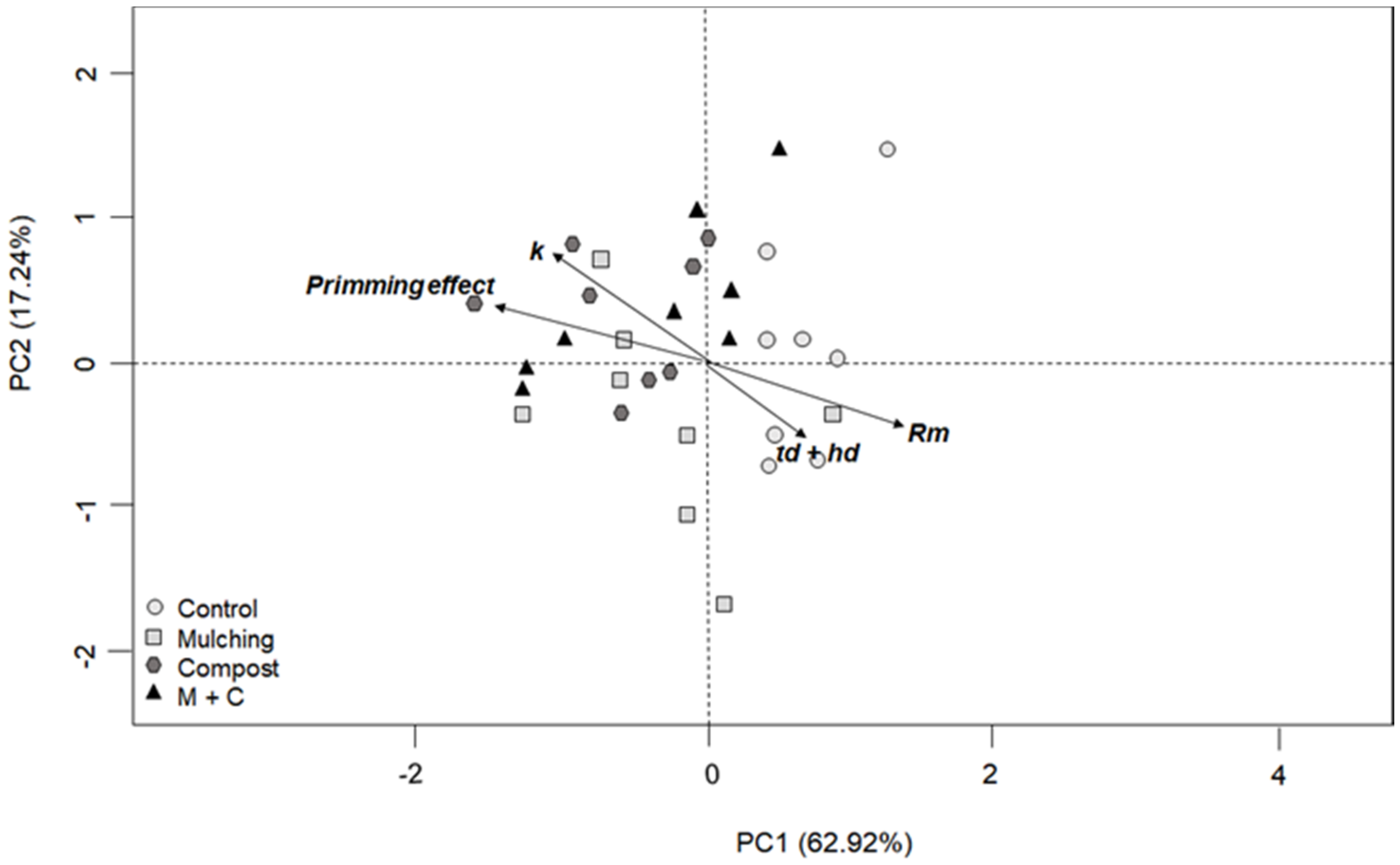
3.3. Multivariate Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chavarria, D.N.; Pérez-Brandan, C.; Serri, D.L.; Meriles, J.M.; Restovich, S.B.; Andiulo, A.E.; Jacquelin, L.; Vargas-Gil, S. Response of soil microbial communities to agroecological versus conventional systems of extensive agriculture. Agric. Ecosyst. Environ. 2018, 264, 1–8. [Google Scholar] [CrossRef]
- Da Silva, L.J.R.; Kormann, S.; Laurindo, L.K.; Barbosa, L.S.; Souza, T.A.F. O agronegócio da pera asiática no Sul do Brasil. In O Agronegócio da Pera Asiática no Sul do Brasil, 1st ed.; da Silva, L.J.R., Souza, T.A.F., Eds.; UFSC: Curitibanos, Brazil, 2021; Volume 1, pp. 1–24. [Google Scholar]
- Massaccesi, L.; Rondoni, G.; Tosti, G.; Conti, E.; Guiducci, M.; Agnelli, A. Soil functions are affected by transition from conventional to organic mulch-based cropping system. Appl. Soil Ecol. 2020, 153, 103639. [Google Scholar] [CrossRef]
- Wan, N.F.; Ji, X.Y.; Kiær, L.P.; Liu, S.; Deng, J.; Jiang, J.; Li, B. Ground cover increases spatial aggregation and association of insect herbivores and their predators in an agricultural landscape. Landsc. Ecol. 2018, 33, 799–809. [Google Scholar] [CrossRef]
- Cen, Y.; Li, L.; Guo, L.; Li, C.; Jiang, G. Organic management enhances both ecological and economic profitability of apple orchard: A case study in Shandong Peninsula. Sci. Hortic. 2020, 265, 109201. [Google Scholar] [CrossRef]
- De Leijster, V.; Verburg, R.W.; Santos, M.J.; Wassen, M.J.; Martínez-Mena, M.; de Vente, J.; Verweij, P.A. Almond farm profitability under agroecological management in south-eastern Spain: Accounting for externalities and opportunity costs. Agric. Syst. 2020, 183, 102878. [Google Scholar] [CrossRef]
- Zipori, I.; Dag, A.; Laor, Y.; Levy, G.L.; Einzenberg, H.; Yermiyahu, U.; Medina, S.; Saadi, I.; Krasnovski, A.; Raviv, M. Potential nutritional value of olive-mill wastewater applied to irrigated olive (Olea europaea L.) orchard in a semi-arid environment over 5 years. Sci. Hortic. 2018, 241, 218–224. [Google Scholar] [CrossRef]
- Barreto, C.F.; Antunes, L.E.C.; Ferreira, L.V.; Navroski, R.; Benati, J.A.; Nava, G. Nitrogen fertilization and genotypes of peaches in high-density. Rev. Bras. Frutic. 2020, 42, e-629. [Google Scholar] [CrossRef]
- Duan, S.; Iwanowicz, L.R.; Noguera-Oviedo, K.; Kaushal, S.S.; Rosenfeldt, E.J.; Aga, D.S.; Murthy, S. Evidence that watershed nutrient management practices effectively reduce estrogens in environmental waters. Sci. Total Environ. 2021, 758, 143904. [Google Scholar] [CrossRef] [PubMed]
- Melo, L.N.; Souza, T.A.F.; Santos, D. Cover crop farming system affect macroarthropods community diversity of Caatinga Brazil. Biologia 2019, 74, 1653–1660. [Google Scholar] [CrossRef]
- Sofo, A.; Mininni, A.N.; Ricciuti, P. Comparing the effects of soil fauna on litter decomposition and organic matter turnover in sustainably and conventionally managed olive orchards. Geoderma 2020, 372, 114393. [Google Scholar] [CrossRef]
- Coulis, M. Abundance, biomass, and community composition of soil saprophagous macrofauna in conventional and organic sugarcane fields. Soil Appl. Ecol. 2021, 164, 103923. [Google Scholar] [CrossRef]
- Wu, L.; Jiang, Y.; Zhao, F.; He, X.; Liu, H.; Yu, K. Increased organic fertilizer application and reduced chemical fertilizer application affect the soil properties and bacterial communities of grape rhizosphere soil. Sci. Rep. 2020, 10, 9568. [Google Scholar] [CrossRef] [PubMed]
- Orpet, R.J.; Jones, V.P.; Beers, E.H.; Reganold, J.P.; Goldberger, J.R.; Crowder, D.W. Perceptions and outcomes of conventional vs. organic apple orchard management. Agric. Ecosyst. Environ. 2020, 289, 106723. [Google Scholar] [CrossRef]
- Rieff, G.G.; Natal-da-Luz, T.; Renaud, M.; Azevedo-Pereira, H.M.V.S.; Chichorro, F.; Schmelz, R.M.; de Sá, E.L.S.; Sousa, J.P. Impact of no-tillage versus conventional maize plantation on soil mesofauna with and without the use of a lambda-cyhalothrin based insecticide: A terrestrial model ecosystem experiment. Appl. Soil Ecol. 2020, 147, 103381. [Google Scholar] [CrossRef]
- Libutti, A.; Cammerino, A.R.B.; Monteleone, M. Management of Residues from Fruit Tree Pruning: A Trade-Off between Soil Quality and Energy Use. Agronomy 2021, 11, 236. [Google Scholar] [CrossRef]
- Forstall-Sosa, K.S.; Souza, T.A.F.; Lucena, E.O.; da Silva, S.A.I.; Ferreira, J.T.A.; Silva, T.N.; Santos, D.; Niemeyer, J.C. Soil macroarthropod community and soil biological quality index in a green manure farming system of the Brazilian semi-arid. Biologia 2020, 76, 907–917. [Google Scholar] [CrossRef]
- Jacobsen, S.K.; Moraes, G.J.; Sørensen, H.; Sigsgaard, L. Organic cropping practice decreases pest abundance and positively influences predator-prey interactions. Agric. Ecosyst. Environ. 2019, 272, 1–9. [Google Scholar] [CrossRef]
- Li, F.; Sørensen, P.; Li, X.; Olesen, J.E. Carbon and nitrogen mineralization differ between incorporated shoots and roots of legume versus non-legume-based cover crops. Plant Soil 2020, 446, 243–257. [Google Scholar] [CrossRef]
- Kai, T.; Adhikari, D. Effect of Organic and Chemical Fertilizer Application on Apple Nutrient Content and Orchard Soil Condition. Agriculture 2021, 11, 340. [Google Scholar] [CrossRef]
- Popov, V.; Kostadinova, E.; Rancheva, E.; Yancheva, C. Causal relationship between biodiversity of insect population and agro-management in organic and conventional apple orchard. Org. Agr. 2018, 8, 355–370. [Google Scholar] [CrossRef]
- Yang, B.; Banerjee, S.; Herzong, C.; Ramírez, C.; Dahlin, P.; van der Heijden, G.A. Impact of land use type and organic farming on the abundance, diversity, community composition and functional properties of soil nematode communities in vegetable farming. Agric. Ecosyst. Environ. 2021, 318, 107488. [Google Scholar] [CrossRef]
- Souza, T.A.F.; Freitas, H. Long-term effects of fertilization on soil organism diversity. In Sustainable Agriculture Reviews; Gaba, S., Smith, B., Lichtfouse, E., Eds.; Springer: Cham, Switzerland, 2018; pp. 211–247. [Google Scholar] [CrossRef]
- Zhang, K.; Maltais-Landy, G.; Liao, H. How soil biota regulate C cycling and soil C pools in diversified crop rotations. Soil Biol. Biochem. 2021, 156, 108219. [Google Scholar] [CrossRef]
- Araújo, M.D.M.; Feitosa, M.M.; Primo, A.A.; Taniguchi, C.A.K.; Souza HA, D. Mineralization of nitrogen and carbon from organic compost from animal production waste. Rev. Caatinga 2020, 33, 310–320. [Google Scholar] [CrossRef]
- Liu, M.; Qiao, N.; Xu, X.; Fang, H.; Wang, H.; Kuzyakov, Y. C: N stoichiometry of stable and labile organic compounds determine priming patterns. Geoderma 2020, 362, 114122. [Google Scholar] [CrossRef]
- Anjum; Khan, A. Decomposition of soil organic matter is modulated by soil amendments. Carbon Manag. 2021, 12, 37–50. [Google Scholar] [CrossRef]
- Tian, K.; Kong, X.; Yuan, L.; Lin, H.; He, Z.; Yao, B.; Ji, Y.; Yang, J.; Sun, S.; Tian, X. Priming effect of litter mineralization: The role of root exudate depends on its interactions with litter quality and soil condition. Plant Soil 2019, 440, 457–471. [Google Scholar] [CrossRef]
- Baldi, E.; Gioacchini, P.; Montecchio, D.; Mocali, S.; Antonielli, L.; Masoero, G.; Toselli, M. Effect of biofertilizers application on soil biodiversity and litter degradation in a commercial apricot orchard. Agronomy 2021, 11, 1116. [Google Scholar] [CrossRef]
- Kan, Z.; Virk, A.L.; Wu, G.; Qi, J.; Ma, S.; Wang, X.; Zhao, X.; Lal, R.; Zhang, H. Priming effect intensity of soil organic carbon mineralization under no-till and residue retention. Appl. Soil Ecol. 2020, 147, 103445. [Google Scholar] [CrossRef]
- FAOSTAT—Food and Agriculture Organization of the United Nations Statistics. Production/Yield Quantities of Pears in Brazil. 2020. Available online: http://www.fao.org/faostat/en/#data/QC/visualize (accessed on 23 August 2021).
- IUSS Working Group WRB. World Reference Base for Soil Resources 2014, update 2015: International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; World Soil Resources Reports, 106; FAO: Rome, Italy, 2015. [Google Scholar]
- Laurindo, L.K.; Souza, T.A.F.; Silva, L.J.R.; Kormann, S.; Lucena, E.O. Propriedades químicas do solo. In Indicadores da Qualidade do Solo em Sistemas Agroflorestais e Ecossistemas Associados, 1st ed.; Laurindo, L.K., Souza, T.A.F., Eds.; PPGEAN: Curitibanos, Brazil, 2020; pp. 61–75. [Google Scholar]
- Tedesco, M.J.; Gianello, C.; Bissani, C.A.; Bohnen, H.; Volkweiss, S.J. Análise do Solo, Planta e Outros Materiais, 2nd ed.; UFRGS: Porto Alegre, Brazil, 1995; 174p. [Google Scholar]
- Teixeira, P.C.; Donagemma, G.K.; Fontana, A.; Teixeira, W.G. Manual de Métodos de Análise Do Solo; Embrapa Solos: Brasília, Brasil, 2017; 212p. [Google Scholar]
- Nascimento, G.S.; Souza, T.A.F.; da Silva, L.J.R.; Santos, D. Soil physico-chemical properties, biomass production, and root density in a green manure farming system from tropical ecosystem, North-eastern Brazil. J. Soils Sediments 2021, 21, 2203–2211. [Google Scholar] [CrossRef]
- Olson, J.S. Energy storage and the balance of producers and decomposers in ecological systems. Ecology 1963, 44, 322–331. [Google Scholar] [CrossRef] [Green Version]
- Tan, B.; Yin, R.; Zhang, J.; Xu, Z.; Liu, Y.; He, S.; Zhang, L.; Li, H.; Wang, L.; Liu, S.; et al. Temperature and Moisture Modulate the Contribution of Soil Fauna to Litter Decomposition via Different Pathways. Ecosystems 2021, 24, 1142–1156. [Google Scholar] [CrossRef]
- Anderson, J.N.; Ingram, J.S.I. Tropical Solo Biology and Fertility: A Handbook of Methods; CAB International: Wallingford, UK, 1993. [Google Scholar] [CrossRef]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Champaign, IL, USA, 1949. [Google Scholar] [CrossRef] [Green Version]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org/ (accessed on 23 August 2021).
- Asigbaase, M.; Dawoe, E.; Sjogersten, S.; Lomax, B.H. Decomposition and nutrient mineralisation of leaf litter in smallholder cocoa agroforests: A comparison of organic and conventional farms in Ghana. J. Soils Sediments 2021, 21, 1010–1023. [Google Scholar] [CrossRef]
- Tassinari, A.; da Silva, L.O.S.; Drescher, G.L.; de Oliveira, R.A.; Baldi, E.; de Melo, G.W.B.; Zalamena, J.; Mayer, N.A.; Giacomini, S.J.; Carranca, C.L.A.F.; et al. Contribution of Cover Crop Residue Decomposition to Peach Tree Nitrogen Nutrition. J. Soil Sci. Plant Nutr. 2021, 21, 2124–2136. [Google Scholar] [CrossRef]
- Gonçalves, F.; Nunes, C.; Carlos, C.; López, A.; Oliveira, I.; Crespi, A.; Teixeira, B.; Pinto, R.; Costa, C.A.; Torres, L. Do soil management practices affect the activity density, diversity, and stability of soil arthropods in vineyards? Agric. Ecosyst. Environ. 2020, 294, 106863. [Google Scholar] [CrossRef]
- Kitamura, A.E.; Tavares, R.L.M.; Alves, M.C.; de Souza, Z.M.; Siqueira, D.S. Soil macrofauna as bioindicator of the recovery of degraded Cerrado soil. Soil Sci. 2020, 50, e20190606. [Google Scholar] [CrossRef]
- Geldenhuys, M.; Gaigher, R.; Pryke, J.S.; Samways, M.J. Diverse herbaceous cover crops promote vineyard arthropod diversity across different management regimes. Agric. Ecosyst. Environ. 2021, 307, 107222. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, B.; An, S. Ecological stoichiometry in leaves, roots, litters and soil among different plant communities in a desertified region of Northern China. Catena 2018, 166, 238–338. [Google Scholar] [CrossRef]
- Maran, A.M.; Weintraub, M.N.; Pelini, S.L. Does stimulating ground arthropods enhance nutrient cycling in conventionally managed corn fields? Agric. Ecosyst. Environ. 2020, 297, 106934. [Google Scholar] [CrossRef]
- Almagro, M.; Ruiz-Navaro, A.; Diaz-Pereira, E.; Albaladejo, J.; Martínez-Mena, M. Plant residue chemical quality modulates the soil microbial response related to decomposition and soil organic carbon and nitrogen stabilization in a rainfed Mediterranean agroecosystem. Soil Biol. Biochem. 2021, 156, 108198. [Google Scholar] [CrossRef]
- Long, J.; Zhang, M.; Li, J.; Liao, H.; Wang, X. Soil macro- and mesofauna-mediated litter decomposition in a subtropical karst forest. Biotropica 2021, 53, 1465–1474. [Google Scholar] [CrossRef]
- Shang, L.; Wan, L.; Zhou, X.; Li, S.; Li, X. Effects of organic fertilizer on soil nutrient status, enzyme activity, and bacterial community diversity in Leymus chinensis steppe in Inner Mongolia, China. PLoS ONE 2020, 15, e0240559. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.; Savin, M.C.; Rom, C.R.; Gbur, E. Soil microbial and nutrient responses over seven years of organic apple orchard maturation. Nutr. Cycl. Agroecosyst. 2020, 118, 23–38. [Google Scholar] [CrossRef]
- Thakur, M.; Kumar, R. Mulching: Boosting crop productivity and improving soil environment in herbal plants. J. Appl. Res. Med. Aromat. Plants 2020, 20, 100287. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, L.; He, R.; Chen, Y.; Xu, Z.; Tan, B.; Zhang, L.; Xiao, J.; Zhu, P.; Chen, L.; et al. Higher soil fauna abundance accelerates litter carbon release across an alpine forest-tundra ecotone. Sci. Rep. 2019, 9, 10561. [Google Scholar] [CrossRef]
- Liu, S.; Behm, J.E.; Wan, S.; Yan, J.; Ye, Q.; Zhang, W.; Yang, X.; Fu, S. Effects of canopy nitrogen addition on soil fauna and litter decomposition rate in a temperate forest and a subtropical forest. Geoderma 2021, 389, 114703. [Google Scholar] [CrossRef]
- Plaas, E.; Meyer-Wolfarth, F.; Banse, M.; Bengtsson, J.; Bergmann, H.; Faber, J.; Potthoff, M.; Runge, T.; Schrader, S.; Taylor, A. Towards valuation of biodiversity in agricultural soils: A case for earthworms. Ecol. Econ. 2019, 159, 291–300. [Google Scholar] [CrossRef]
- Mockeviciene, I.; Repsiene, R.; Amaleviciute-Volunge, K.; Karcauskiene, D.; Slepetiene, A.; Lepane, V. Effect of long-term application of organic fertilizers on improving organic matter quality in acid soil. Arch. Agron. Soil Sci. 2021, 195, e104382. [Google Scholar] [CrossRef]
- Frouz, J. Effects of soil macro- and mesofauna on litter decomposition and soil organic matter stabilization. Geoderma 2018, 332, 161–172. [Google Scholar] [CrossRef]
- Sharma, S.; Singh, P.; Choudhary, O.P.; Neemisha. Nitrogen and rice straw incorporation impact nitrogen use efficiency, soil nitrogen pools and enzyme activity in rice-wheat system in north-western India. Field Crops Res. 2021, 266, 108131. [Google Scholar] [CrossRef]
- Mariotte, P.; Mehrabi, Z.; Bezemer, T.M.; de Deyn, G.B.; Kulmastiski, A.; Drigo, B.; Veen, C.; Van der Heijden, M.G.A.; Kardol, P. Plant–Soil Feedback. Bridg. Nat. Agric. Sci. 2018, 33, 129–142. [Google Scholar] [CrossRef] [Green Version]
- Vignozzi, N.; Angelli, A.E.; Brandi, G.; Gagnarli, E.; Goggiolo, D.; Lagomarsino, A.; Pellegrini, S.; Simoncini, S.; Valboa, G.; Caruso, G.; et al. Soil ecosystem functions in a high-density olive orchard managed by different soil conservation practices. Appl. Soil Ecol. 2019, 134, 64–79. [Google Scholar] [CrossRef]
- De Pedro, L.; Perera-Fernández, L.G.; López-Gallego, E.; Pérez-Marcos, M.; Sanchez, J.Á. The effect of cover crops on the biodiversity and abundance of ground-dwelling arthropods in a mediterranean pear orchard. Agronomy 2020, 10, 580. [Google Scholar] [CrossRef] [Green Version]
- Gómez, J.A.; Campos, M.; Guzmán, G.; Castillo-Llanque, F.; Vanwalleghem, T.; Lora, A.; Giráldez, J.V. Soil erosion control, plant diversity, and arthropod communities under heterogeneous cover crops in an olive orchard. Environ. Pollut. Res. 2018, 25, 977–989. [Google Scholar] [CrossRef] [PubMed]
- Bufebo, B.; Elias, E.; Getu, E. Abundance and diversity of soil invertebrate macro-fauna in different land uses at Shenkolla watershed, South Central Ethiopia. J. Basic Appl. Zool. 2021, 82, 11. [Google Scholar] [CrossRef]
- Simoni, S.; Caruso, G.; Vignozzi, N.; Gucci, R.; Valboa, G.; Pellegrini, S.; Palai, G.; Goggioli, D.; Gagnarli, E. Effect of Long-Term Soil Management Practices on Tree Growth, Yield and Soil Biodiversity in a High-Density Olive Agro-Ecosystem. Agronomy 2021, 11, 1036. [Google Scholar] [CrossRef]
- Li, S.; Song, M.; Jing, S. Effects of different carbon inputs on soil nematode abundance and community composition. Appl. Soil Ecol. 2021, 163, 103915. [Google Scholar] [CrossRef]
- Galloway, A.D.; Seymour, C.L.; Gaigher, R.; Pryke, J.S. Organic farming promotes arthropod predators, but this depends on neighbouring patches of natural vegetation. Agric. Ecosyst. Environ. 2021, 310, 107295. [Google Scholar] [CrossRef]
- Mabin, M.D.; Welty, C.; Gardiner, M.M. Predator richness predicts pest suppression within organic and conventional summer squash (Cucurbita pepo L. Cucurbitales: Cucurbitaceae). Agric. Ecosyst. Environ. 2020, 287, 106689. [Google Scholar] [CrossRef]
- Wang, M.; Yu, Z.; Liu, Y.; Wu, P.; Axmacher, J.C. Taxon- and functional group-specific responses of ground beetles and spiders to landscape complexity and management intensity in apple orchards of the North China Plain. Agric. Ecosyst. Environ. 2022, 323, 107700. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organic Residues | C/N Ratio | N (g kg−1) | P (g kg−1) | K (g kg−1) |
|---|---|---|---|---|
| Mulching | 45.85 ± 0.98 1 | 8.52 ± 1.12 | 13.87 ± 1.34 | 86.68 ± 4.23 |
| Compost | 21.13 ± 1.02 | 20.84 ± 1.18 | 16.18 ± 1.37 | 31.18 ± 4.39 |
| Treatments | pH (H2O) | P (mg dm−3) | K+ (mg dm−3) | Ca2+ (cmolc dm−3) | Mg2+ (cmolc dm−3) | SOC 1 (g kg−1) | TN 2 (g kg−1) |
|---|---|---|---|---|---|---|---|
| Control | 6.28 ± 0.03 | 30.22 ± 1.04 | 408.23 ± 2.32 | 10.28 ± 0.08 | 3.08 ± 0.02 | 30.59 ± 2.34 | 1.62 ± 0.08 |
| Mulching (M) | 6.35 ± 0.03 | 48.98 ± 1.29 | 461.30 ± 2.09 | 11.88 ± 0.08 | 3.06 ± 0.03 | 30.59 ± 1.99 | 1.81 ± 0.05 |
| Compost (C) | 6.15 ± 0.02 | 35.07 ± 1.27 | 326.31 ± 1.99 | 10.96 ± 0.05 | 3.26 ± 0.02 | 27.98 ± 2.11 | 1.80 ± 0.06 |
| M + C | 6.23 ± 0.04 | 43.12 ± 1.39 | 576.92 ± 2.05 | 10.36 ± 0.09 | 2.92 ± 0.03 | 30.16 ± 2.10 | 1.78 ± 0.02 |
| Organic Residues Management | Mesh—Type | |||
|---|---|---|---|---|
| hd (Days) | ||||
| Compost, 4 mm2 | Compost, 15 mm2 | Mulching, 4 mm2 | Mulching, 15 mm2 | |
| Control | 50.86 (0.59) cA 1 | 61.61 (2.64) bA | 86.03 (0.67) aB | 86.88 (2.56) aB |
| Mulching (M) | 47.88 (1.21) cB 2 | 37.28 (0.76) dB | 81.00 (0.90) bB | 105.21 (1.53) aA |
| Compost (C) | 44.72 (0.39) bB | 27.54 (0.45) cC | 132.25 (9.64) aA | 45.77 (0.53) bC |
| M + C | 35.12 (0.36) bC | 30.97 (0.21) bC | 64.43 (0.58) aC | 53.84 (0.78) aC |
| td (days) | ||||
| Control | 308.99 (9.44) bA | 374.33 (19.58) bA | 522.64 (15.25) aB | 527.83 (21.80) aB |
| Mulching (M) | 290.92 (10.76) cA | 226.48 (7.95) cB | 492.11 (14.93) bB | 637.79 (19.61) aA |
| Compost (C) | 274.05 (9.16) bB | 167.31 (5.48) cC | 806.88 (6.56) aA | 275.71 (7.23) bC |
| M + C | 213.19 (6.41) bC | 188.18 (5.44) cC | 391.42 (11.57) aC | 327.07 (10.40) aC |
| Rm (%) | ||||
| Control | 19.75 (0.31) cA | 24.85 (1.47) bA | 38.35 (0.16) aB | 37.59 (0.10) aB |
| Mulching (M) | 17.75 (0.68) cA | 11.05 (0.46) cB | 36.05 (0.34) bB | 45.42 (0.45) aA |
| Compost (C) | 15.87 (0.26) bA | 5.15 (0.22) cB | 43.42 (2.86) aA | 16.47 (0.20) bC |
| M + C | 9.60 (0.18) bB | 7.00 (0.04) bB | 27.80 (0.23) aC | 21.55 (0.43) aC |
| Order—Family | Control | Mulching (M) | Compost (C) | M + C | F-Value |
|---|---|---|---|---|---|
| Acari—Acaridae | 0.62 (0.11) b 1 | 1.75 (0.21) a | 0.12 (0.03) c | 0.50 (0.07) b | 10.62 *** 2 |
| Araneae—Araneidae | 2.25 (0.15) a | 1.50 (0.13) b | 0.87 (0.10) d | 1.12 (0.10) c | 8.25 ** 3 |
| Araneae—Filistatidae | 4.62 (0.26) a | 5.25 (0.38) a | 4.37 (0.19) a | 5.75 (0.22) a | 3.07 ns 4 |
| Blattodea—Blattidae | - | 0.50 (0.05) b | 0.37 (0.05) c | 0.62 (0.07) a | 11.83 *** |
| Blattodea—Termitidae | 0.37 (0.05) a | 0.12 (0.03) b | - | - | 13.50 *** |
| Coleoptera—Carabidae | 15.75 (1.49) a | 12.37 (0.98) a | 15.00 (0.99) a | 13.87 (1.76) a | 2.00 ns |
| Coleoptera—Cerambycidae | 0.12 (0.03) a | 0.12 (0.03) a | - | 0.12 (0.03) a | 2.17 ns |
| Coleoptera—Cuccilinidae | - | 0.12 (0.03) a | - | - | 6.09 ns |
| Coleoptera—Cugygidae | - | 0.12 (0.03) b | 0.25 (0.04) a | - | 7.96 ** |
| Coleoptera—Gyrinidae | - | 0.12 (0.03) a | 0.37 (0.07) a | 0.12 (0.03) a | 4.77 ns |
| Coleoptera—Nitidulidae | 34.37 (1.22) a | 32.50 (1.15) a | 33.75 (1.04) a | 44.87 (2.13) a | 4.41 ns |
| Coleoptera—Passalidae | 0.12 (0.03) a | - | - | - | 6.09 ns |
| Coleoptera—Scarabaeidae | 7.37 (0.58) a | 8.62 (0.92) a | 9.37 (0.93) a | 3.75 (0.25) a | 4.42 ns |
| Coleoptera—Staphylinidae | 2.12 (0.33) a | 1.12 (0.16) c | 0.25 (0.04) d | 1.87 (0.22) b | 10.07 *** |
| Dermaptera—Forficulidae | 1.87 (0.15) d | 3.50 (0.22) b | 7.50 (0.70) a | 2.50 (0.12) c | 9.96 *** |
| Diptera—Muscoidea | 1.12 (0.27) d | 2.37 (0.23) c | 3.00 (0.23) b | 3.37 (0.27) a | 10.51 *** |
| Gastropoda—Gymnomorpha | 0.62 (0.07) c | 1.50 (0.13) b | 1.62 (0.16) a | 1.75 (0.11) a | 8.75 ** |
| Gastropoda—Pulmonata | 1.37 (0.14) b | 2.37 (0.47) a | 0.12 (0.03) d | 1.87 (0.22) c | 10.87 *** |
| Haplotaxida—Lumbricidae | 0.62 (0.08) a | 1.12 (0.08) a | 1.00 (0.14) a | 0.37 (0.05) a | 7.11 ns |
| Hemiptera—Cicadidae | 0.37 (0.05) a | - | - | 0.12 (0.03) b | 13.50 *** |
| Hemiptera—Pentatomidae | 0.12 (0.03) a | - | - | - | 6.09 ns |
| Hymenoptera—Formicidae | 100.24 (7.65) a | 68.25 (3.35) a | 67.27 (4.57) a | 65.65 (5.63) a | 3.32 ns |
| Hymenoptera—Vespidae | 0.12 (0.03) a | 0.12 (0.03) a | - | 0.12 (0.03) a | 2.17 ns |
| Larvae of Lepidoptera | 5.87 (0.45) a | 3.12 (0.28) a | 8.00 (0.59) a | 8.25 (0.79) a | 6.94 ns |
| Lepidoptera | 0.12 (0.03) a | 0.12 (0.03) a | 0.12 (0.03) a | 0.25 (0.04) a | 1.40 ns |
| Mollusca—Pulmonata | 0.50 (0.05) a | 0.62 (0.07) a | 0.25 (0.04) a | 0.62 (0.07) a | 3.50 ns |
| Neuroptera—Chrysopidae | - | 0.25 (0.06) b | - | 0.37 (0.07) a | 7.77 ** |
| Orthoptera—Grylloidea | 0.12 (0.03) a | 0.12 (0.03) a | - | - | 4.20 ns |
| Opiliones | 0.12 (0.03) a | 0.12 (0.03) a | 0.25 (0.04) a | 0.12 (0.03) a | 1.40 ns |
| Scutigeromorpha—Scutigeridae | - | 0.12 (0.03) a | 0.12 (0.03) a | - | 4.20 ns |
| Spirobolida—Scolopendromorpha | 0.12 (0.03) a | - | - | - | 6.09 ns |
| Strepsiptera—Halictophagidae | - | 1.87 (0.20) a | 1.62 (0.25) b | 1.25 (0.17) c | 12.51 *** |
| Thysanoptera—Thripidae | 3.00 (0.92) a | 3.75 (0.38) a | 6.62 (0.67) a | (4.75 ± 0.46) a | 4.88 ns |
| Ecological indices | Control | Mulching (M) | Compost (C) | M + C | F-value |
| Richness—S | 17.75 (0.26) b | 19.87 (0.15) a | 17.50 (0.19) c | 19.37 (0.23) a | 11.28 *** |
| Shannon’s diversity index—H | 2.00 (0.05) a | 2.14 (0.03) a | 2.15 (0.04) a | 2.08 (0.04) a | 5.06 ns |
| Simpson’s dominance index—C | 0.81 (0.05) a | 0.83 (0.03) a | 0.84 (0.04) a | 0.82 (0.05) a | 5.58 ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodrigues da Silva, L.J.; Feitosa de Souza, T.A.; Klestadt Laurindo, L.; Freitas, H.; Costa Campos, M.C. Decomposition Rate of Organic Residues and Soil Organisms’ Abundance in a Subtropical Pyrus pyrifolia Field. Agronomy 2022, 12, 263. https://doi.org/10.3390/agronomy12020263
Rodrigues da Silva LJ, Feitosa de Souza TA, Klestadt Laurindo L, Freitas H, Costa Campos MC. Decomposition Rate of Organic Residues and Soil Organisms’ Abundance in a Subtropical Pyrus pyrifolia Field. Agronomy. 2022; 12(2):263. https://doi.org/10.3390/agronomy12020263
Chicago/Turabian StyleRodrigues da Silva, Lucas Jónatan, Tancredo Augusto Feitosa de Souza, Lídia Klestadt Laurindo, Helena Freitas, and Milton César Costa Campos. 2022. "Decomposition Rate of Organic Residues and Soil Organisms’ Abundance in a Subtropical Pyrus pyrifolia Field" Agronomy 12, no. 2: 263. https://doi.org/10.3390/agronomy12020263