
Effects of Grazing Intensity on the Carbon, Nitrogen and Phosphorus Content, Stoichiometry and Storage of Plant Functional Groups in a Meadow Steppe

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Sampling
2.2. Sample Collection and Stoichiometry
2.3. Calculation and Statistics
3. Results
3.1. Changes in the Nutrient Content of Plant Functional Groups
3.2. Changes in the Stoichiometric Ratios of Plant Functional Groups
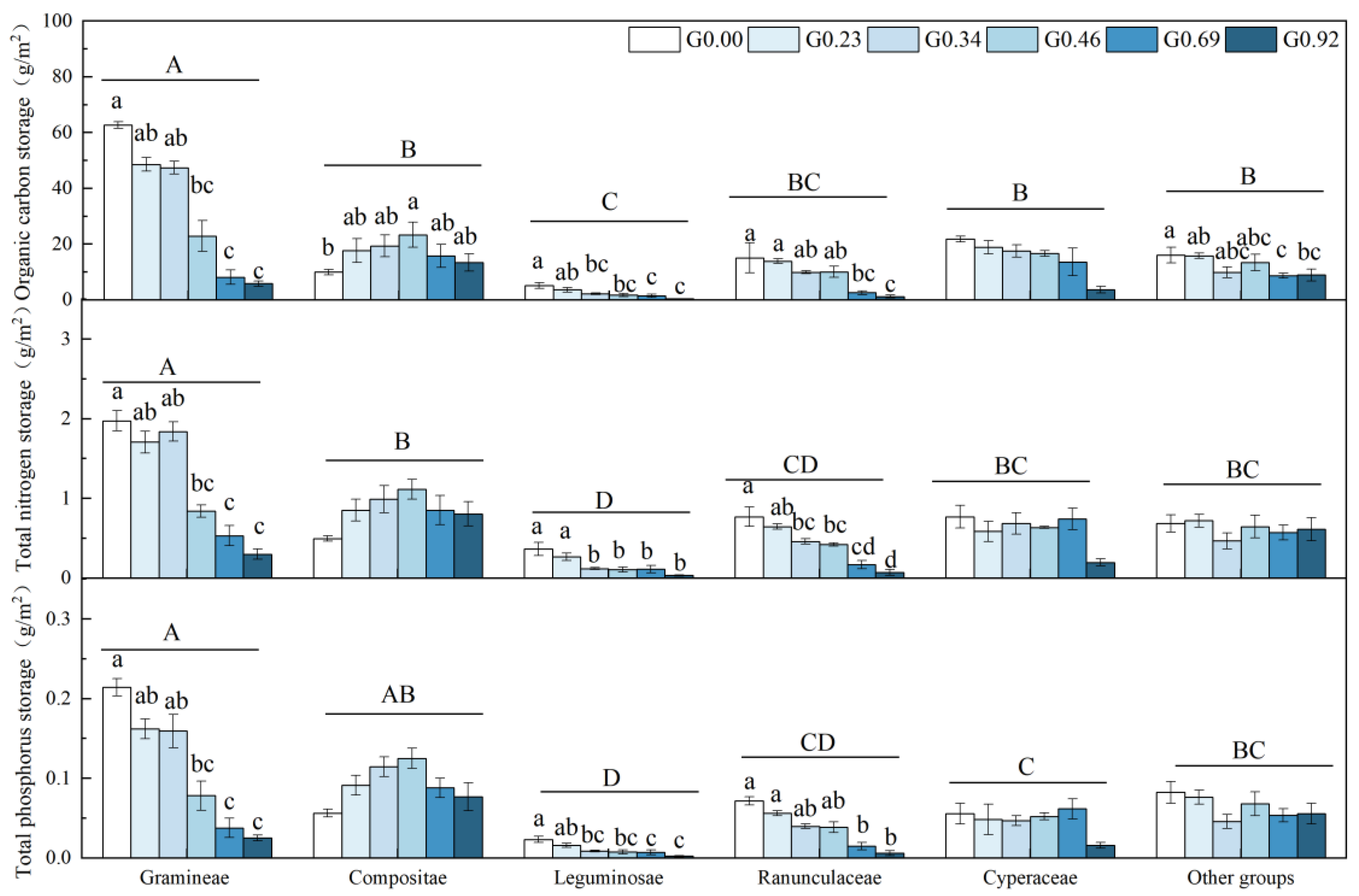
3.3. Changes in the Nutrient Storage of Plant Functional Groups
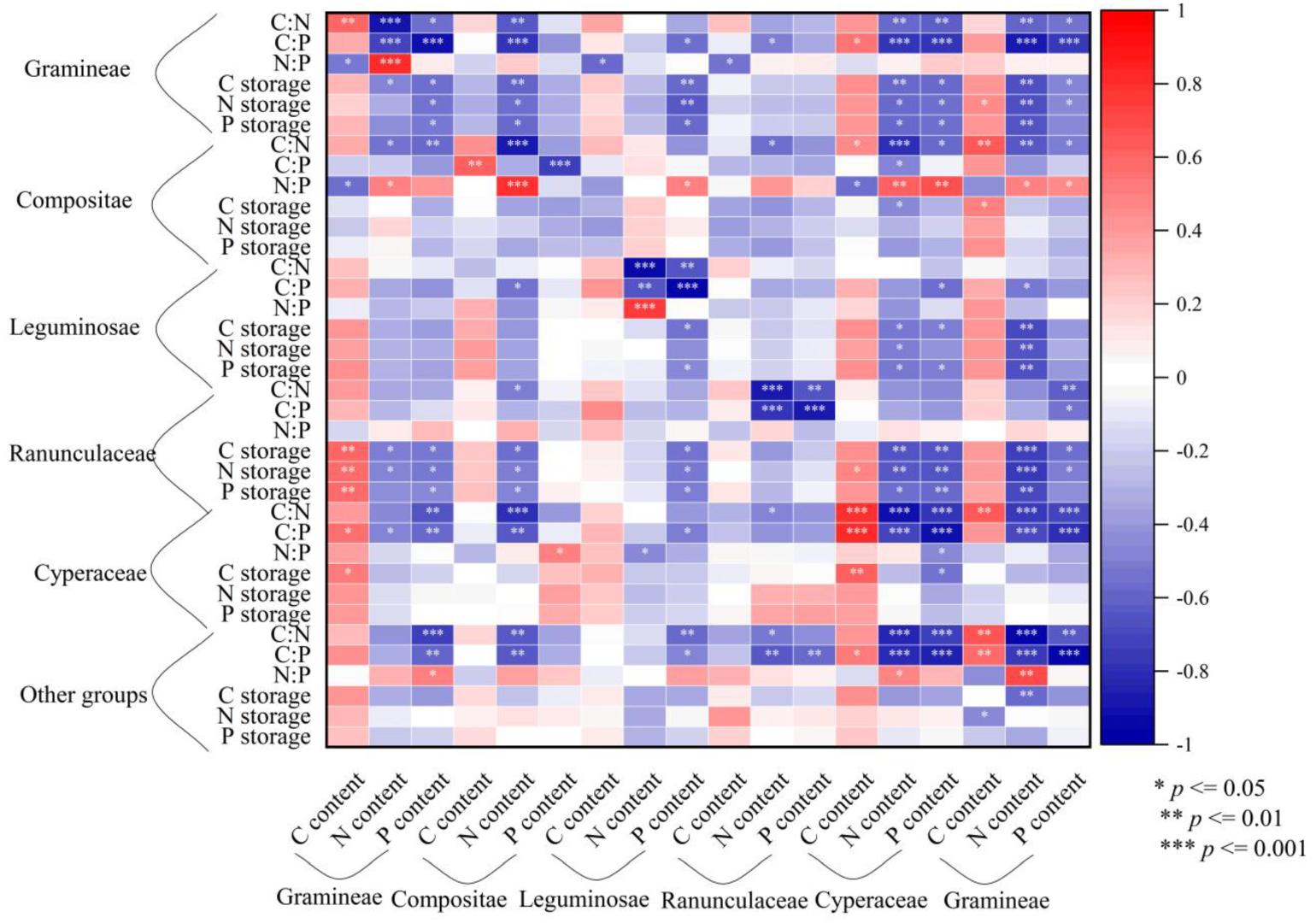
3.4. Correlation between Nutrient Content, Stoichiometry and Storage in Plant Functional Groups
4. Discussion
4.1. Effects of Grazing on the Nutrient Content of Different Plant Functional Groups
4.2. Effects of Grazing on the Stoichiometry of Different Plant Functional Groups
4.3. Effects of Grazing on the Nutrient Storage of Different Plant Functional Groups
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reed, M.S.; Stringer, L.C. Land degradation, desertification and climate change: Anticipating, assessing and adapting to future change. Taylor Fr. 2016, 10, 4324. [Google Scholar] [CrossRef]
- Olofsson, J.; Kitti, H.; Rautiainen, P.; Stark, S.; Oksanen, L. Effects of summer grazing by reindeer on composition of vegetation, productivity and nitrogen cycling. Ecography 2001, 24, 13–24. [Google Scholar] [CrossRef]
- Wang, S.P.; Duan, J.C.; Xu, G.P.; Wang, Y.F.; Zhang, Z.H.; Rui, Y.C.; Luo, C.Y.; Xu, B.B.; Zhu, X.X.; Chang, X.F.; et al. Effects of warming and grazing on soil N availability, species composition, and ANPP in an alpine meadow. Ecology 2012, 93, 2365–2376. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Zhou, G.; Zhang, F. Effects of different grazing intensities on grassland production in China: A meta-analysis. PLoS ONE 2013, 8, e81466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, N.; Kan, H.M.; Yang, G.W.; Zhang, Y.J. Changes in plant, soil, and microbes in a typical steppe from simulated grazing: Explaining potential change in soil C. Ecol. Monogr. 2015, 85, 269–286. [Google Scholar] [CrossRef]
- Sterner, R.W.; Elser, J.J.; Vitousek, P. Ecological stoichiometry: The biology of elements from molecules to the biosphere. Princet. Univ. Press 2017, 2, 15. [Google Scholar] [CrossRef] [Green Version]
- Zhou, G.Y.; Zhou, X.H.; He, Y.G.; Shao, J.J.; Hu, Z.H.; Liu, R.Q.; Zhou, H.M.; Hosseinibai, S. Grazing intensity significantly affects belowground carbon and nitrogen cycling in grassland ecosystems: A meta-analysis. Global Chang. Biol. 2017, 23, 1167–1179. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.P.; Zhang, W.P.; Yu, Y.C.; Yu, S.B.; Lambers, A.H.; Li, L. Linking shifts in species composition induced by grazing with root traits for phosphorus acquisition in a typical steppe in Inner Mongolia. Sci. Total Environ. 2020, 712, 136495. [Google Scholar] [CrossRef]
- Li, W.; Cao, W.X.; Wang, J.L.; Li, X.L.; Xu, C.L.; Shi, S.L. Effects of grazing regime on vegetation structure, productivity, soil quality, carbon and nitrogen storage of alpine meadow on the Qinghai-Tibetan Plateau. Ecol. Eng. 2017, 98, 123–133. [Google Scholar] [CrossRef]
- Stark, S.; Männistö, M.K.; Eskelinen, A. When do grazers accelerate or decelerate soil carbon and nitrogen cycling in tundra? A test of theory on grazing effects in fertile and infertile habitats. Oikos 2015, 124, 593–602. [Google Scholar] [CrossRef]
- Yang, X.J.; Huang, Z.Y.; Zhang, K.L.; Cornelissen, J.H.C.; Nardoto, G.B. C:N:P stoichiometry of Artemisia species and close relatives across northern China: Unravelling effects of climate, soil and taxonomy. J. Ecol. 2015, 103, 1020–1031. [Google Scholar] [CrossRef]
- Zheng, S.X.; Ren, H.Y.; Li, W.H.; Lan, Z.C.; Kytöviita, M.M. Scale-Dependent Effects of Grazing on Plant C: N: P Stoichiometry and Linkages to Ecosystem Functioning in the Inner Mongolia Grassland. PLoS ONE 2012, 7, e51750. [Google Scholar] [CrossRef] [PubMed]
- He, N.P.; Zhang, Y.H.; Yu, Q.; Cheng, Q.S.; Pan, Q.M.; Zhang, G.M.; Han, X.G. Grazing intensity impacts soil carbon and nitrogen storage of continental steppe. Ecosphere 2011, 2, art8. [Google Scholar] [CrossRef]
- Gusewell, S.; Koerselman, W.; Verhoeven, J.T.A. Biomass N: P ratios as indicators of nutrient limitation for plant populations in wetlands. Ecol. Appl. 2003, 13, 372–384. [Google Scholar] [CrossRef]
- Zeng, Y.L.; Fang, X.; Xiang, W.H.; Deng, X.W.; Peng, C.H. Stoichiometric and nutrient resorption characteristics of dominant tree species in subtropical Chinese forests. Ecol. Evol. 2017, 3, 3527. [Google Scholar] [CrossRef] [Green Version]
- Han, G.D.; Hao, X.Y.; Zhao, M.L.; Wang, M.J.; Ellert, B.H.; Willms, W.; Wang, M.J. Effect of grazing intensity on carbon and nitrogen in soil and vegetation in a meadow steppe in Inner Mongolia. Agric. Ecosyst. Environ. 2008, 125, 21–32. [Google Scholar] [CrossRef]
- Rui, Y.C.; Wang, S.P.; Xu, Z.H.; Wang, Y.F.; Chen, C.G.; Zhou, X.Q.; Kang, X.M.; Lu, S.B.; Hu, Y.G.; Lin, Q.Y.; et al. Warming and grazing affect soil labile carbon and nitrogen pools differently in an alpine meadow of the Qinghai–Tibet Plateau in China. J. Soils Sediments 2011, 11, 903–914. [Google Scholar] [CrossRef]
- Cease, A.J.; Elser, J.J.; Ford, C.F.; Hao, S.; Kang, L.; Harrison, J.F. Heavy livestock grazing promotes locust outbreaks by lowering plant nitrogen content. Science 2012, 335, 467–469. [Google Scholar] [CrossRef]
- Douglas, A.F. Ungulate and topographic control of nitrogen: Phosphorus stoichiometry in a temperate grassland; soils, plants and mineralization rates. Oikos 2008, 117, 591–601. [Google Scholar] [CrossRef]
- Wang, M.; Zhang, Y.; Li, R.Q.; Cao, J.; Xin, X.P.; Zhu, X.Y.; Yan, R.R. Response of carbon, nitrogen and phosphorus stoichiometry and storage of temperate meadow plants to grazing intensity. Soil Fert. Sci. China. 2022, 22, 201–212. [Google Scholar]
- Zhang, W.Y.; Fan, J.W.; Zhong, H.P.; Hu, Z.M.; Song, L.L.; Wang, N. Nitrogen and phosphorus stoichiometry of dominant plant functional groups in typical grasslands in China. Acta Agrestia Sin. 2010, 18, 503–509. [Google Scholar] [CrossRef]
- He, J.S.; Wang, L.; Flynn, D.F.B.; Wang, X.P.; Ma, W.H.; Fang, J.Y. Leaf nitrogen: Phosphorus stoichiometry across Chinese grassland biomes. Ecol. Lett. 2008, 155, 301–310. [Google Scholar] [CrossRef]
- Cao, J.; Yan, R.R.; Chen, X.Y.; Wang, X.; Yu, Q.; Zhang, Y.L.; Ning, C.; Hou, L.L.; Zhang, Y.J.; Xin, X.P. Grazing Affects the Ecological Stoichiometry of the Plant–Soil–Microbe System on the Hulunber Steppe, China. Sustainability 2019, 11, 5226. [Google Scholar] [CrossRef] [Green Version]
- Heyburn, J.; McKenzie, P.; Crawley, M.J.; Fornara, D.A. Effects of grassland management on plant C:N:P stoichiometry: Implications for soil element cycling and storage. Ecosphere 2017, 8, e01963. [Google Scholar] [CrossRef]
- Bai, Y.F.; Wu, J.G.; Clark, C.M.; Pan, Q.M.; Zhang, L.X.; Chen, S.P.; Wang, Q.B.; Han, X.G.; Brian, W. Grazing alters ecosystem functioning and C:N:P stoichiometry of grasslands along a regional precipitation gradient. J. Appl. Ecol. 2012, 49, 1204–1215. [Google Scholar] [CrossRef]
- Toohy, P.; Fenton, O.; Holden, N.M.; Humphereys, J. The effects of treading by two breeds of dairy cow with different live weights on soil physical properties, poaching damae and herbage production on a poorly drained clay-loam soil. J. Agric. Sci. 2015, 153, 1424–1436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.P.; Li, Y.Q.; Zhao, X.Y.; Awada, T.; Shang, W.; Han, J.J. Effects of grazing exclusion on soil properties and on ecosystem carbon and nitrogen storage in a sandy rangeland of Inner Mongolia, Northern China. Environ. Manag. 2012, 50, 622–632. [Google Scholar] [CrossRef]
- Yan, R.R.; Tang, H.J.; Xin, X.P.; Chen, B.R.; Murray, P.J.; Yan, Y.C.; Wang, X.; Yang, G.X. Grazing intensity and driving factors affect soil nitrous oxide fluxes during the growing seasons in the Hulunber meadow steppe of China. Environ. Res. Lett. 2016, 11, 054004. [Google Scholar] [CrossRef] [Green Version]
- He, J.S.; Fang, J.Y.; Wang, Z.H.; Guo, D.L.; Flynn, D.F.B.; Geng, Z. Stoichiometry and Large-Scale Patterns of Leaf Carbon and Nitrogen in the Grassland Biomes of China. Oecologia 2006, 149, 115–122. [Google Scholar] [CrossRef]
- Li, X.l.; Liu, Z.; Ren, W.B.; Ding, Y.; Ji, L.; Guo, F.H.; Hou, X.Y. Linking nutrient strategies with plant size along a grazing gradient: Evidence from Leymus chinensis in a natural pasture. J. Integr. Agric. 2016, 15, 1132–1144. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.X.; Zhu, K.J. Nitrogen and phosphorus stoichiometry of different functional groups of alpine meadows on the eastern edge of the Qinghai-Tibet Plateau. Chin. J. Grassland 2013, 35, 52–58. [Google Scholar]
- Jiang, L.L.; Wang, S.P.; Pang, Z.; Wang, C.S.; Kardol, P.; Zhou, X.Q.; Rui, Y.C.; Chun, L.Z.; Wang, Y.F.; Xu, X.L. Grazing modifies inorganic and organic nitrogen uptake by coexisting plant species in alpine grassland. Biol. Fert. Soils. 2016, 52, 211–221. [Google Scholar] [CrossRef]
- Yang, Z.A.; Zhu, Q.A.; Zhan, W.; Xu, Y.Y.; Zhu, E.X.; Gao, Y.H.; Li, S.Q.; Zheng, Q.Y.; Zhu, D.; He, Y.X.; et al. The linkage between vegetation and soil nutrients and their variation under different grazing intensities in an alpine meadow on the eastern Qinghai-Tibetan Plateau. Ecol. Eng. 2018, 110, 128–136. [Google Scholar] [CrossRef]
- Baron, V.S.; Dick, A.C.; Mapfumo, E.; MALHI, S.S.; Naeth, M.A.; Chanasyk, D.S. Grazing impacts on soil nitrogen and phosphorus under Parkland pastures. J. Range Manag. 2001, 54, 704–710. [Google Scholar] [CrossRef]
- Risch, A.C.; Schutz, M.; Vandegehuchte, M.L.; Van, D.P.W.H.; Duyts, H.; Raschein, U.; Gwiazdowicz, D.J.; Busse, M.D.; Page, D.D.S.; Zimmermann, S. Aboveground vertebrate and invertebrate herbivore impact on net N mineralization in subalpine grasslands. Ecology 2015, 96, 3312–3322. [Google Scholar] [CrossRef] [Green Version]
- Faghihinia, M.; Zou, Y.; Chen, Z.; Bai, Y.F.; Li, W.H.; Marrs, R.; Staddon, P. The response of grassland mycorrhizal fungal abundance to a range of long-term grazing intensities. Rhizosphereneth 2020, 13, 100178. [Google Scholar] [CrossRef]
- Li, Y.J.; Zhu, Y.; Zhao, J.N.; Li, G.; Wang, H.; Lai, X.; Yang, D.L. Effects of Rest Grazing on Organic Carbon Storage in Stipa grandis Steppe in Inner Mongolia, China. J. Integr. Agric. 2014, 13, 624–634. [Google Scholar] [CrossRef]
- Scogings, P.F.; Hjältén, J.; Skarpe, C. Does large herbivore removal affect secondary metabolites, nutrients and shoot length in woody species in semi-arid savannas? J. Arid. Environ. 2013, 88, 4–8. [Google Scholar] [CrossRef] [Green Version]
- Dybzinski, R.; Fargione, J.E.; Zak, D.R.; Fornara, D.; Tilman, D. Soil fertility increases with plant species diversity in a long-term biodiversity experiment. Oecologia 2008, 158, 85–93. [Google Scholar] [CrossRef]
- Reich, P.B.; Oleksyn, J. Global patterns of plant leaf N and P in relation to temperature and latitude. Proc. Natl. Acad. Sci. USA 2004, 101, 11001–11006. [Google Scholar] [CrossRef] [Green Version]
- Song, S.S.; Zhu, J.L.; Zheng, T.L.; Tang, Z.Y.; Zhang, F.; Ji, C.J.; Shen, Z.H.; Zhu, J.X. Long-term grazing exclusion reduces species diversity but increases community heterogeneity in an alpine grassland. Front. Ecol. Evol. 2020, 8, 66. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.Y.; Zhang, W.Q.; Li, J.X.; Zhao, L.M.; Ulan, T.T.; Wan, Z.Q. Response of carbon and nitrogen contents of soil and dominant plants to grazing intensity in steppe of Stipa krylovii. Acta Agrestia Sin. 2022, 30, 2255–2263. [Google Scholar] [CrossRef]
- Stefan, L.; Michael, B.E.; Amir, M. Effects of grazing control on ecosystem recovery, biological productivity gains, and soil carbon sequestration in long-term degraded loess farmlands in the Northern Negev, Israel. Land Degrad. Dev. 2021, 32, 2580–2594. [Google Scholar] [CrossRef]
- Veldhuis, M.P.; Gommers, M.I.; Olff, H.; Berg, M.P. Spatial redistribution of nutrients by large herbivores and dung beetles in a savanna ecosystem. J. Ecol. 2018, 106, 422–433. [Google Scholar] [CrossRef] [Green Version]
- Poorter, H.; Niklas, K.J.; Reich, P.B.; Oleksyn, J.; Poot, P.; Mommer, L. Biomass allocation to leaves, stems and roots: Meta-analyses of interspecific variation and environmental control. New Phytol. 2012, 193, 30–50. [Google Scholar] [CrossRef] [PubMed]
- Desurmont, G.A.; Agrawal, A.A. Do plant defenses predict damage by an invasive herbivore? A comparative study of the viburnum leaf beetle. Ecol. Appl. 2014, 24, 759–769. [Google Scholar] [CrossRef] [PubMed]
- Hedin, L.O. Global organization of terrestrial plant nutrient interactions. Proc. Natl. Acad. Sci. USA 2004, 101, 10849–10850. [Google Scholar] [CrossRef] [Green Version]
- Dong, X.Y.; Fu, H.; Li, X.D.; Niu, D.C.; Guo, D.; Li, X.D. Effects of grazing and enclosure on biomass and carbon, nitrogen and phosphorus storage of typical steppe plants on the Loess Plateau. Acta Prata. Sin. 2010, 19, 175–184. [Google Scholar] [CrossRef]
- Song, S.S.; Zhang, J.S.; Zheng, T.L.; Zhang, F.; Zhu, J.X. Effects of fence closure on carbon storage in alpine meadow vegetation in Qinghai-Hebei. Prata. Sci. 2020, 37, 2414–2421. [Google Scholar] [CrossRef]
- Wang, S.L.; Hou, F.J. Short-term study on the yak dung seed bank on the Qinghai-Tibetan Plateau: Effects of grazing season, seed characteristics and forage preferences. Plant Soil 2021, 465, 367–383. [Google Scholar] [CrossRef]
- Fan, Y.J.; Hou, X.Y.; Shi, H.X.; Shi, S.L. Effects of grazing and fencing on carbon and nitrogen reserves in plants and soils of alpine meadow in the three headwater resource regions. Russ. J. Ecol. 2013, 44, 80–88. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Grazing Intensity | Gramineae | Compositae | Leguminosae | Ranunculaceae | Cyperaceae | Other Groups |
|---|---|---|---|---|---|---|
| G0.00 | Leymus chinensis, Achnatherum sibiricum Keng, Stipa baicalensis | Artemisia laciniata Linn, Artemisia dracunculu L., Serratula centauroides Cass | Pulsatilla turczaninovii Kryl, Thalictrum squarrosum Steph | Carex duriuscula L., Carex pediformis | Astragalus melilotoides Pall, Vicia amoena Fisch | Schizonepeta multifida Linn, Galium verum Linn, Potentilla bifurca |
| G0.23 | Leymus chinensis, Achnatherum sibiricum, Stipa baicalensis | Artemisia laciniata, Artemisia frigida, Heteropappus altaicus Novopokr | Pulsatilla turczaninovii, Thalictrum squarrosum | Carex duriuscula, Carex pediformis | Astragalus adsurgens Pall, Astragalus melilotoides, Vicia amoena | Schizonepeta multifida, Galium verum, Potentilla bifurca |
| G0.34 | Leymus chinensis, Achnatherum sibiricum, Stipa baicalensis | Artemisia laciniata, Artemisia dracunculus, Artemisia frigida | Pulsatilla turczaninovii, Thalictrum squarrosum | Carex duriuscula, Carex pediformis | Astragalus adsurgens, Astragalus melilotoides, Vicia amoena | Schizonepeta multifida, Galium verum, Potentilla bifurca |
| G0.46 | Leymus chinensis, Koeleria macrantha Schult, Cleistogenes squarrosa | Artemisia laciniata, Artemisia frigida | Pulsatilla turczaninovii, Thalictrum squarrosum | Carex duriuscula, Carex pediformis | Oxytropis myriophylla Pall, Astragalus adsurgens, Vicia amoena | Schizonepeta multifida, Galium verum, Potentilla tanacetifolia Willd |
| G0.69 | Koeleria macrantha, Cleistogenes squarrosa | Artemisia laciniata, Artemisia frigida | Pulsatilla turczaninovii, Thalictrum squarrosum | Carex duriuscula, Carex pediformis | Medicago falcata L., Oxytropis myriophylla, Astragalus adsurgens | Schizonepeta multifida, Potentilla acaulis L., Potentilla bifurca |
| G0.92 | Koeleria macrantha, Cleistogenes squarrosa | Artemisia laciniata, Artemisia frigida | Pulsatilla turczaninovii, Thalictrum squarrosum, Thalictrum petaloideum L | Carex duriuscula | Oxytropis myriophylla, Astragalus adsurgens | Schizonepeta multifida, Potentilla acaulis, Gentiana squarrosa Ledeb |
| Grazing Intensity | G0.00 | G0.23 | G0.34 | G0.46 | G0.69 | G0.92 | |
|---|---|---|---|---|---|---|---|
| Gramineae | biomass (g/m2) | 164.41 ± 53.69 a | 135.91 ± 27.20 ab | 127.58 ± 16.78 ab | 61.69 ± 14.44 bc | 22.66 ± 7.32 c | 17.08 ± 3.62 c |
| Proportion (%) | 45.95 | 41.67 | 43.08 | 24.83 | 15.44 | 17.31 | |
| Compositae | Biomass (g/m2) | 27.72 ± 1.55 b | 49.53 ± 12.11 ab | 58.83 ± 12.95 ab | 65.05 ± 12.77 a | 43.18 ± 11.07 ab | 39.07 ± 9.43 ab |
| Proportion (%) | 8.27 | 15.24 | 20.38 | 27.42 | 30.59 | 38.72 | |
| Leguminosae | Biomass (g/m2) | 14.37 ± 3.05 a | 10.08 ± 2.09 ab | 5.87 ± 0.79 bc | 4.91 ± 1.67 bc | 3.97 ± 1.84 bc | 1.19 ± 0.20 c |
| Proportion (%) | 4.44 | 3.14 | 2.04 | 2.11 | 2.76 | 1.21 | |
| Ranunculaceae | Biomass (g/m2) | 42.20 ± 13.64 a | 39.45 ± 1.83 a | 28.96 ± 0.76 a | 28.84 ± 5.53 a | 7.63 ± 2.25 b | 3.46 ± 1.63 b |
| Proportion (%) | 13.31 | 12.28 | 9.92 | 11.68 | 5.23 | 3.3 | |
| Cyperaceae | biomass (g/m2) | 53.19 ± 30.28 a | 43.20 ± 18.68 a | 44.90 ± 6.93 a | 44.91 ± 2.23 a | 38.20 ± 14.39 a | 10.14 ± 2.92 a |
| Proportion (%) | 14.79 | 13.78 | 15.28 | 18.5 | 28.17 | 10.13 | |
| other groups | biomass (g/m2) | 45.78 ± 8.74 a | 44.61 ± 2.74 a | 27.88 ± 6.00 a | 37.80 ± 8.87 a | 24.88 ± 2.44 a | 27.26 ± 6.93 a |
| Proportion (%) | 13.24 | 13.88 | 9.3 | 15.46 | 17.81 | 29.34 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, M.; Zhang, C.; Chen, S.; Zhang, Y.; Li, Y.; Xin, X.; Wang, X.; Yan, R. Effects of Grazing Intensity on the Carbon, Nitrogen and Phosphorus Content, Stoichiometry and Storage of Plant Functional Groups in a Meadow Steppe. Agronomy 2022, 12, 3057. https://doi.org/10.3390/agronomy12123057
Wang M, Zhang C, Chen S, Zhang Y, Li Y, Xin X, Wang X, Yan R. Effects of Grazing Intensity on the Carbon, Nitrogen and Phosphorus Content, Stoichiometry and Storage of Plant Functional Groups in a Meadow Steppe. Agronomy. 2022; 12(12):3057. https://doi.org/10.3390/agronomy12123057
Chicago/Turabian StyleWang, Miao, Chu Zhang, Sisi Chen, Yu Zhang, Yingxin Li, Xiaoping Xin, Xiaoli Wang, and Ruirui Yan. 2022. "Effects of Grazing Intensity on the Carbon, Nitrogen and Phosphorus Content, Stoichiometry and Storage of Plant Functional Groups in a Meadow Steppe" Agronomy 12, no. 12: 3057. https://doi.org/10.3390/agronomy12123057