

Contribution to Improving the Chickpea (Cicer arietinum L.) Efficiency in Low-Phosphorus Farming Systems: Assessment of the Relationships between the P and N Nutrition, Nodulation Capacity and Productivity Performance in P-Deficient Field Conditions

 ,
,  ,
, 
Abstract
:
1. Introduction
2. Materials and Methods
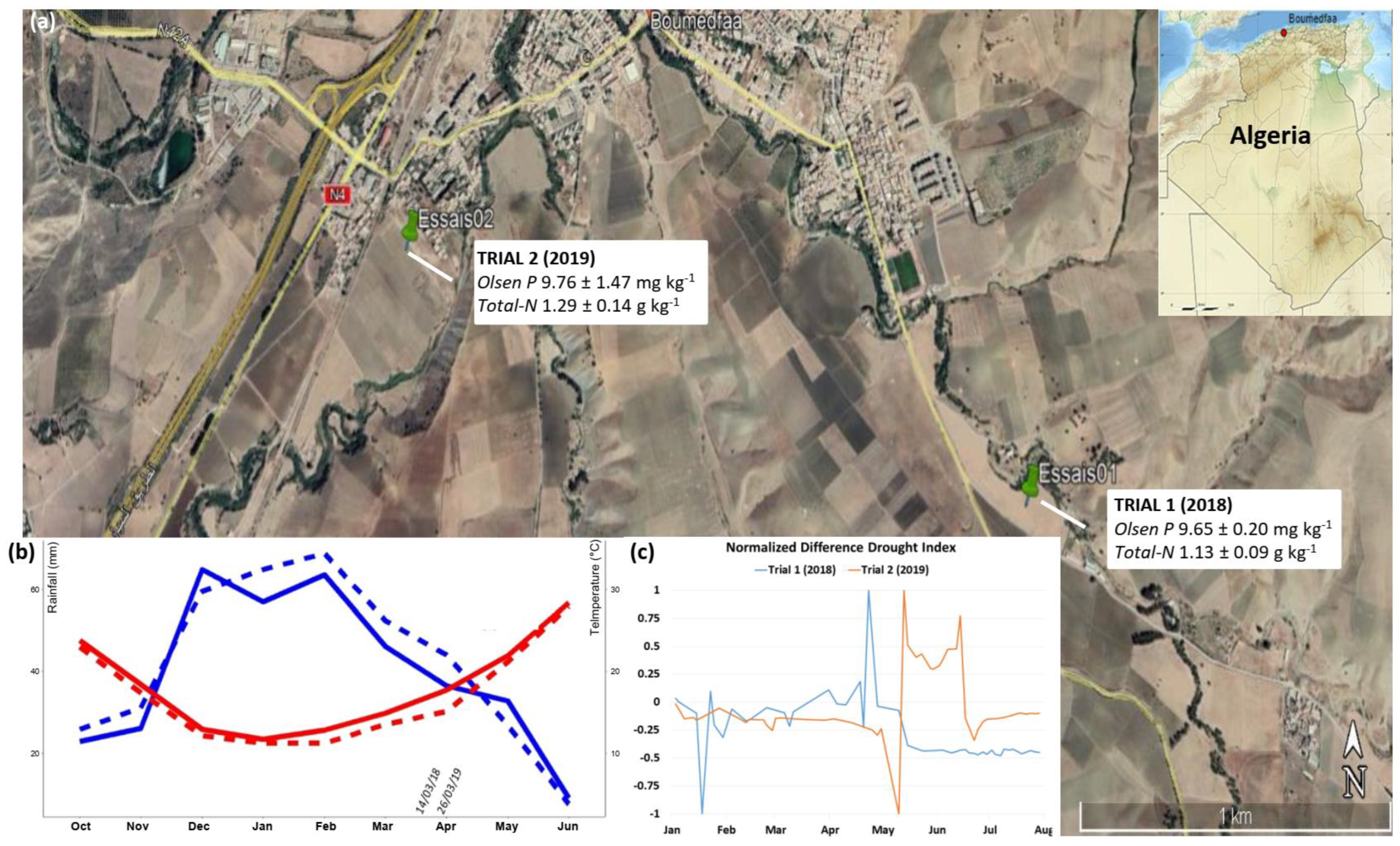
2.1. Description of the Field Experimental Sites
2.2. Plant Material and Experimental Design
2.3. Assessment of the Growth, Nodulation, and Yield Performances
2.4. In Vitro Seed Germination Assays
2.5. Quantification of the Nitrogen and Phosphorus Content in the Nodules, Shoots, and Rhizosphere
2.6. Statistical Analyses
3. Results
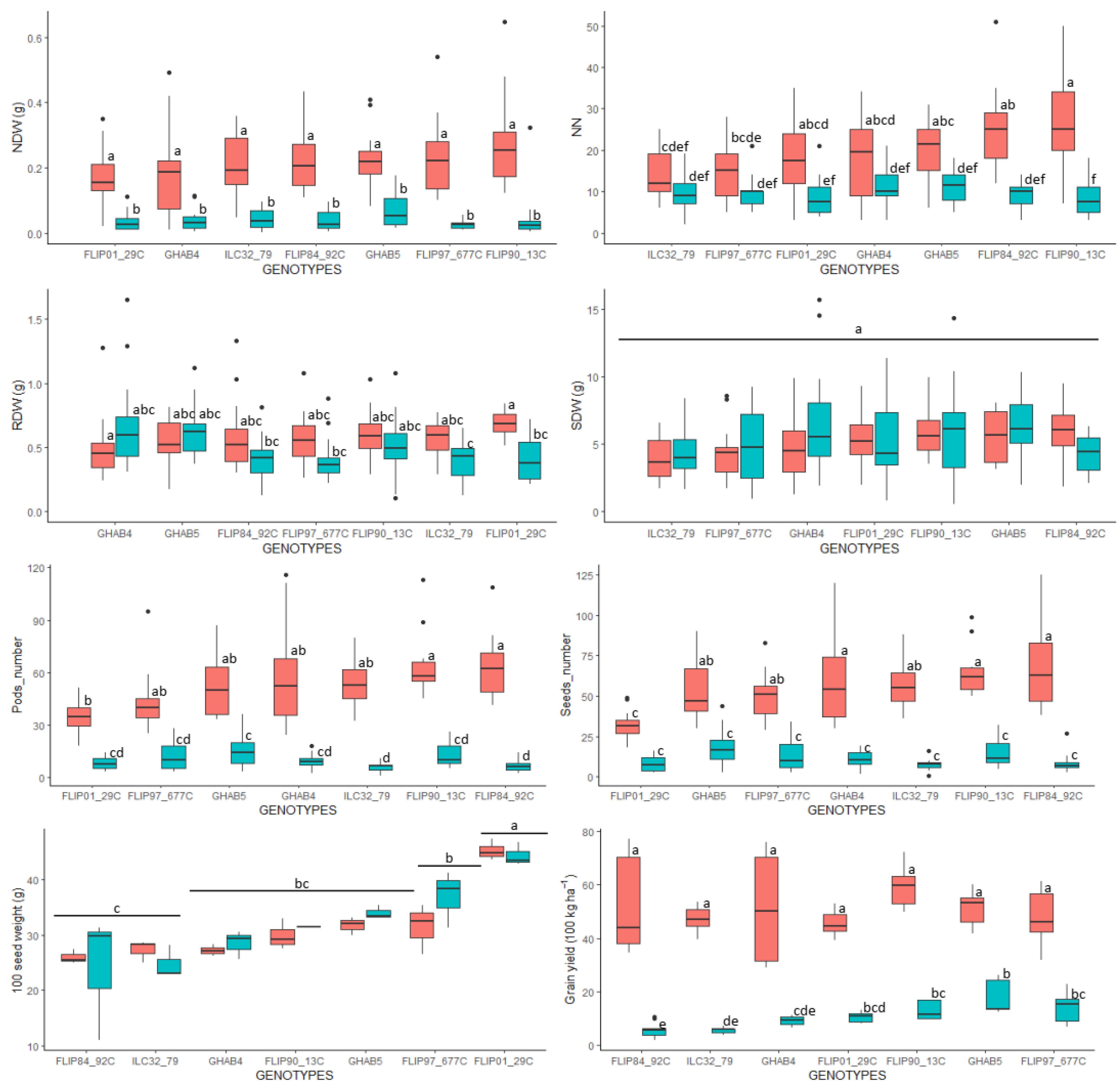
3.1. P-Deficiency Affects Certain Functional Growth and Nodulation Traits Differentially between Cultivars
3.2. P-Deficiency Differently Alters the Yield Components but Does Not Impact the Seed Germinability
3.3. P-Deficiency Can Hinder the Proper Establishment of the Symbiosis in a Cultivar-Dependent Manner
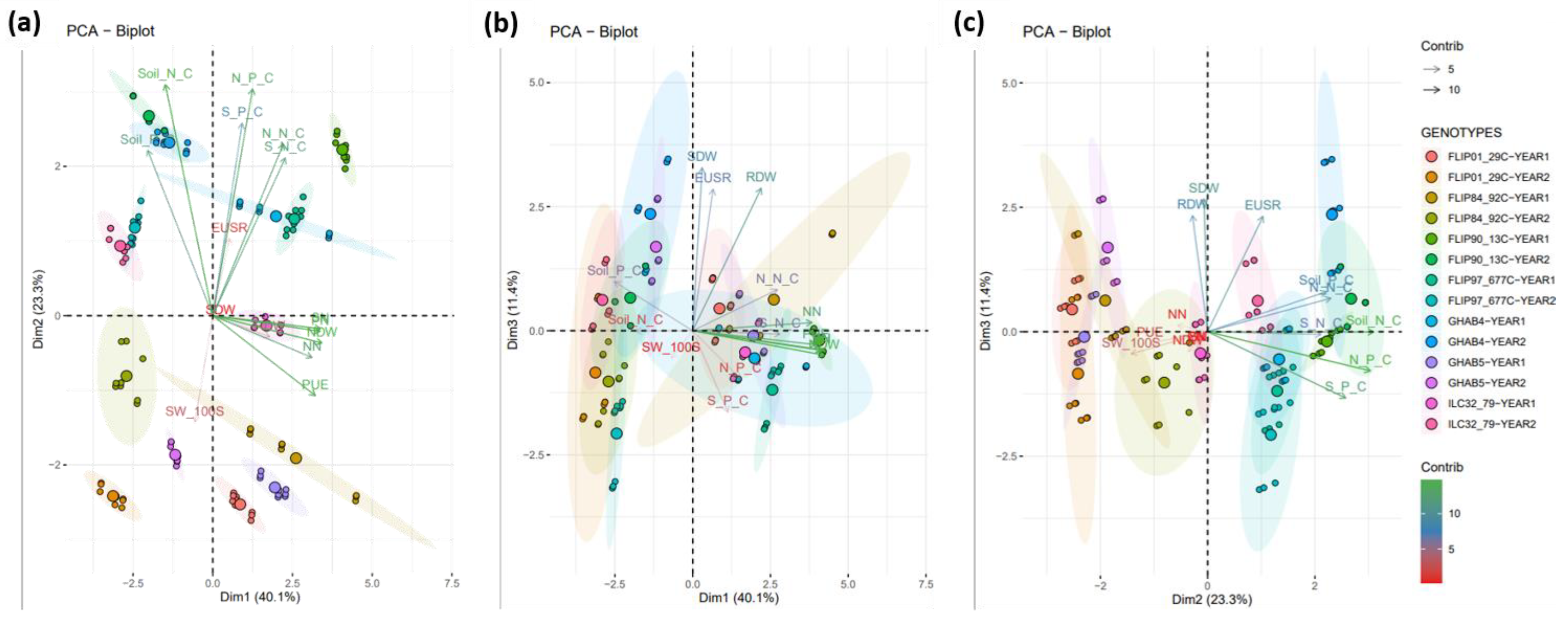
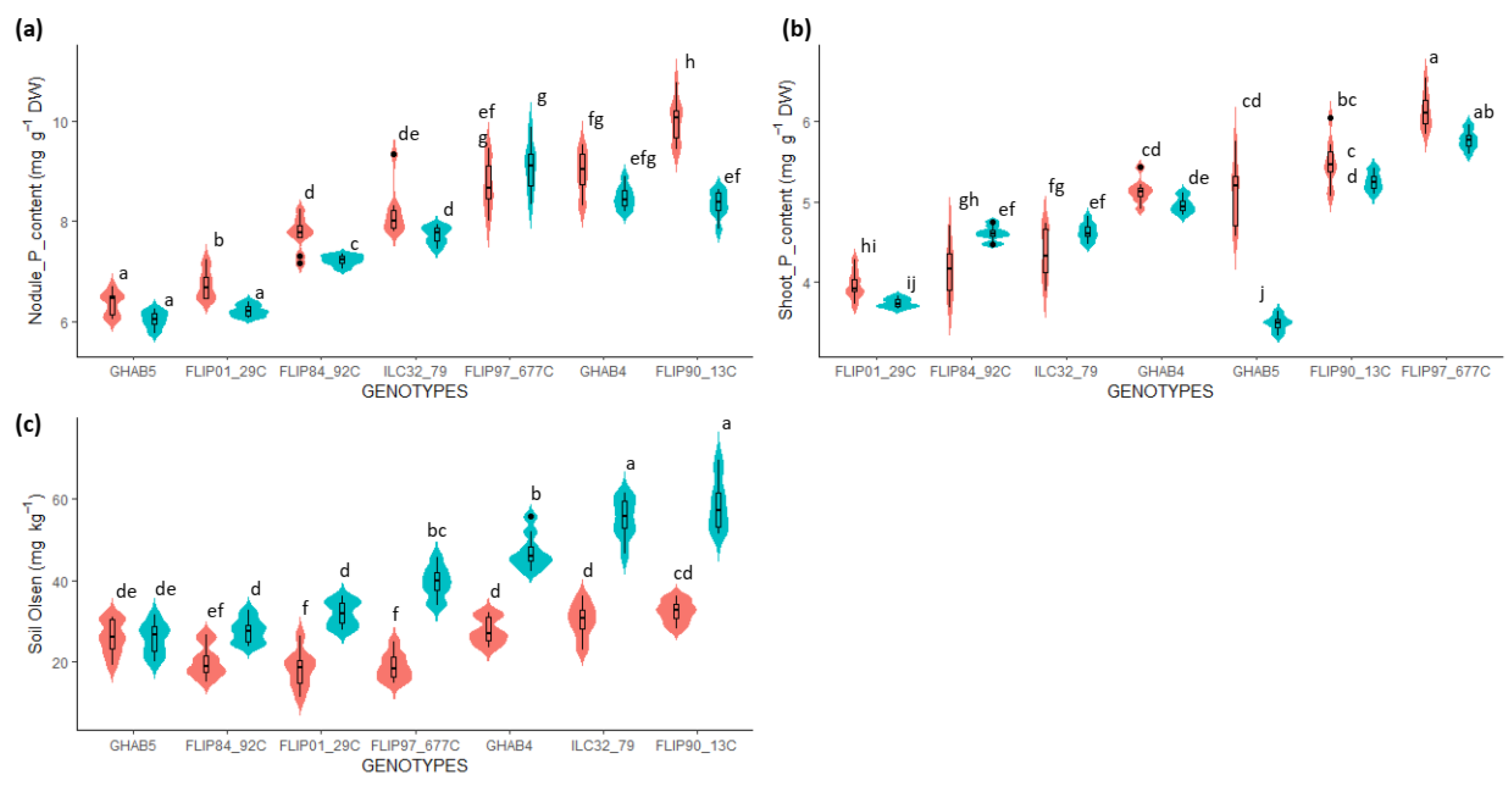
3.4. Phosphorus Acquisition and the Use Efficiency for the Nodule Formation Are Genetically Uncoupled
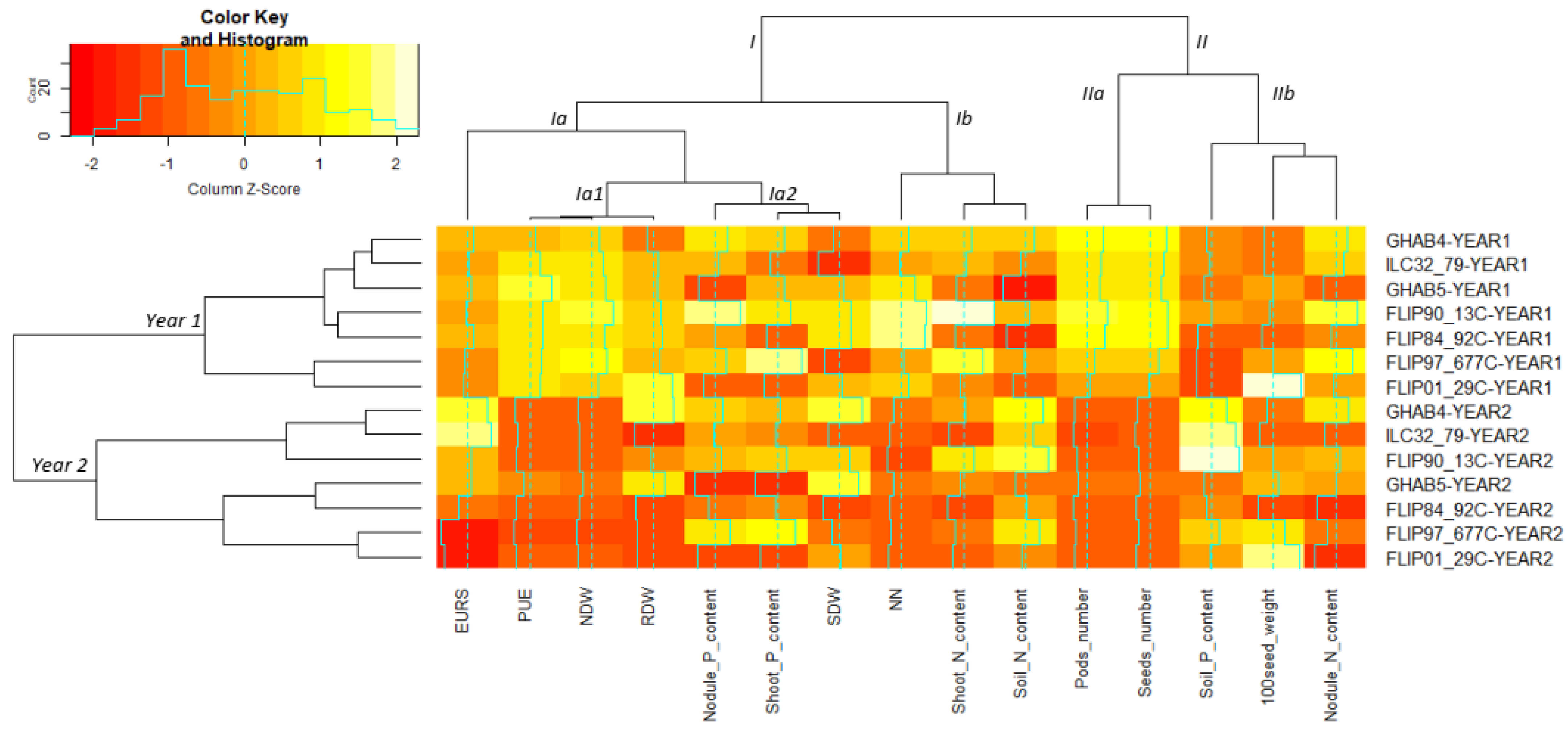
3.5. Functional Integrative Analysis Evidences the Efficient Varieties for Low-Input Agriculture
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Normalized Difference Drought Index (NDDI) Calculation
References
- Plaxton William, L.H. Annual Plant Reviews, Phosphorus Metabolism in Plants; Wiley-Blackwell: Hoboken, NJ, USA, 2015; Volume 48, ISBN 9780123643193. [Google Scholar]
- Tiziani, R.; Pii, Y.; Celletti, S.; Cesco, S.; Mimmo, T. Phosphorus Deficiency Changes Carbon Isotope Fractionation and Triggers Exudate Reacquisition in Tomato Plants. Sci. Rep. 2020, 10, 15970. [Google Scholar] [CrossRef] [PubMed]
- Bargaz, A.; Zaman-allah, M.; Farissi, M.; Lazali, M. Physiological and Molecular Aspects of Tolerance to Environmental Constraints in Grain and Forage Legumes. Int. J. Mol. Sci. 2015, 16, 18976–19008. [Google Scholar] [CrossRef] [Green Version]
- Lazali, M.; Bargaz, A. Examples of Belowground Mechanisms Enabling Legumes to Mitigate Phosphorus Deficiency. In Legume Nitrogen Fixation in Soils with Low Phosphorus Availability; Springer International Publishing: Cham, Switzerland, 2017; pp. 135–152. ISBN 9783319557298. [Google Scholar]
- Wen, Z.; Pang, J.; Tueux, G.; Liu, Y.; Shen, J.; Ryan, M.H.; Lambers, H.; Siddique, K.H.M. Contrasting Patterns in Biomass Allocation, Root Morphology and Mycorrhizal Symbiosis for Phosphorus Acquisition among 20 Chickpea Genotypes with Different Amounts of Rhizosheath Carboxylates. Funct. Ecol. 2020, 34, 1311–1324. [Google Scholar] [CrossRef]
- Tiziani, R.; Puschenreiter, M.; Smolders, E.; Mimmo, T.; Herrera, J.C.; Cesco, S.; Santner, J. Millimetre-Resolution Mapping of Citrate Exuded from Soil-Grown Roots Using a Novel, Low-Invasive Sampling Technique. J. Exp. Bot. 2021, 72, 3513–3525. [Google Scholar] [CrossRef]
- Kohli, P.S.; Kumar Verma, P.; Verma, R.; Parida, S.K.; Thakur, J.K.; Giri, J. Genome-Wide Association Study for Phosphate Deficiency Responsive Root Hair Elongation in Chickpea. Funct. Integr. Genom. 2020, 20, 775–786. [Google Scholar] [CrossRef] [PubMed]
- Prodhan, M.A.; Finnegan, P.M.; Lambers, H. How Does Evolution in Phosphorus-Impoverished Landscapes Impact Plant Nitrogen and Sulfur Assimilation? Trends Plant Sci. 2019, 24, 69–82. [Google Scholar] [CrossRef] [PubMed]
- Lazali, M.; Blavet, D.; Pernot, C.; Desclaux, D.; Drevon, J.J. Efficiency of Phosphorus Use for Dinitrogen Fixation Varies between Common Bean Genotypes under Phosphorus Limitation. Agron. J. 2017, 109, 283–290. [Google Scholar] [CrossRef]
- Han, Y.; White, P.J.; Cheng, L. Mechanisms for Improving Phosphorus Utilization Efficiency in Plants. Ann. Bot. 2022, 129, 247–258. [Google Scholar] [CrossRef]
- Lazali, M.; Brahimi, S.; Jacques, J. Rhizosphere High Yields in a Low-P Tolerant Recombinant Inbred Line of Common Bean under Fi Eld Conditions. Rhizosphere 2018, 8, 27–33. [Google Scholar] [CrossRef]
- Keneni, G.; Bekele, E.; Assefa, F.; Imtiaz, M.; Debele, T. Characterization of Ethiopian Chickpea (Cicer arietinum L.) Germplasm Accessions for Phosphorus Uptake and Use Efficiency II. Interrelationships of Characters and Gains from Selection. Ethiop. J. Appl. Sci. Technol. 2015, 6, 77–96. [Google Scholar]
- Sulieman, S.; Tran, L.S.P. Phosphorus Homeostasis in Legume Nodules as an Adaptive Strategy to Phosphorus Deficiency. Plant Sci. 2015, 239, 36–43. [Google Scholar] [CrossRef]
- Drevon, J.J.; Abadie, J.; Alkama, N.; Andriamananjara, A.; Amenc, L.; Bargaz, A.; Carlssonn, G.; Jaillard, B.; Lazali, M.; Ghoulam, C.; et al. Phosphorus Use Efficiency for N2 Fixation in the Rhizobial Symbiosis with Legumes. Biol. Nitrogen Fixat. 2015, 1–2, 455–464. [Google Scholar] [CrossRef]
- Abdelrahman, M.; El-sayed, M.A.; Hashem, A.; Valentine, A.J. Metabolomics and Transcriptomics in Legumes Under Phosphate Deficiency in Relation to Nitrogen Fixation by Root Nodules. Front. Plant Sci. 2018, 9, 922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, J.; Ryan, M.H.; Lambers, H.; Siddique, K.H. Phosphorus Acquisition and Utilisation in Crop Legumes under Global Change. Curr. Opin. Plant Biol. 2018, 45, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Varshney, R.K.; Roorkiwal, M.; Sun, S.; Bajaj, P.; Chitikineni, A.; Thudi, M.; Singh, N.P.; Du, X.; Upadhyaya, H.D.; Khan, A.W.; et al. A Chickpea Genetic Variation Map Based on the Sequencing of 3366 Genomes. Nature 2021, 599, 622–627. [Google Scholar] [CrossRef]
- Giller, K.E. Nitrogen fixation in tropical cropping systems; CAB International: Wallingford, UK, 2001. [Google Scholar]
- Khaitov, B.; Abdiev, A. Performance of chickpea (Cicer arietinum L.) to bio-fertilizer and nitrogen application in arid condition. J. Plant Nutr. 2018, 41, 1980–1987. [Google Scholar] [CrossRef]
- Alem, C.; Asres, T. Participatory evaluation and selection of chickpea varieties at Debre Mawi and Debre Yakob watersheds, Western Amhara Region, Ethiopia. In Proceedings of the Harnessing Chickpea Value Chain for Nutrition Security and Commercialization of Smallholder Agriculture in Africa, Debre Zeit, Ethiopia, 30 January–1 February 2014; Korbu, L., Damte, T., Fikre, A., Eds.; 2016; pp. 163–167. [Google Scholar]
- Ullah, A.; Farooq, M.; Rehman, A.; Hussain, M.; Siddique, K.H.M. Zinc Nutrition in Chickpea (Cicer arietinum): A Review. Crop Pasture Sci. 2020, 71, 199–218. [Google Scholar] [CrossRef]
- Abebe, T.N.; Beyene, S.; Hawassa, E. Growth Limiting Nutrient(s) and Their Effects on the Yield and Nutrient Uptake of Chickpea (Cicer arietinum L.) in Nitisols, Southern Ethiopia. J. Environ. Earth Sci. 2021, 11, 12. [Google Scholar] [CrossRef]
- Lazali, M.; Boudsocq, S.; Taschen, E.; Farissi, M.; Hamdi, W.; Ralli, P.; Sentenac, H. Crosymed Project: Enhancing Nutrient Use Efficiency through Legumes in Agroecosystems of the Mediterranean Basin. Sustainability 2021, 13, 4695. [Google Scholar] [CrossRef]
- Gu, Y.; Brown, J.F.; Verdin, J.P.; Wardlow, B. A Five-Year Analysis of MODIS NDVI and NDWI for Grassland Drought Assessment over the Central Great Plains of the United States. Geophys. Res. Lett. 2007, 34, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Du, T.L.T.; Du Bui, D.; Nguyen, M.D.; Lee, H. Satellite-Based, Multi-Indices for Evaluation of Agricultural Droughts in a Highly Dynamic Tropical Catchment, Central Vietnam. Water 2018, 10, 659. [Google Scholar] [CrossRef]
- Latati, M.; Blavet, D.; Alkama, N.; Laoufi, H.; Drevon, J.J.; Gérard, F.; Pansu, M.; Ounane, S.M. The Intercropping Cowpea-Maize Improves Soil Phosphorus Availability and Maize Yields in an Alkaline Soil. Plant Soil 2014, 385, 181–191. [Google Scholar] [CrossRef]
- Yong, T.W.; Chen, P.; Dong, Q.; Du, Q.; Yang, F.; Wang, X.; Liu, W.C.; Yang, W.Y. Optimized Nitrogen Application Methods to Improve Nitrogen Use Efficiency and Nodule Nitrogen Fixation in a Maize-Soybean Relay Intercropping System. J. Integr. Agric. 2018, 17, 664–676. [Google Scholar] [CrossRef]
- Zhang, J.; Peng, S.; Shang, Y.; Brunel, B.; Li, S.; Zhao, Y.; Liu, Y.; Chen, W.; Wang, E.; Singh, R.P.; et al. Genomic Diversity of Chickpea-Nodulating Rhizobia in Ningxia (North Central China) and Gene Flow within Symbiotic Mesorhizobium Muleiense Populations. Syst. Appl. Microbiol. 2020, 43, 126089. [Google Scholar] [CrossRef] [PubMed]
- Aydi, S.; Drevon, J.J.; Abdelly, C. Effect of Salinity on Root-Nodule Conductance to the Oxygen Diffusion in the Medicago Truncatula-Sinorhizobium Meliloti Symbiosis. Plant Physiol. Biochem. 2004, 42, 833–840. [Google Scholar] [CrossRef]
- Kjeldahl, J. New method for the determination of nitrogen in organic substances. Fresenius J. Anal. Chem. 1883, 22, 366–383. [Google Scholar] [CrossRef] [Green Version]
- Walkley, A.; Armstrong, B.I. An examination of the Degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Olsen, Sterling Robertson. Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate; No. 939; US Department of Agriculture: Washington, DC, USA, 1954.
- Valizadeh, G.R.; Rengel, Z.; Rate, A.W. Response of wheat genotypes efficient in P utilisation and genotypes responsive to P fertilisation to different P banding depths and watering regimes. Aust. J. Agric. Res. 2003, 54, 59–65. [Google Scholar] [CrossRef]
- Team RC. R: A Language and Environment for Statistical Computing. In R Foundation for Statistical Computing [Internet]; Team RC: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 8 December 2022).
- Sarkar, D.; Kar, S.K.; Chattopadhyay, A.; Shikha; Rakshit, A.; Tripathi, V.K.; Dubey, P.K.; Abhilash, P.C. Low Input Sustainable Agriculture: A Viable Climate-Smart Option for Boosting Food Production in a Warming World. Ecol. Indic. 2020, 115, 106412. [Google Scholar] [CrossRef]
- Cesco, S.; Lucini, L.; Miras-Moreno, B.; Borruso, L.; Mimmo, T.; Pii, Y.; Puglisi, E.; Spini, G.; Taskin, E.; Tiziani, R.; et al. The Hidden Effects of Agrochemicals on Plant Metabolism and Root-Associated Microorganisms. Plant Sci. 2021, 311, 111012. [Google Scholar] [CrossRef]
- Pang, J.; Megan, H.; Lambers, H.; Siddique, K.H.M. Leaf Transpiration Plays a Role in Phosphorus Acquisition among a Large Set of Chickpea Genotypes. Plant Cell Environ. 2018, 41, 2069–2079. [Google Scholar] [CrossRef] [PubMed]
- Srinivasarao, C.; Ganeshamurthy, A.; Ali, M.; Venkateswarlu, B. Phosphorus and Micronutrient Nutrition of Chickpea Genotypes in a Multi-Nutrient-Deficient Typic Ustochrept. J. Plant Nutr. 2006, 29, 747–763. [Google Scholar] [CrossRef]
- Liu, A.; Contador, C.A.; Fan, K.; Lam, H.M. Interaction and Regulation of Carbon, Nitrogen, and Phosphorus Metabolisms in Root Nodules of Legumes. Front. Plant Sci. 2018, 871, 1860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valentine, A.J.; Kleinert, A.; Benedito, V.A. Adaptive Strategies for Nitrogen Metabolism in Phosphate Deficient Legume Nodules. Plant Sci. 2017, 256, 46–52. [Google Scholar] [CrossRef]
- Zerihun, A. On-Farm Yield Variability and Responses of Common Bean (Phaseolus vulgaris L.) Varieties to Rhizobium Inoculation with Inorganic Fertilizer Rates. J. Anim. Plant Sci. 2017, 32, 5120–5133. [Google Scholar]
- Stevens, G.G.; Pérez-Fernández, M.A.; Morcillo, R.J.L.; Kleinert, A.; Hills, P.; Brand, D.J.; Steenkamp, E.T.; Valentine, A.J. Roots and Nodules Response Differently to P Starvation in the Mediterranean-Type Legume Virgilia Divaricata. Front. Plant Sci. 2019, 10, 73. [Google Scholar] [CrossRef] [Green Version]
- Farooq, M.; Gogoi, N.; Barthakur, S.; Baroowa, B.; Bharadwaj, N.; Alghamdi, S.S. Drought Stress in Grain Legumes during Reproduction and Grain Filling. J. Agron. Crop Sci. 2016, 203, 81–102. [Google Scholar] [CrossRef]
- Adusei, G.; Aidoo, M.K.; Srivastava, A.K.; Asibuo, J.Y.; Gaiser, T. The Variability of Grain Yield of Some Cowpea Genotypes in Response to Phosphorus and Water Stress under Field Conditions. Agronomy 2021, 11, 28. [Google Scholar] [CrossRef]
- Thudi, M.; Chen, Y.; Pang, J.; Kalavikatte, D.; Bajaj, P.; Roorkiwal, M.; Chitikineni, A.; Ryan, M.H.; Lambers, H.; Siddique, K.H.M.; et al. Novel Genes and Genetic Loci Associated With Root Morphological Traits, Phosphorus-Acquisition Efficiency and Phosphorus-Use Efficiency in Chickpea. Front. Plant Sci. 2021, 12, 636973. [Google Scholar] [CrossRef]
- Sulieman, S.; Kusano, M.; Van Ha, C.; Watanabe, Y.; Abdalla, M.A.; Abdelrahman, M.; Kobayashi, M.; Saito, K.; Mühling, K.H.; Tran, L.S.P. Divergent Metabolic Adjustments in Nodules Are Indispensable for Efficient N2 Fixation of Soybean under Phosphate Stress. Plant Sci. 2019, 289, 110249. [Google Scholar] [CrossRef]
- Sulieman, S.; Abdelrahman, M.; Tran, L.S.P. Carbon Metabolic Adjustment in Soybean Nodules in Response to Phosphate Limitation: A Metabolite Perspective. Environ. Exp. Bot. 2022, 196, 104810. [Google Scholar] [CrossRef]
- Allito, B.; Nana, E.-M.; Alemneh, A. Rhizobia Strain and Legume Genome Interaction Effects on Nitrogen Fixation and Yield of Grain Legume: A Review. Mol. Soil Biol. 2015, 6, 1–12. [Google Scholar] [CrossRef]
- Marra, L.M.; de Oliveira-Longatti, S.M.; Soares, C.R.F.S.; Olivares, F.L.; de Souza Moreira, F.M. The Amount of Phosphate Solubilization Depends on the Strain, C-Source, Organic Acids and Type of Phosphate. Geomicrobiol. J. 2019, 36, 232–242. [Google Scholar] [CrossRef]
- Mabrouk, Y.; Hemissi, I.; Salem, I.B.; Mejri, S.; Saidi, M.; Belhadj, O. Potential of rhizobia in improving nitrogen fixation and yields of legumes. Symbiosis 2018, 107, 73495. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SDW (g) | RDW (g) | NDW (g) | NN | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Source of Variation | DF | MS | F Value | Pr(>F) | MS | F Value | Pr(>F) | MS | F Value | Pr(>F) | MS | F Value | Pr(>F) |
| genotype | 6 | 0.54 | 2.72 | 0.01 * | 0.03 | 2.09 | 0.05 . | 0.02 | 3.09 | 0.006 ** | 0.26 | 2.68 | 0.02 * |
| year | 1 | 0.02 | 0.12 | 0.73 | 0.24 | 19.59 | 1.47 × 10−5 *** | 2.86 | 508.33 | <2.2 × 10−16 *** | 12.65 | 132.25 | <2 × 10−16 *** |
| block (year) | 4 | 0.51 | 2.55 | 0.04 * | 0.03 | 2.22 | 0.07. | 0.04 | 6.23 | 8.77 × 10−5 *** | 0.19 | 2.01 | 0.09 . |
| genotype × year | 6 | 0.34 | 1.71 | 0.12 | 0.08 | 6.92 | 8.80 × 10−7 *** | 0.01 | 2.26 | 0.04 * | 0.44 | 4.63 | 0.0002 *** |
| residuals | 234 | 0.20 | 0.01 | 0.01 | 0.10 | ||||||||
| Pod Numbers | Seed Numbers | |||||||
| Source of Variation | DF | MS | F Value | Pr(>F) | MS | F Value | Pr(>F) | |
| genotype | 6 | 0.61 | 5.72 | 1.67 × 10−5 *** | 0.77 | 6.11 | 6.95 × 10−6 *** | |
| year | 1 | 93.71 | 879.78 | <2.2 × 10−16 *** | 91.41 | 723.74 | <2.2 × 10−16 *** | |
| block (year) | 4 | 0.31 | 2.87 | 0.02 * | 0.29 | 2.32 | 0.06 . | |
| genotype × year | 6 | 0.59 | 5.57 | 2.35 × 10−5 *** | 0.52 | 4.08 | 0.0007 *** | |
| residuals | 195 | 0.11 | 0.13 | |||||
| 100 Seed Weights (g) | Grain Yield (100 kg ha−1) | |||||||
| Source of Variation | DF | MS | F Value | Pr(>F) | DF | MS | F Value | Pr(>F) |
| genotype | 6 | 1,309,448 | 32.19 | 2.44 × 10−10 *** | 6 | 0.63 | 10.29 | 5.38 × 10−9 *** |
| year | 1 | 48,312 | 1.19 | 0.29 | 1 | 82.23 | 1340.47 | <2.2 × 10−16 *** |
| block (year) | 4 | 59,857 | 1.47 | 0.24 | 4 | 0.37 | 6.06 | 0.0002 *** |
| genotype × year | 6 | 43,675 | 1.07 | 0.41 | 6 | 0.43 | 7.08 | 2.24 × 10−6 *** |
| residuals | 24 | 40,680 | 108 | 0.06 | ||||
| Nodule N Content (mg kg−1) | Shoot N Content (mg kg−1) | Soil N Content (mg kg−1) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Source of Variation | DF | MS | F Value | Pr(>F) | MS | F Value | Pr(>F) | MS | F Value | Pr(>F) |
| genotype | 6 | 52.64 | 70,973.88 | <2.2 × 10−16 *** | 11.14 | 25,156.44 | <2.2 × 10−16 *** | 204,371 | 3527.49 | <2 × 10−16 *** |
| year | 1 | 110.71 | 149,280.35 | <2.2 × 10−16 *** | 22.90 | 51,709.79 | <2 × 10−16 *** | 501,741 | 8660.16 | <2 × 10−16 *** |
| block (year) | 4 | 0.03 | 33.63 | <2.2 × 10−16 *** | 0.0001 | 0.16 | 0.96 | 95 | 1.64 | 0.17 |
| genotype × year | 6 | 21.23 | 28,624.30 | <2.2 × 10−16 *** | 2.32 | 5228.35 | <2.2 × 10−16 *** | 7547 | 130.27 | <2 × 10−16 *** |
| residuals | 108 | 0.001 | 0.0004 | 58 | ||||||
| Nodule P Content (mg g−1) | Shoot P Content (mg g−1) | Soil P Content (mg kg−1) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Source of Variation | DF | MS | F Value | Pr(>F) | MS | F Value | Pr(>F) | MS | F Value | Pr(>F) |
| genotype | 6 | 0.003 | 338.85 | <2.2 × 10−16 *** | 0.10 | 202.94 | <2.2 × 10−16 *** | 131.32 | 89.97 | <2 × 10−16 *** |
| year | 1 | 0.001 | 92.44 | 3.51 × 10−16 *** | 0.02 | 50.31 | 1.43 × 10−10 *** | 750.15 | 513.93 | <2 × 10−16 *** |
| block (year) | 4 | 0.00002 | 1.73 | 0.15 | 0.0001 | 0.28 | 0.89 | 4.08 | 2.79 | 0.03 * |
| genotype × year | 6 | 0.0001 | 9.32 | 3.17 × 10−8 *** | 0.02 | 50.95 | <2.2 × 10−16 *** | 35.09 | 24.04 | <2 × 10−16 *** |
| residuals | 108 | 0.00001 | 0.0005 | 1.46 | ||||||
| (a) | P Use Efficiency | |||
| Source of Variation | DF | MS | F value | Pr(>F) |
| genotype | 6 | 0.0001 | 3.04 | 0.02 * |
| year | 1 | 0.005 | 216.56 | 1.65 × 10−13 *** |
| block (year) | 4 | 0.00002 | 0.84 | 0.51 |
| genotype × year | 6 | 0.00001 | 0.49 | 0.81 |
| residuals | 24 | 0.00002 | ||
| (b) | Genotype | P Use Efficiency | ||
| GHAB4 | 0.0124 a | |||
| FLIP97_677C | 0.0147 ab | |||
| ILC32_79 | 0.0153 ab | |||
| FLIP01_29C | 0.0154 ab | |||
| FLIP90_13C | 0.0160 ab | |||
| FLIP84_92C | 0.0165 ab | |||
| GHAB5 | 0.0231 b | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Djouider, S.I.; Gentzbittel, L.; Jana, R.; Rickauer, M.; Ben, C.; Lazali, M. Contribution to Improving the Chickpea (Cicer arietinum L.) Efficiency in Low-Phosphorus Farming Systems: Assessment of the Relationships between the P and N Nutrition, Nodulation Capacity and Productivity Performance in P-Deficient Field Conditions. Agronomy 2022, 12, 3150. https://doi.org/10.3390/agronomy12123150
Djouider SI, Gentzbittel L, Jana R, Rickauer M, Ben C, Lazali M. Contribution to Improving the Chickpea (Cicer arietinum L.) Efficiency in Low-Phosphorus Farming Systems: Assessment of the Relationships between the P and N Nutrition, Nodulation Capacity and Productivity Performance in P-Deficient Field Conditions. Agronomy. 2022; 12(12):3150. https://doi.org/10.3390/agronomy12123150
Chicago/Turabian StyleDjouider, Souad Insaf, Laurent Gentzbittel, Raghavendra Jana, Martina Rickauer, Cécile Ben, and Mohamed Lazali. 2022. "Contribution to Improving the Chickpea (Cicer arietinum L.) Efficiency in Low-Phosphorus Farming Systems: Assessment of the Relationships between the P and N Nutrition, Nodulation Capacity and Productivity Performance in P-Deficient Field Conditions" Agronomy 12, no. 12: 3150. https://doi.org/10.3390/agronomy12123150