
Investigating the Responses of Microbial Communities to Banana Fusarium Wilt in Suppressive and Conducive Soils Based on Soil Particle-Size Differentiation

Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil Collection and Preparation
2.2. Determination of Enzyme Activities
2.3. Soil DNA Extraction and Real-Time PCR
2.4. Illumina MiSeq High-Throughput Sequencing and Data Analysis
2.5. Statistical Analysis
3. Results
3.1. Soil Properties, Multiple Enzyme Activities and Particle-Size Distribution
3.2. The Gene Abundances for Bacteria, Fungi, and F. oxysporum
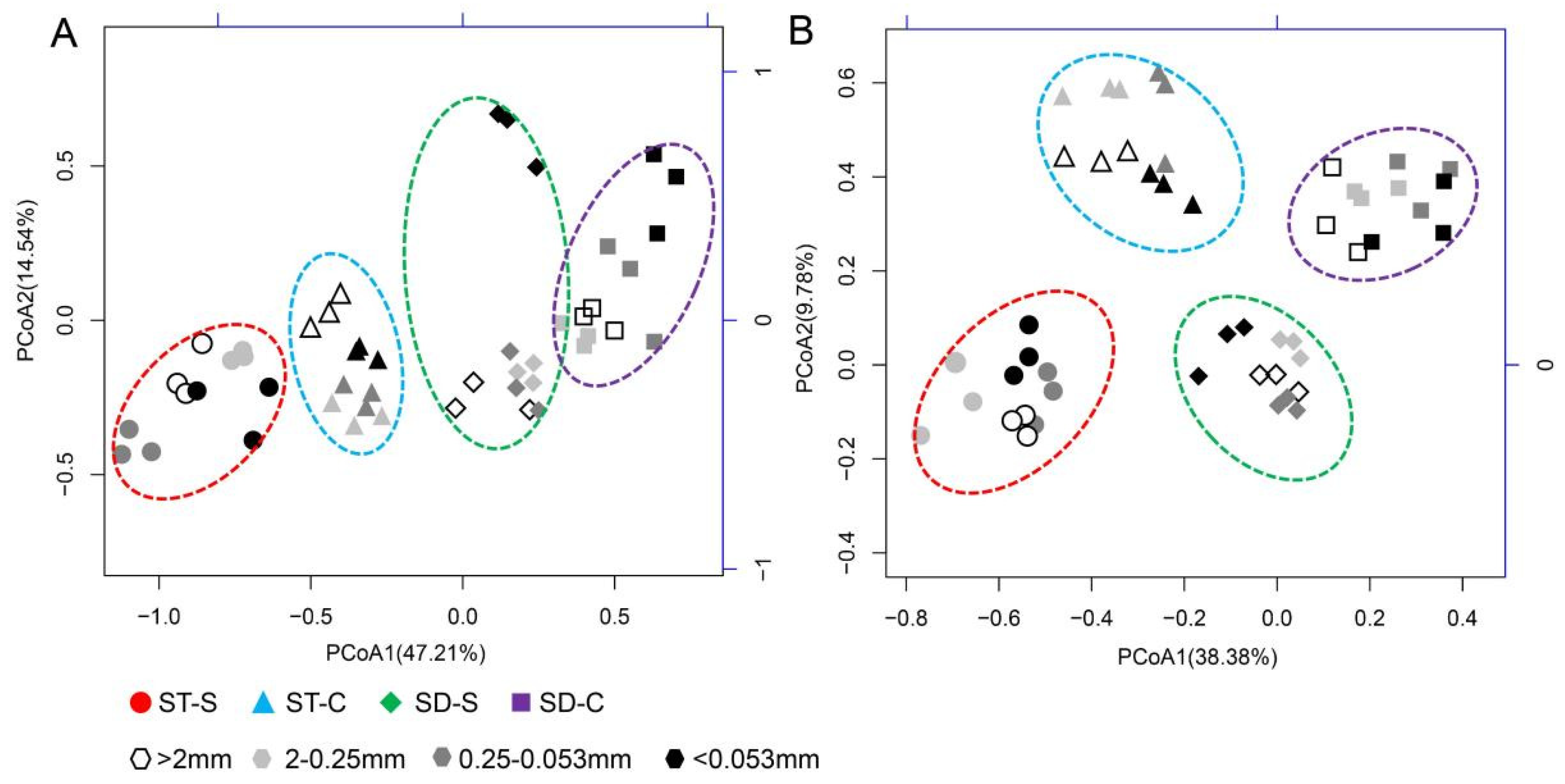
3.3. Community Structure of Bacteria and Fungi in The suppressive and Conducive Soils
3.4. The Taxonomic Composition of Bacteria and Fungi in the Suppressive and Conducive Soils
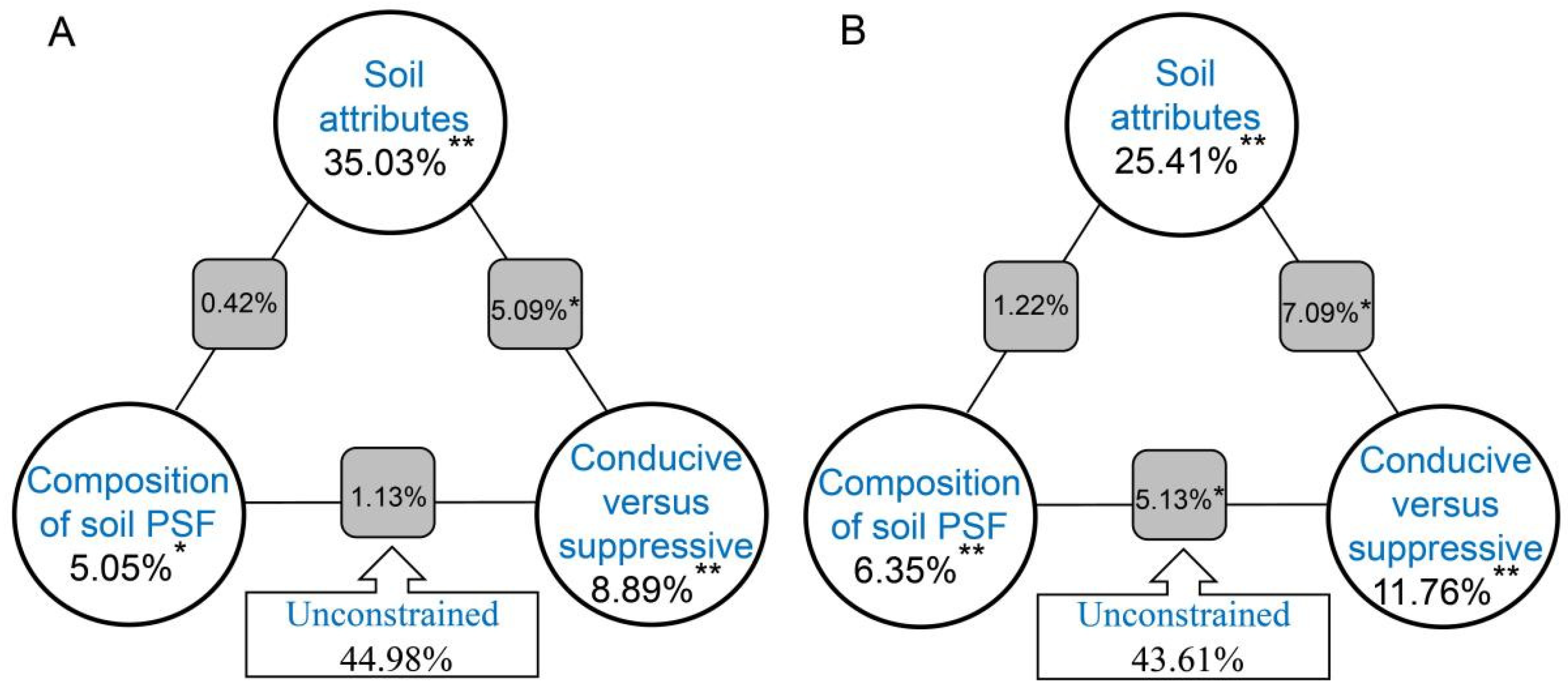
3.5. The Primary Factors Influencing the Microbial Community Structure in the Suppressive and Conducive Soils
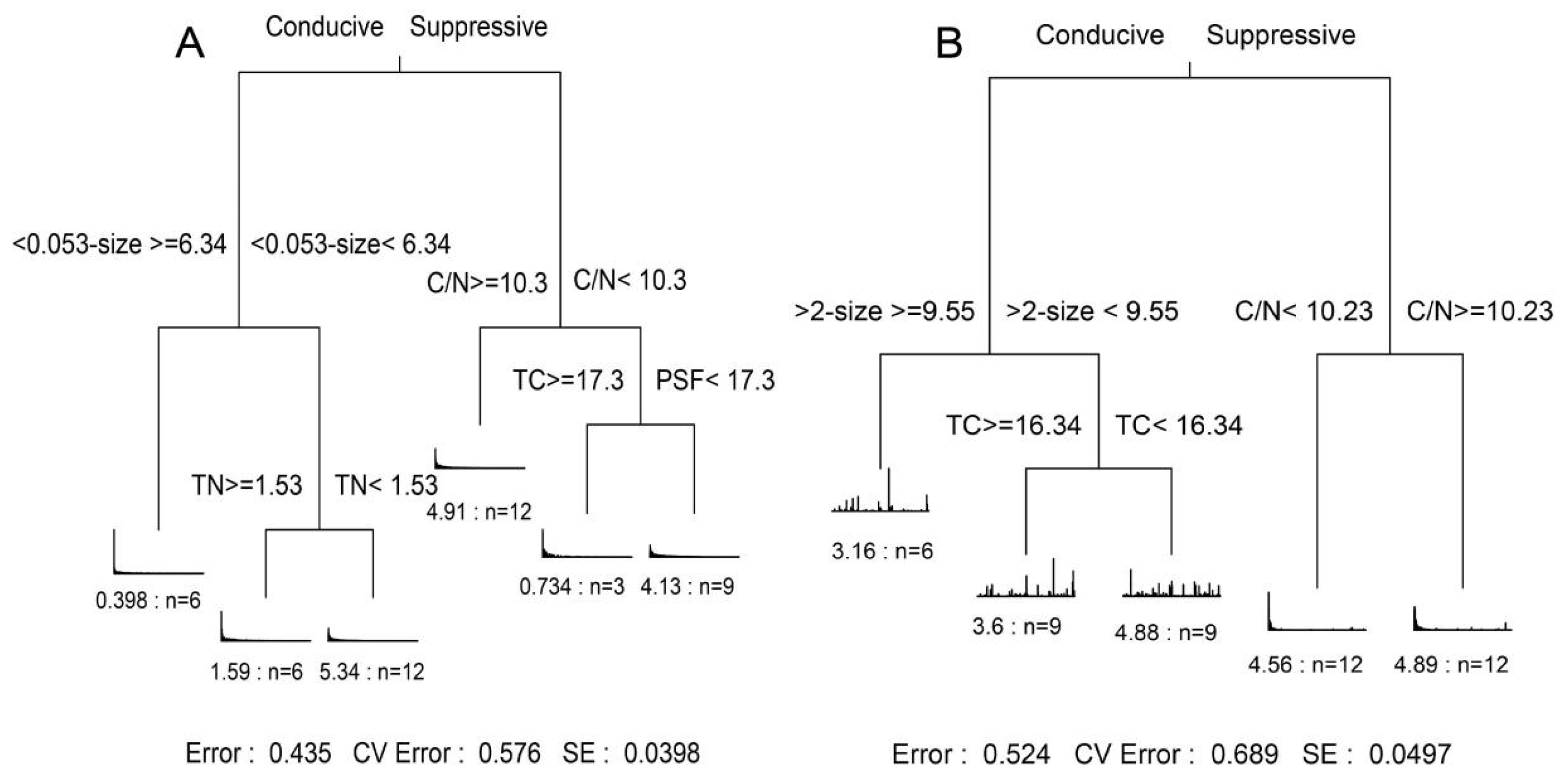
3.6. Deciphering the Direct and Indirect Pathways of Microbial Traits and Their Effects on Enzymatic Patterns in Suppressive and Conducive Soils
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Pegg, K.; Moore, N.; Bentley, S. Fusarium wilt of banana in Australia: A review. Aust. J. Agric. Res. 1996, 47, 637–650. [Google Scholar] [CrossRef]
- Butler, D. Fungus Threatens Top Banana. Nature 2013, 504, 195–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.F.; Chen, W.; Huang, X.; Hu, X.; Zhao, J.T.; Gong, Q.; Li, X.J. Fusarium wilt-resistant lines of Brazil banana (Musa spp., AAA) obtained by EMS-induced mutation in a micro-cross-section cultural system. Plant Pathol. 2013, 62, 112–119. [Google Scholar] [CrossRef]
- Alabouvette, C. Fusarium wilt suppressive soils: An example of disease-suppressive soils. Australas. Plant Pathol. 1999, 28, 57–64. [Google Scholar] [CrossRef]
- Fravel, D.; Olivain, C.; Alabouvette, C. Fusarium oxysporum and its biocontrol. New Phytol. 2003, 157, 493–502. [Google Scholar] [CrossRef] [PubMed]
- Nel, B.; Steinberg, C.; Labuschagne, N.; Viljoen, A. The potential of nonpathogenic Fusarium oxysporum and other biological control organisms for suppressing fusarium wilt of banana. Plant Pathol. 2006, 55, 217–223. [Google Scholar] [CrossRef]
- Xiong, W.; Zhao, Q.; Xue, C.; Xun, W.; Zhao, J.; Wu, H.; Li, R.; Shen, Q. Comparison of Fungal Community in Black Pepper-Vanilla and Vanilla Monoculture Systems Associated with Vanilla Fusarium Wilt Disease. Front. Microbiol. 2016, 7, 117. [Google Scholar] [CrossRef]
- Shen, Z.; Ruan, Y.; Xue, C.; Zhong, S.; Li, R.; Shen, Q. Soils naturally suppressive to banana Fusarium wilt disease harbor unique bacterial communities. Plant Soil 2015, 393, 21–33. [Google Scholar] [CrossRef]
- Xiong, W.; Zhao, Q.; Zhao, J.; Xun, W.; Li, R.; Zhang, R.; Wu, H.; Shen, Q. Different Continuous Cropping Spans Significantly Affect Microbial Community Membership and Structure in a Vanilla-Grown Soil as Revealed by Deep Pyrosequencing. Microb. Ecol. 2014, 70, 209–218. [Google Scholar] [CrossRef]
- Xiong, W.; Li, R.; Ren, Y.; Liu, C.; Zhao, Q.; Wu, H.; Jousset, A.; Shen, Q. Distinct roles for soil fungal and bacterial communities associated with the suppression of vanilla Fusarium wilt disease. Soil Biol. Biochem. 2017, 107, 198–207. [Google Scholar] [CrossRef]
- Boudreau, M.A. Factors Influencing Antagonism of Chaetomium globosum to Venturia inaequalis: A Case Study in Failed Biocontrol. Phytopathology 1987, 77, 1470–1475. [Google Scholar] [CrossRef]
- Kubicek, C.P.; Mach, R.L.; Peterbauer, C.K.; Lorito, M. Trichoderma: From Genes to Biocontrol. J. Plant Pathol. 2001, 83, 11–23. [Google Scholar]
- De Boer, M.; Bom, P.; Kindt, F.; Keurentjes, J.J.B.; van der Sluis, I.; van Loon, L.C.; Bakker, P.A.H.M. Control of Fusarium Wilt of Radish by Combining Pseudomonas putida Strains that have Different Disease-Suppressive Mechanisms. Phytopathology 2003, 93, 626–632. [Google Scholar] [CrossRef] [Green Version]
- Kyselková, M.; Kopecky, J.; Frapolli, M.; Défago, G.; Ságová-Marečková, M.; Grundmann, G.L.; Moënne-Loccoz, Y. Comparison of rhizobacterial community composition in soil suppressive or conducive to tobacco black root rot disease. ISME J. 2009, 3, 1127–1138. [Google Scholar] [CrossRef] [Green Version]
- Huse, S.M.; Dethlefsen, L.; Huber, J.; Welch, D.M.; Relman, D.; Sogin, M.L. Exploring Microbial Diversity and Taxonomy Using SSU rRNA Hypervariable Tag Sequencing. PLoS Genet. 2008, 4, e1000255. [Google Scholar] [CrossRef]
- Huang, X.; Liu, L.; Wen, T.; Zhu, R.; Zhang, J.; Cai, Z. Illumina MiSeq investigations on the changes of microbial community in the Fusarium oxysporum f. sp. cubense infected soil during and after reductive soil disinfestation. Microbiol. Res. 2015, 181, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Six, J.; Paustian, K.; Elliott, E.T.; Combrink, C. Soil Structure and Organic Matter I. Distribution of Aggregate-Size Classes and Aggregate-Associated Carbon. Soil Sci. Soc. Am. J. 2000, 64, 681–689. [Google Scholar] [CrossRef]
- Dominguez, J.; Negrin, M.A.; Rodriguez, C.M. Aggregate water-stability, particle-size and soil solution properties in conducive and suppressive soils to Fusarium wilt of banana from Canary Islands (Spain). Soil Biol. Biochem. 2001, 33, 449–455. [Google Scholar] [CrossRef]
- Miller, M.; Dick, R.P. Dynamics of soil C and microbial biomass in whole soil and aggregates in two cropping systems. Appl. Soil Ecol. 1995, 2, 253–261. [Google Scholar] [CrossRef]
- Zhang, P.; Zheng, J.; Pan, G.; Zhang, X.; Li, L.; Rolf, T. Changes in microbial community structure and function within particle size fractions of a paddy soil under different long-term fertilization treatments from the Tai Lake region, China. Colloids Surf. B Biointerfaces 2007, 58, 264–270. [Google Scholar] [CrossRef]
- Roldán, A.; Salinas-García, J.; Alguacil, M.; Caravaca, F. Changes in soil enzyme activity, fertility, aggregation and C sequestration mediated by conservation tillage practices and water regime in a maize field. Appl. Soil Ecol. 2005, 30, 11–20. [Google Scholar] [CrossRef]
- Oades, J. The role of biology in the formation, stabilization and degradation of soil structure. Geoderma 1993, 56, 377–400. [Google Scholar] [CrossRef]
- Cheshire, M.V.; Sparling, G.P.; Mundie, C.M. Effect of periodate treatment of soil on carbohydrate constituents and soil aggregation. Eur. J. Soil Sci. 1983, 34, 105–112. [Google Scholar] [CrossRef]
- Bastida, F.; Moreno, J.L.; Hernandez, T.; García, C. Microbiological activity in a soil 15 years after its devegetation. Soil Biol. Biochem. 2006, 38, 2503–2507. [Google Scholar] [CrossRef]
- Loeppmann, S.; Blagodatskaya, E.; Pausch, J.; Kuzyakov, Y. Substrate quality affects kinetics and catalytic efficiency of exoenzymes in rhizosphere and detritusphere. Soil Biol. Biochem. 2016, 92, 111–118. [Google Scholar] [CrossRef]
- Strickland, M.S.; Rousk, J. Considering fungal:bacterial dominance in soils—Methods, controls, and ecosystem implications. Soil Biol. Biochem. 2010, 42, 1385–1395. [Google Scholar] [CrossRef]
- Basaran, P.; Ozcan, M.; Denisov, Y.; Freeman, S. Elucidation of Pectinolytic Enzyme Activities of a Non-Pathogenic Watermelon Pathogen Mutant, Fusarium oxysporum f.sp Niveum M87. Australas. Plant Pathol. 2007, 36, 135–141. [Google Scholar] [CrossRef]
- Giannakis, C.; Bucheli, C.S.; Skene, K.G.M.; Robinson, S.P.; Scott, N.S. Chitinase and β-1,3-glucanase in grapevine leaves: A possible defence against powdery mildew infection. Aust. J. Grape Wine Res. 1998, 4, 14–22. [Google Scholar] [CrossRef]
- Huang, Y.; Li, C.; Wei, Y.; Yi, G. Effects of Fusarium oxysporum f. sp. Cubense Race 4 on Microorganisms and Enzyme Activities in the Rhizosphere Soil of Banana. J. Hunan Agric. Univ. 2012, 38, 173–176. [Google Scholar] [CrossRef]
- Yao, S.H.; Jia, L.J.; Zhang, X.W.; Zhang, D.; Xu, L. Biological Function Analysis of a Pectate Lyase Pela in Fusarium Graminerum. Plant Physiol. J. 2014, 50, 243–252. [Google Scholar]
- Elliott, E.T. Aggregate Structure and Carbon, Nitrogen, and Phosphorus in Native and Cultivated Soils. Soil Sci. Soc. Am. J. 1986, 50, 627–633. [Google Scholar] [CrossRef]
- Claesson, M.J.; O’Sullivan, O.; Wang, Q.; Nikkilä, J.; Marchesi, J.R.; Smidt, H.; De Vos, W.M.; Paul Ross, R.; O’Toole, P.W. Comparative Analysis of Pyrosequencing and a Phylogenetic Microarray for Exploring Microbial Community Structures in the Human Distal Intestine. PLoS ONE 2009, 4, e6669. [Google Scholar] [CrossRef] [Green Version]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes—Application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee SJ, W.T.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal Rna Genes for Phylogenetics. Pcr Protoc. A Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Lievens, B.; Brouwer, M.; Vanachter, A.C.R.C.; Levesque, C.A.; Cammue, B.P.A.; Thomma, B.P.H.J. Quantitative assessment of phytopathogenic fungi in various substrates using a DNA macroarray. Environ. Microbiol. 2005, 7, 1698–1710. [Google Scholar] [CrossRef] [PubMed]
- Muyzer, G.; de Waal, E.C.; Uitterlinden, A.G. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl. Environ. Microbiol. 1993, 59, 695–700. [Google Scholar] [CrossRef] [Green Version]
- May, L.A.; Smiley, B.; Schmidt, M.G. Comparative denaturing gradient gel electrophoresis analysis of fungal communities associated with whole plant corn silage. Can. J. Microbiol. 2001, 47, 829–841. [Google Scholar] [CrossRef]
- López-Mondéjar, R.; Antón, A.; Raidl, S.; Ros, M.; Pascual, J. Quantification of the biocontrol agent Trichodermaharzianum with real-time TaqMan PCR and its potential extrapolation to the hyphal biomass. Bioresour. Technol. 2010, 101, 2888–2891. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Edwards, J.; Johnson, C.; Santos-Medellin, C.; Lurie, E.; Podishetty, N.K.; Bhatnagar, S.; Eisen, J.A.; Sundaresan, V. Structure, variation, and assembly of the root-associated microbiomes of rice. Proc. Natl. Acad. Sci. USA 2015, 112, E911–E920. [Google Scholar] [CrossRef] [Green Version]
- Grace, J.B. Structural Equation Modeling and Natural Systems; Cambridge University Press: Cambridge, UK, 2006. [Google Scholar] [CrossRef]
- Schermelleh-Engel, K.; Moosbrugger, H.; Müller, H. Evaluating the Fit of Structural Equation Models: Tests of Significance and Descriptive Goodness-of-Fit Measures. Methods Psychol. Res. Online 2003, 8, 23–74. [Google Scholar]
- Xu, L.; Huang, B.; Wu, Y.; Huang, Y.; Dong, T. The Cost-Benefit Analysis for Bananas Diver-sity Production in China Foc. Zones. Am. J. Plant Sci. 2011, 2, 561–568. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Li, R.; Ruan, Y.; Ou, Y.; Zhao, Y.; Shen, Q. Pineapple–banana rotation reduced the amount of Fusarium oxysporum more than maize–banana rotation mainly through modulating fungal communities. Soil Biol. Biochem. 2015, 86, 77–86. [Google Scholar] [CrossRef]
- He, Y.; Zhou, X.; Li, J.; Li, H.; Li, Y.; Nie, Y. In Vitro Secretome Analysis Suggests Differential Pathogenic Mechanisms between Fusarium oxysporum f. sp. cubense Race 1 and Race 4. Biomolecules 2021, 11, 1353. [Google Scholar] [CrossRef] [PubMed]
- Kema, G.H.J.; Drenth, A.; Dita, M.; Jansen, K.; Vellema, S.; Stoorvogel, J.J. Editorial: Fusarium Wilt of Banana, a Recurring Threat to Global Banana Production. Front. Plant Sci. 2021, 11, 2177. [Google Scholar] [CrossRef] [PubMed]
- Thangavelu, R.; Edwin Raj, E.; Muthukathan, G.; Murugan, L.; Uma, S. Draft Genome Resource of a Novel Virulent Fusarium oxysporum f. sp. cubense Race 1 Strain (VCG 0124) Infecting Cavendish (AAA) Group of Banana in India. Plant Dis. 2021, 105, 2708–2710. [Google Scholar] [CrossRef]
- Wang, X.; Yu, R.; Li, J. Using Genetic Engineering Techniques to Develop Banana Cultivars with Fusarium Wilt Resistance and Ideal Plant Architecture. Front. Plant Sci. 2021, 11, 2202. [Google Scholar] [CrossRef]
- Mazzola, M. Assessment and Management of Soil Microbial Community Structure for Disease Suppression. Annu. Rev. Phytopathol. 2004, 42, 35–59. [Google Scholar] [CrossRef]
- Manici, L.; Caputo, F. Fungal community diversity and soil health in intensive potato cropping systems of the east Po valley, northern Italy. Ann. Appl. Biol. 2009, 155, 245–258. [Google Scholar] [CrossRef]
- Kim, Y.C.; Leveau, J.; Gardener, B.B.M.; Pierson, E.A.; Pierson, L.S.; Ryu, C.-M. The Multifactorial Basis for Plant Health Promotion by Plant-Associated Bacteria. Appl. Environ. Microbiol. 2011, 77, 1548–1555. [Google Scholar] [CrossRef] [Green Version]
- Palaniyandi, S.A.; Yang, S.H.; Zhang, L.; Suh, J.-W. Effects of actinobacteria on plant disease suppression and growth promotion. Appl. Microbiol. Biotechnol. 2013, 97, 9621–9636. [Google Scholar] [CrossRef] [PubMed]
- Mendes, R.; Kruijt, M.; de Bruijn, I.; Dekkers, E.; Van Der Voort, M.; Schneider, J.H.; Piceno, Y.M.; DeSantis, T.Z.; Andersen, G.L.; Bakker, P.A.; et al. Deciphering the Rhizosphere Microbiome for Disease-Suppressive Bacteria. Science 2011, 332, 1097–1100. [Google Scholar] [CrossRef] [PubMed]
- Kavino, M.; Harish, S.; Kumar, N.; Saravanakumar, D.; Samiyappan, R. Effect of chitinolytic PGPR on growth, yield and physiological attributes of banana (Musa spp.) under field conditions. Appl. Soil Ecol. 2010, 45, 71–77. [Google Scholar] [CrossRef]
- Buysens, S.; Heungens, K.; Poppe, J.; Hofte, M. Involvement of Pyochelin and Pyoverdin in Suppression of Pythium-Induced Damping-Off of Tomato by Pseudomonas Aeruginosa 7nsk2. Appl. Environ. Microbiol. 1996, 62, 865–871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berta, G.; Sampo, S.; Gamalero, E.; Massa, N.; Lemanceau, P. Suppression of Rhizoctonia root-rot of tomato by Glomus mossae BEG12 and Pseudomonas fluorescens A6RI is associated with their effect on the pathogen growth and on the root morphogenesis. Eur. J. Plant Pathol. 2005, 111, 279–288. [Google Scholar] [CrossRef]
- Wiggins, B.E.; Kinkel, L.L. Green manures and crop sequences influence alfalfa root rot and pathogen inhibitory activity among soil-borne streptomycetes. Plant Soil 2005, 268, 271–283. [Google Scholar] [CrossRef]
- Faheem, M.; Raza, W.; Zhong, W.; Nan, Z.; Shen, Q.; Xu, Y. Evaluation of the biocontrol potential of Streptomyces goshikiensis Ycxu against Fusarium oxysporum f. sp. Niveum. Biol. Control 2015, 81, 101–110. [Google Scholar] [CrossRef]
- Shen, T.; Wang, C.; Yang, H.; Deng, Z.; Wang, S.; Shen, B.; Shen, Q. Identification, solid-state fermentation and biocontrol effects of Streptomyces hygroscopicus B04 on strawberry root rot. Appl. Soil Ecol. 2016, 103, 36–43. [Google Scholar] [CrossRef]
- Yu, H.Y.; Ding, W.X.; Luo, J.F.; Geng, R.L.; Cai, Z.C. Long-term application of organic manure and mineral fertilizers on aggregation and aggregate-associated carbon in a sandy loam soil. Soil Tillage Res. 2012, 124, 170–177. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhou, W.; Liang, G.; Sun, J.; Wang, X.; He, P. Distribution of soil nutrients, extracellular enzyme activities and microbial communities across particle-size fractions in a long-term fertilizer experiment. Appl. Soil Ecol. 2015, 94, 59–71. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soils b | Particle Size (mm) | Fraction Amount (%) | TC c (g/kg) | TN (g/kg) | C/N |
|---|---|---|---|---|---|
| ST-S | >2 | 18.28 ± 4.65 b | 15.87 ± 0.06 b | 1.50 ± 0.36 b | 10.96 ± 0.42 a |
| 0.25–2 | 72.53 ± 5.93 a | 17.73 ± 1.17 a | 1.83 ± 0.21 a | 9.71 ± 0.49 c | |
| 0.052–0.25 | 5.50 ± 1.82 c | 18.63 ± 1.52 a | 1.90 ± 0.36 a | 9.96 ± 0.20 c | |
| <0.053 | 5.63 ± 0.83 c | 18.60 ± 0.20 a | 1.87 ± 0.35 a | 10.22 ± 0.09 b | |
| ST-C | >2 | 9.71 ± 3.31 b | 14.87 ± 0.31 c | 1.37 ± 0.15 c | 10.96 ± 0.15 a |
| 0.25–2 | 72.39 ± 5.35 a | 15.97 ± 0.55 b | 1.50 ± 0.10 b | 10.67 ± 0.35 a | |
| 0.052–0.25 | 4.99 ± 2.62 c | 16.70 ± 0.89 b | 1.52 ± 0.09 b | 11.17 ± 0.22 a | |
| <0.053 | 12.91 ± 1.07 b | 18.70 ± 1.59 a | 1.83 ± 0.15 a | 10.21 ± 0.16 b | |
| SD-S | >2 | 6.11 ± 3.68 b | 16.97 ± 0.71 a | 1.70 ± 0.36 a | 10.37 ± 0.27 b |
| 0.25–2 | 78.66 ± 5.53 a | 16.17 ± 1.12 a | 1.57 ± 0.15 b | 10.39 ± 0.32 b | |
| 0.052–0.25 | 8.68 ± 1.11 b | 17.47 ± 1.27 a | 1.63 ± 0.35 b | 11.04 ± 2.67 a | |
| <0.053 | 6.55 ± 2.32 b | 17.73 ± 3.00 a | 1.73 ± 0.32 a | 10.25 ± 0.30 b | |
| SD-C | >2 | 2.57 ± 0.61 c | 11.77 ± 1.72 c | 1.43 ± 0.49 b | 8.55 ± 0.47 c |
| 0.25–2 | 77.74 ± 2.67 a | 15.90 ± 1.57 b | 1.40 ± 0.26 b | 11.50 ± 0.36 a | |
| 0.052–0.25 | 11.04 ± 1.84 b | 15.47 ± 0.81 b | 1.43 ± 0.23 b | 10.91 ± 0.09 b | |
| <0.053 | 8.65 ± 1.93 b | 17.60 ± 0.62 a | 1.57 ± 0.12 a | 11.28 ± 0.17 a |
| Particle Size (mm) | Bacteria Shannon Diversity | Fungi Shannon Diversity | |||
|---|---|---|---|---|---|
| Conducive | Suppressive | Conducive | Suppressive | ||
| SD b | >2 | 6.23 ± 0.28 a | 6.27 ± 0.41 a | 3.73 ± 0.53 a | 4.00 ± 0.68 a |
| 2–0.25 | 6.16 ± 0.11 a | 6.24 ± 0.34 a | 3.08 ± 0.85 b | 4.15 ± 0.25 a | |
| 0.25–0.053 | 6.23 ± 0.37 a | 6.58 ± 0.17 a | 3.49 ± 0.51 a | 4.01 ± 0.45 a | |
| <0.053 | 3.27 ± 0.32 b | 3.70 ± 0.54 b | 3.20 ± 0.44 a | 2.13 ± 0.67 b | |
| ST | >2 | 6.34 ± 0.19 a | 6.30 ± 0.22 b | 2.93 ± 0.24 a | 3.15 ± 0.19 a |
| 2–0.25 | 6.60 ± 0.05 a | 6.65 ± 0.13 a | 2.90 ± 0.43 a | 3.10 ± 0.33 a | |
| 0.25–0.053 | 5.98 ± 0.20 a | 6.78 ± 0.11 a | 2.72 ± 0.37 a | 2.76 ± 0.99 a | |
| <0.053 | 3.96 ± 0.98 b | 4.58 ± 0.22 c | 3.25 ± 0.64 a | 3.45 ± 0.37 a | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Q.; Zhou, L.; Jin, H.; Cong, B.; Yang, H.; Wang, S. Investigating the Responses of Microbial Communities to Banana Fusarium Wilt in Suppressive and Conducive Soils Based on Soil Particle-Size Differentiation. Agronomy 2022, 12, 229. https://doi.org/10.3390/agronomy12020229
Wang Q, Zhou L, Jin H, Cong B, Yang H, Wang S. Investigating the Responses of Microbial Communities to Banana Fusarium Wilt in Suppressive and Conducive Soils Based on Soil Particle-Size Differentiation. Agronomy. 2022; 12(2):229. https://doi.org/10.3390/agronomy12020229
Chicago/Turabian StyleWang, Quanzhi, Limin Zhou, Han Jin, Bingcheng Cong, Hua Yang, and Shimei Wang. 2022. "Investigating the Responses of Microbial Communities to Banana Fusarium Wilt in Suppressive and Conducive Soils Based on Soil Particle-Size Differentiation" Agronomy 12, no. 2: 229. https://doi.org/10.3390/agronomy12020229