
Salicylic Acid Pre-Treatment Reduces the Physiological Damage Caused by the Herbicide Mesosulfuron-methyl + Iodosulfuron-methyl in Wheat (Triticum aestivum)



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Treatments
2.2. Antioxidant Analyses
2.2.1. Malondialdehyde and Hydrogen Peroxide Content
2.2.2. Extraction of Leaf Tissue for Determination of Enzyme Activity
2.2.3. Ascorbate Peroxidase and Superoxide Dismutase Assays
2.2.4. Peroxidase and Catalase Assays
2.2.5. Glutathione-S-Transferase and Polyphenol Oxidase Assays
2.2.6. Proline Content
2.3. Statistical Analysis
3. Results
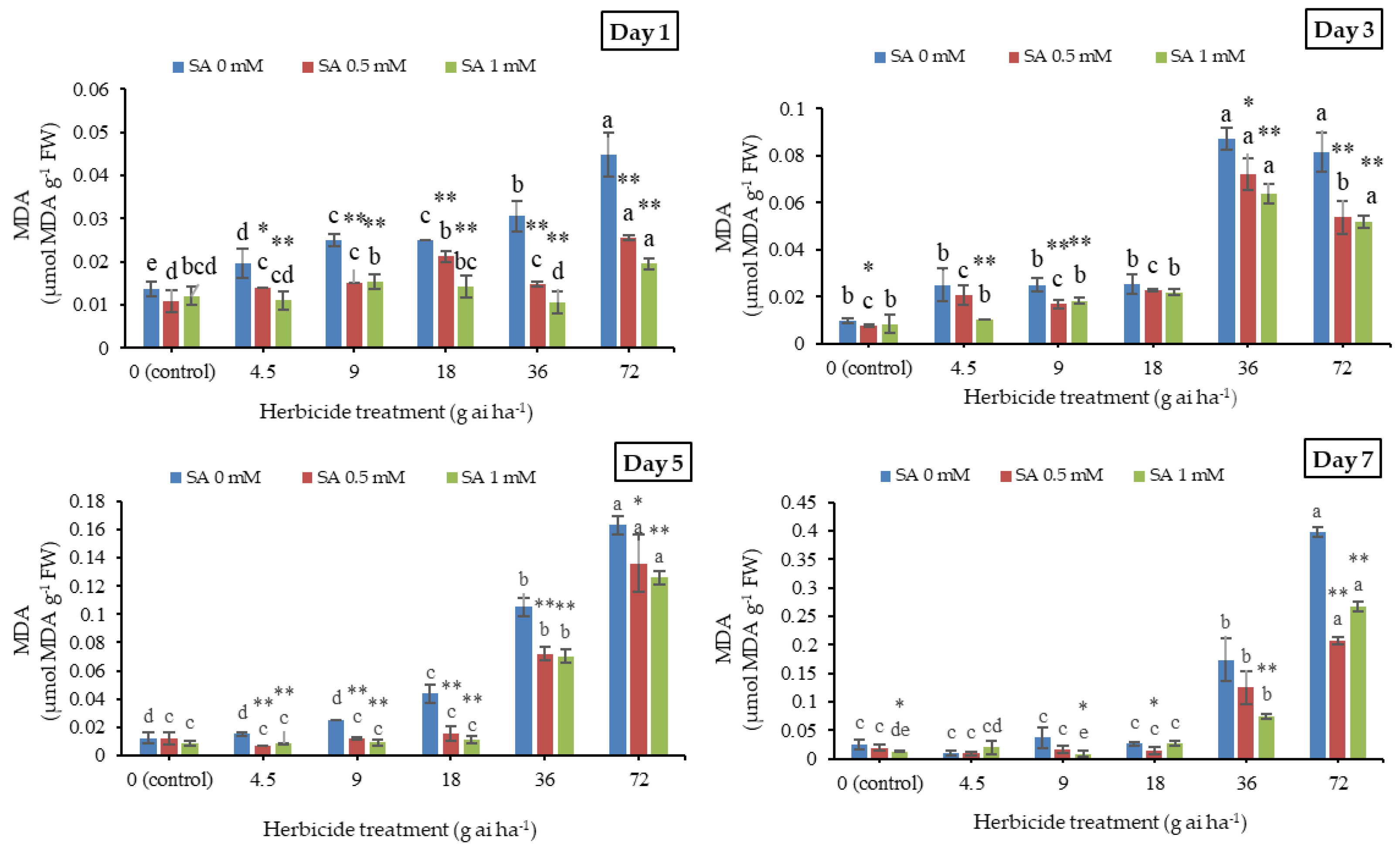
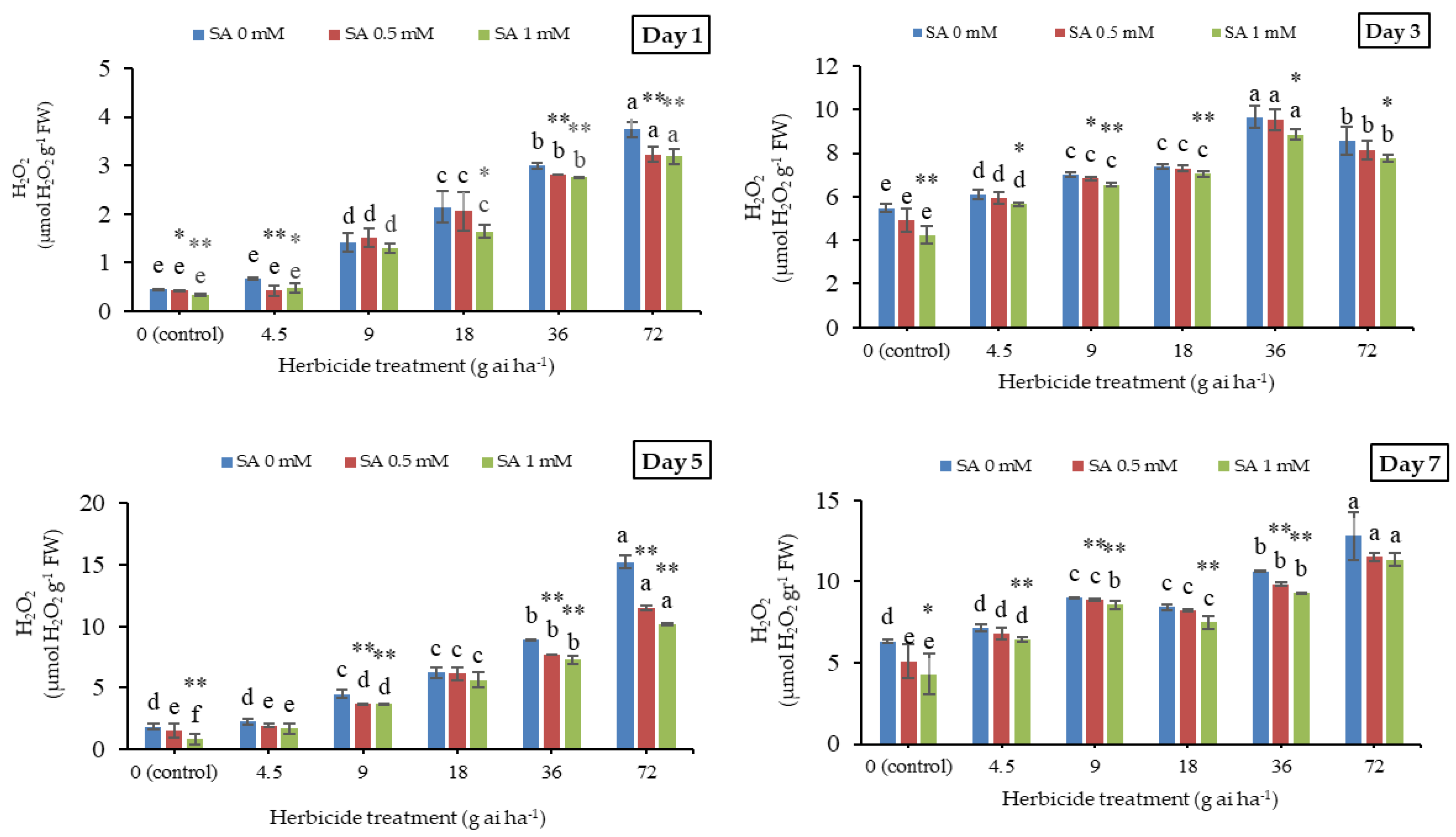
3.1. Malondialdehyde and Hydrogen Peroxide Contents
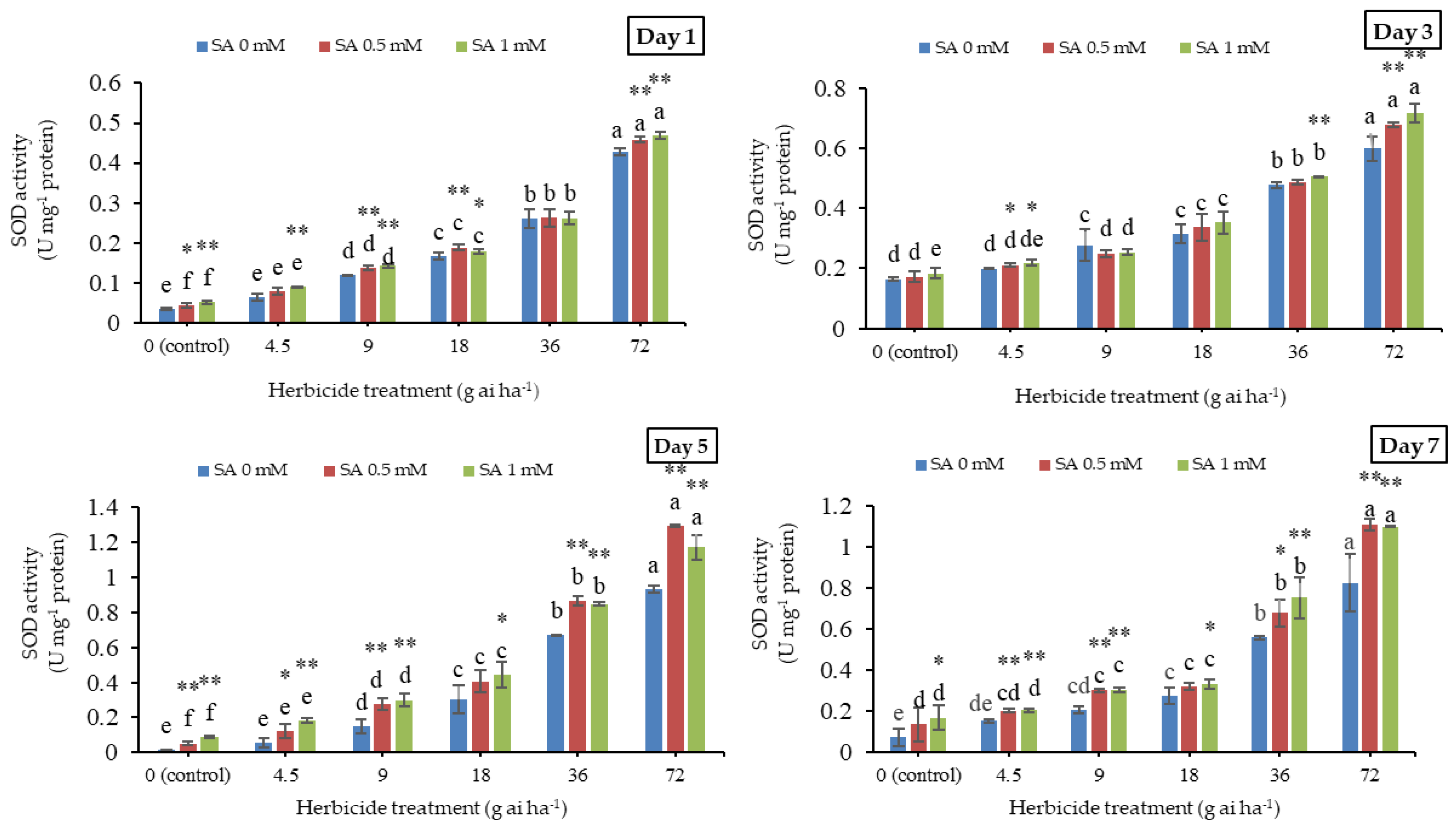
3.2. Superoxide Dismutase Activity
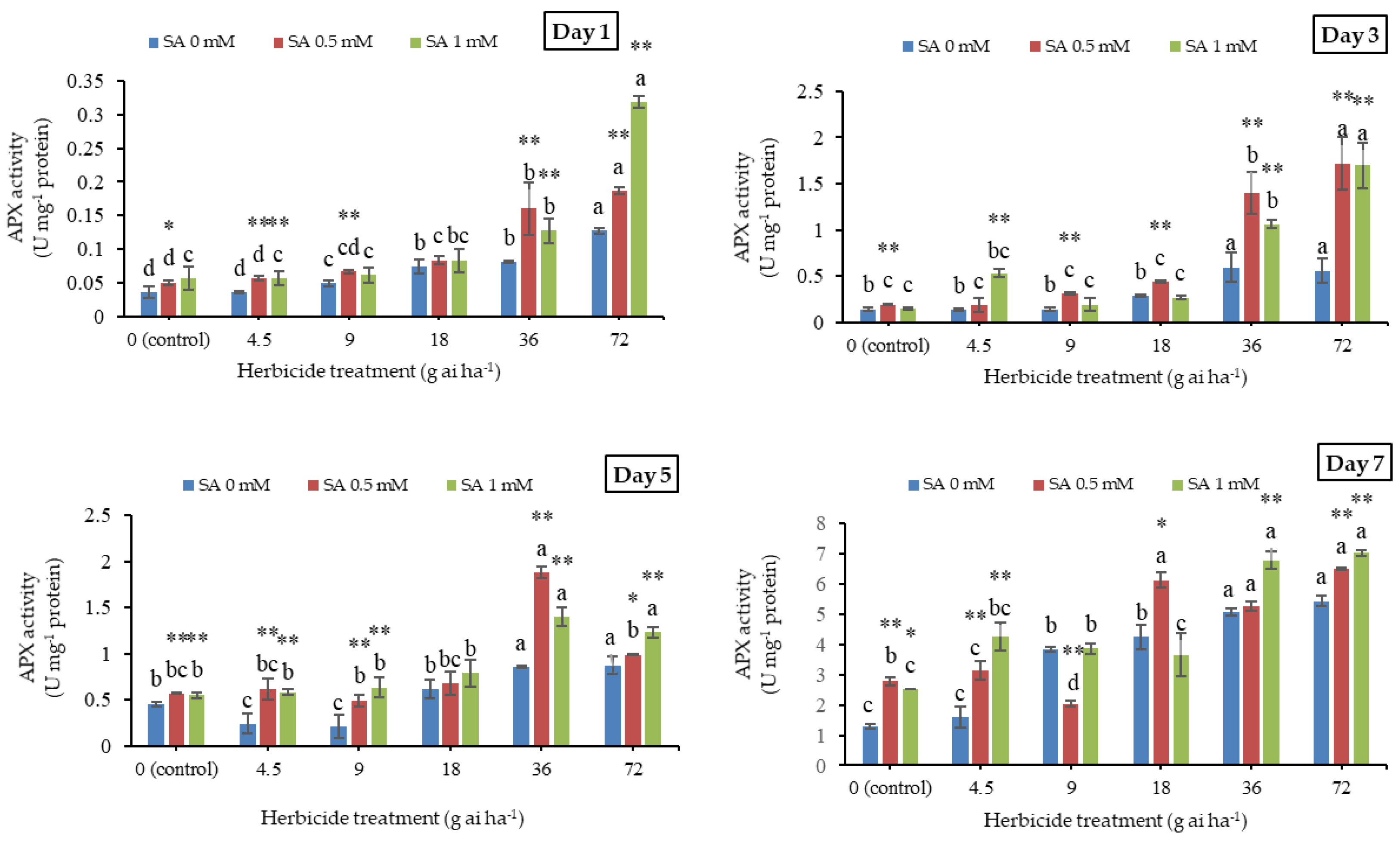
3.3. Ascorbate Peroxidase Activity
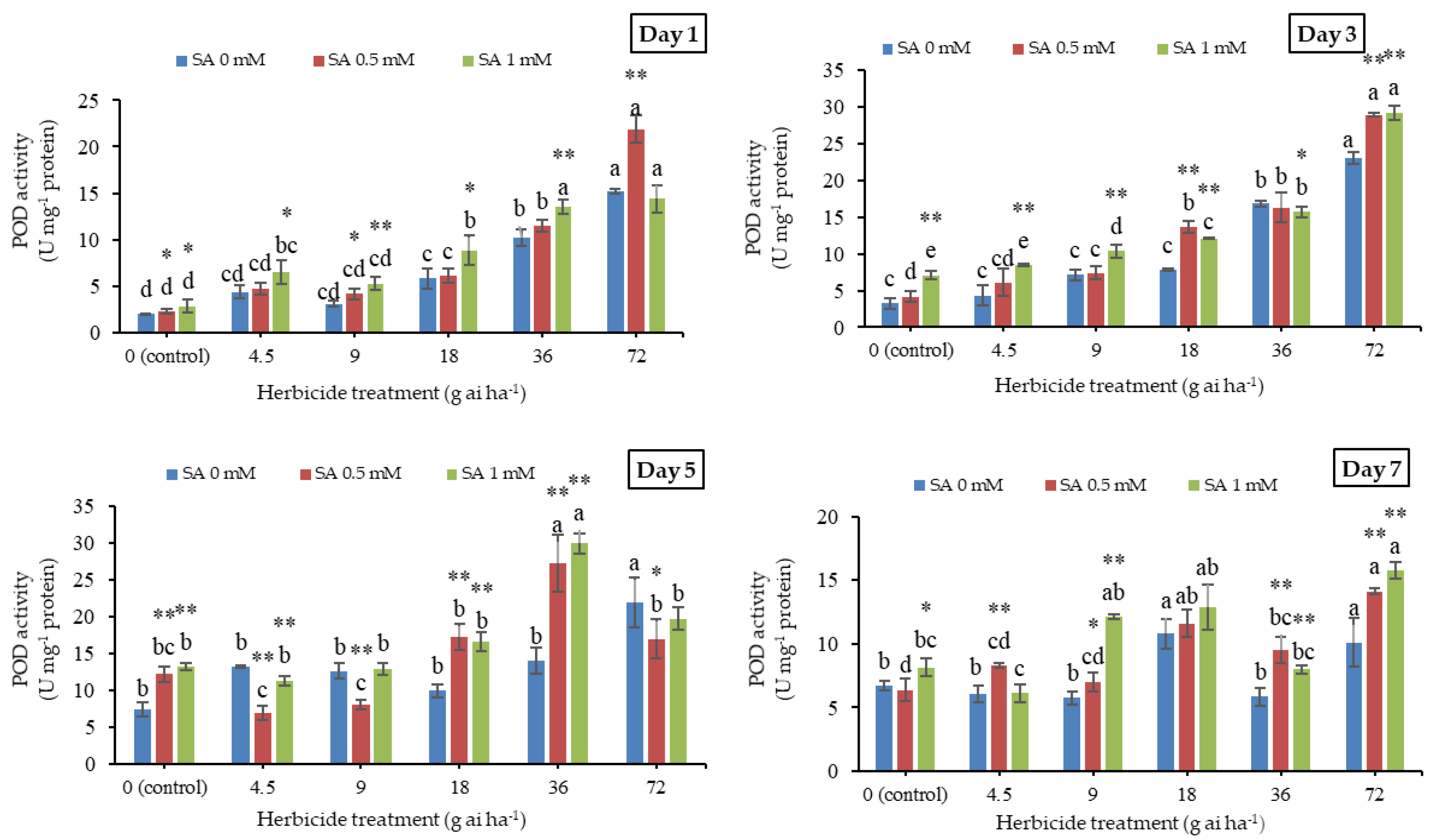
3.4. Peroxidase Activity
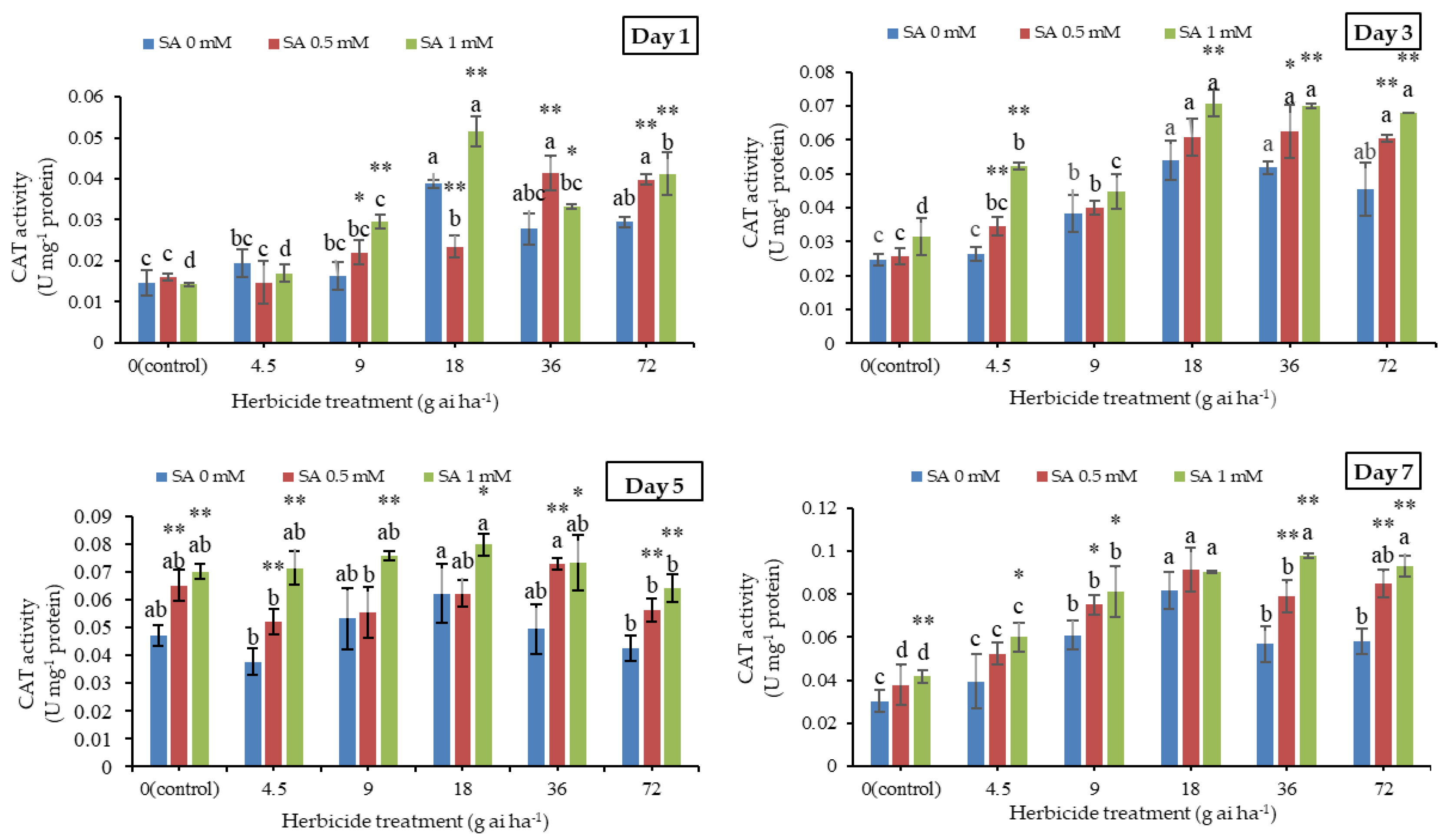
3.5. Catalase Activity
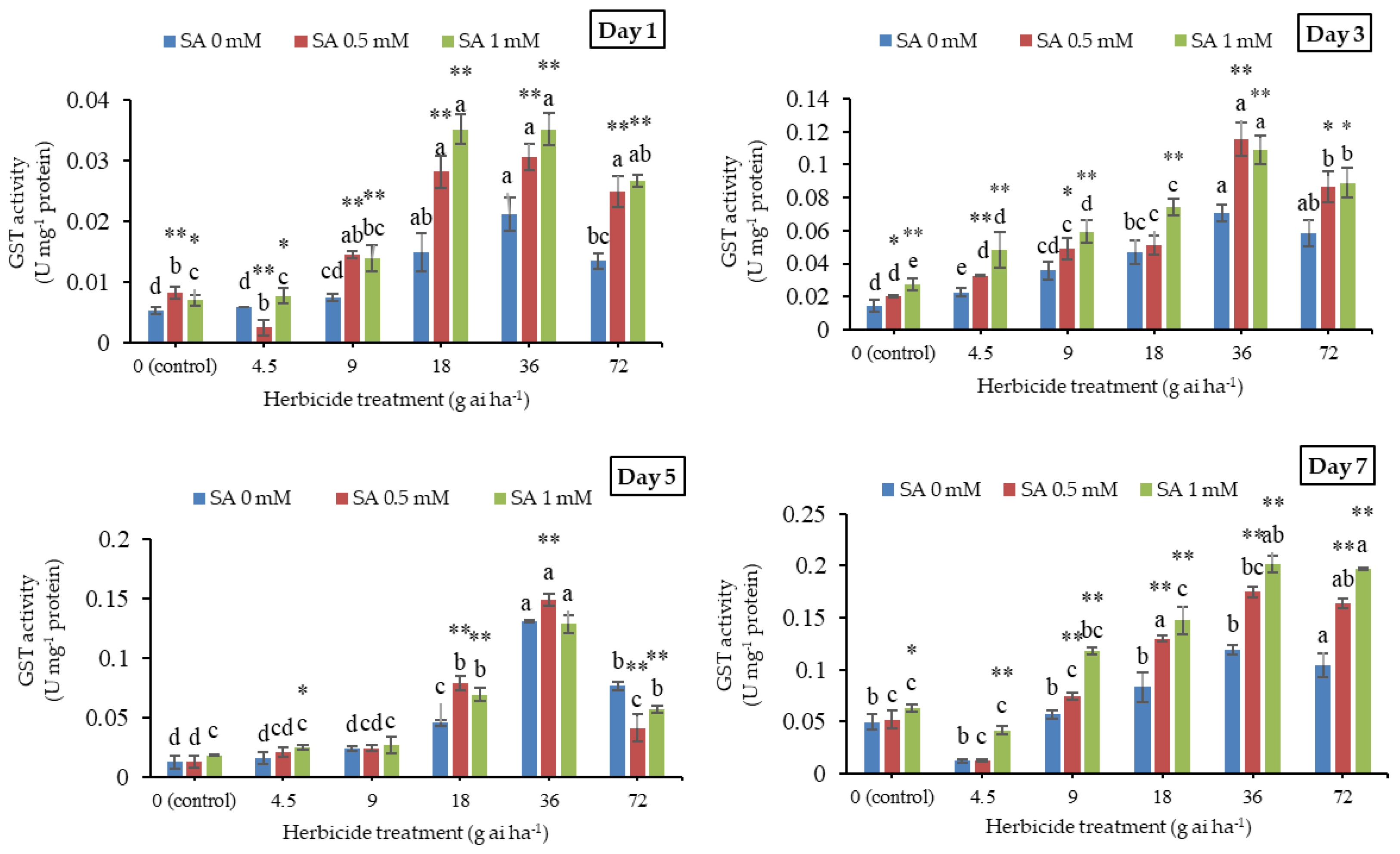
3.6. Glutathione S-Transferase Activity
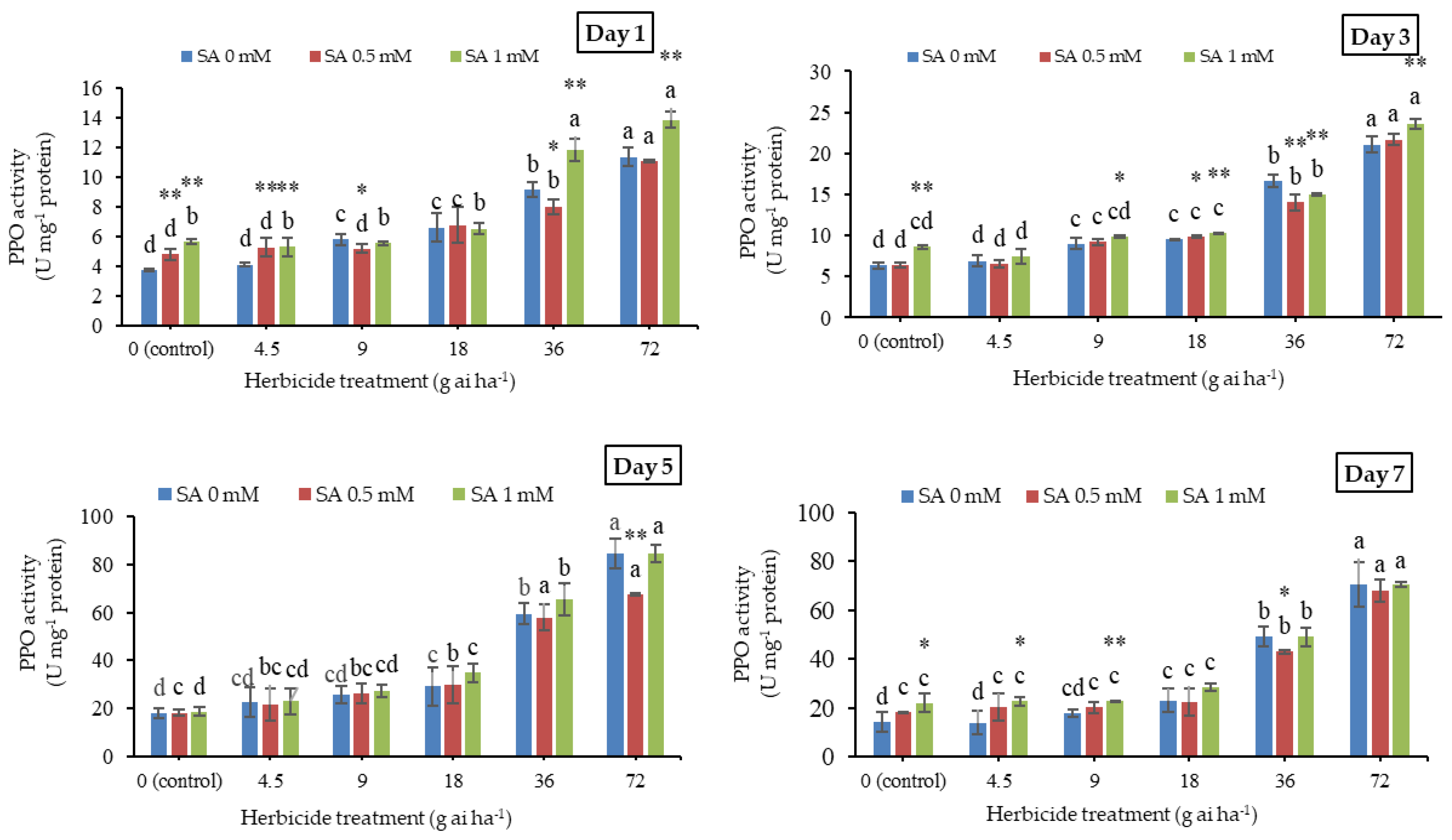
3.7. Polyphenol Oxidase Activity
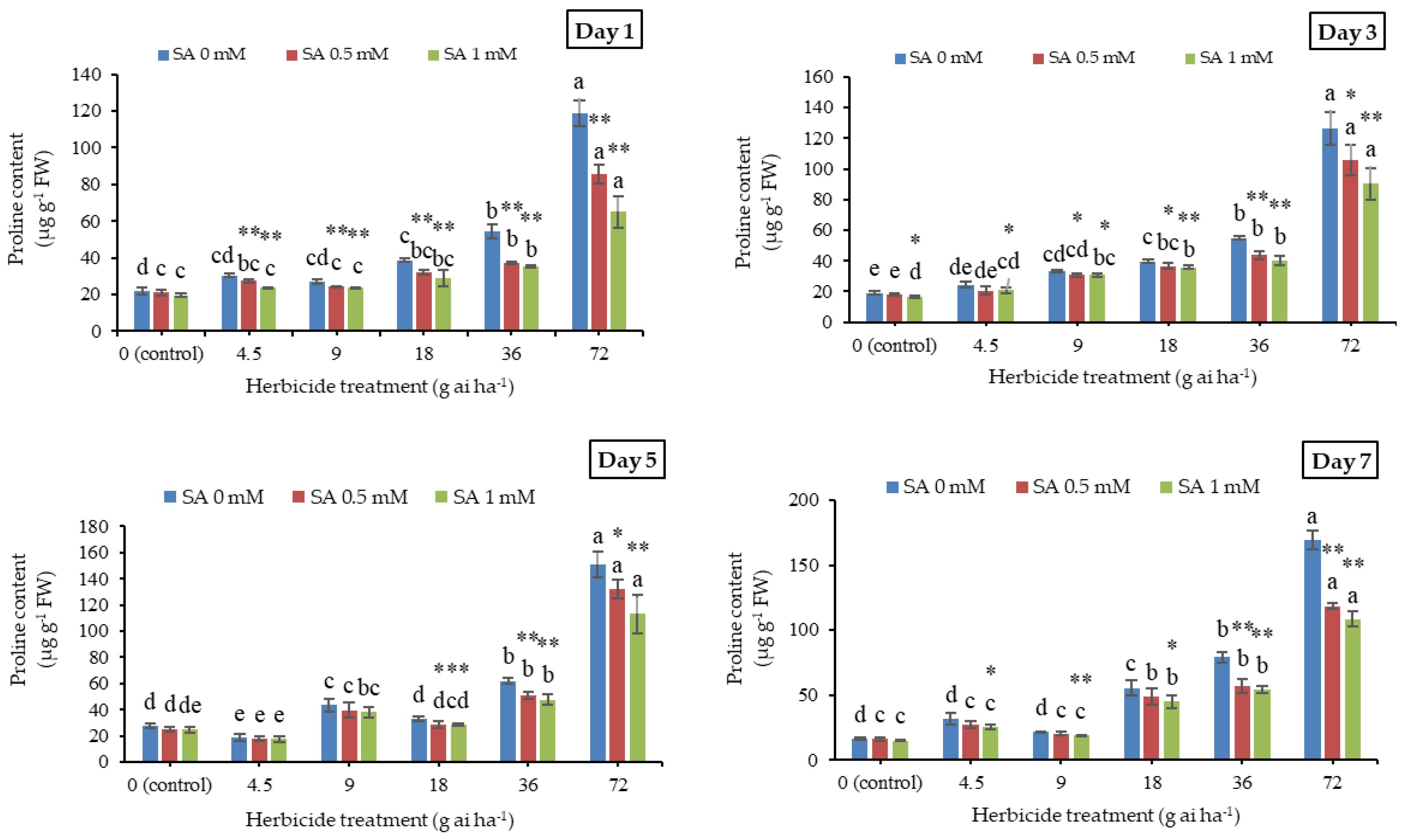
3.8. Proline Content
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- He, H.; He, L. The role of carbon monoxide signaling in the responses of plants to abiotic stresses. Nitric Oxide 2014, 42, 40–43. [Google Scholar] [CrossRef] [PubMed]
- Kosová, K.; Vítámvás, P.; Prášil, I.T.; Renaut, J. Plant proteome changes under abiotic stress—Contribution of proteomics studies to understanding plant stress response. J. Proteom. 2011, 74, 1301–1322. [Google Scholar] [CrossRef] [PubMed]
- Kumar, L.; Singh, P.K.; Alam, M.S.; Ujjwal, A. Herbicides combinations for sustainable weed management on complex weed flora in dry and irrigated direct seeded rice: A review. J. Pharmacogn. Phytochem. 2021, 10, 2539–2544. [Google Scholar] [CrossRef]
- Khatami, S.A.; Alebrahim, M.T.; Majd, R. The effect of rimsulfuron application time and dose on weed control and potato (Solanum tuberosum) tuber yield. Iran. J. Weed Sci. 2017, 12, 185–198. [Google Scholar]
- Hess, F.D. Light-dependent herbicides: An overview. Weed Sci. 2000, 48, 160–170. [Google Scholar] [CrossRef]
- Zangoueinejad, R.; Alebrahim, M.T.; Tseng, T.M. Evaluation of auxin tolerance in selected tomato germplasm under greenhouse and field conditions. Weed Technol. 2019, 33, 815–822. [Google Scholar] [CrossRef]
- Alebrahim, M.T.; Majd, R.; Abdollahi, F.; Zangoueinejad, R.; Dayan, F.E.; Mathiassen, S.K.; Kudsk, P. Absorption and metabolism of foliar-applied rimsulfuron in potato (Solanum tuberosum L.), common lambsquarters (Chenopodium album L.) and redroot pigweed (Amaranthus retroflexus L.). Potato Res. 2021, 64, 635–648. [Google Scholar] [CrossRef]
- Abdollahi, F.; Alebrahim, M.T.; Ngov, C.; Lallemand, E.; Zheng, Y.; Villette, C.; Zumsteg, J.; André, F.; Navrot, N.; Werck-Reichhart, D.; et al. Innate promiscuity of the CYP706 family of P450 enzymes provides a suitable context for the evolution of dinitroaniline resistance in weed. New Phytol. 2021, 229, 3253–3268. [Google Scholar] [CrossRef]
- Song, N.H.; Le Yin, X.; Chen, G.F.; Yang, H. Biological responses of wheat (Triticum aestivum) Plants to the herbicide chlorotoluron in soils. Chemosphere 2007, 68, 1779–1787. [Google Scholar] [CrossRef]
- Caverzan, A.; Piasecki, C.; Chavarria, G.; Stewart, C.N.; Vargas, L. Defenses against ROS in crops and weeds: The effects of interference and herbicides. Int. J. Mol. Sci. 2019, 20, 1086. [Google Scholar] [CrossRef] [Green Version]
- Kruse, N.D.; Vidal, R.A.; Dalmaz, C.; Trezzi, M.M.; Siqueira, I. Oxidative stress in sunflower (Helianthus annuus) indicates synergism for the metribuzin clomazone combination. Planta Daninha 2006, 24, 379–390. [Google Scholar] [CrossRef]
- Prasad, S.M.; Kumar, S.; Parihar, P.; Singh, R. Interactive effects of herbicide and enhanced UV-B on growth, oxidative damage and the ascorbate-glutathione cycle in two azolla species. Ecotoxicol. Environ. Saf. 2016, 133, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Varshney, S.; Khan, M.I.R.; Masood, A.; Per, T.S. Contribution of plant growth regulators in mitigation of herbicidal stress. J. Plant Biochem. Physiol. 2015, 3, 4. [Google Scholar] [CrossRef] [Green Version]
- El-Tayeb, M.A. Response of barley grains to the interactive e.ect of salinity and salicylic acid. Plant Growth Regul. 2005, 45, 215–224. [Google Scholar] [CrossRef]
- Shah, J. The salicylic acid loop in plant defense. Curr. Opin. Plant Biol. 2003, 6, 365–371. [Google Scholar] [CrossRef]
- Klessig, D.F.; Choi, H.W.; Dempsey, D.A. Systemic acquired resistance and salicylic acid: Past, present, and future. Mol. Plant-Microbe Interact. 2018, 31, 871–888. [Google Scholar] [CrossRef] [Green Version]
- Ananieva, E.A.; Alexieva, V.S.; Popova, L.P. Treatment with salicylic acid decreases the effects of paraquat on photosynthesis. J. Plant Physiol. 2002, 159, 685–693. [Google Scholar] [CrossRef]
- Green, J.M. Review of glyphosate and ALS-Inhibiting herbicide crop resistance and resistant weed management. Weed Technol. 2007, 21, 547–558. [Google Scholar] [CrossRef]
- Tranel, P.; Wright, T.R. Resistance of weeds to ALS-Inhibiting herbicides: What have we learned? Weed Sci. 2002, 50, 700–712. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Alexieva, V.; Sergiev, I.; Mapelli, S.; Karanov, E. The effect of drought and ultraviolet radiation on growth and stress markers in pea and wheat. Plant Cell Environ. 2001, 24, 1337–1344. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar] [CrossRef]
- Chen, T.; Zhang, B. Measurements of proline and malondialdehyde content and antioxidant enzyme activities in leaves of drought stressed cotton. Bio-Protocol 2016, 6, e1913. [Google Scholar] [CrossRef] [Green Version]
- Kar, M.; Mishra, D. Catalase, Peroxidase, and polyphenoloxidase activities during rice leaf senescence. Plant Physiol. 1976, 57, 315–319. [Google Scholar] [CrossRef] [Green Version]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Peixoto, F.; Alves-Fernandes, D.; Santos, D.; Fontaínhas-Fernandes, A. Toxicological effects of oxyfluorfen on oxidative stress enzymes in tilapia Oreochromis niloticus. Pestic. Biochem. Physiol. 2006, 85, 91–96. [Google Scholar] [CrossRef]
- Jiang, L.; Yang, H. Prometryne-induced oxidative stress and impact on antioxidant enzymes in wheat. Ecotoxicol. Environ. Saf. 2009, 72, 1687–1693. [Google Scholar] [CrossRef]
- Song, N.H.; Yang, Z.M.; Zhou, L.x.; Wu, X.; Yang, H. Effect of dissolved organic matter on the toxicity of chlorotoluron to Triticum aestivum. J. Environ. Sci. 2006, 18, 101–108. [Google Scholar]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Czarnocka, W.; Karpiński, S. Friend or foe? Reactive oxygen species production, scavenging and signaling in plant response to environmental stresses. Free Radic. Biol. Med. 2018, 122, 4–20. [Google Scholar] [CrossRef]
- Pan, D.; Li, Q.X.; Lin, Z.; Chen, Z.; Tang, W.; Pan, C.; Tan, H.; Zeng, D. Interactions between salicylic acid and antioxidant enzymes tilting the balance of H2O2 from photorespiration in non-target crops under halosulfuron-methyl stress. Pestic. Biochem. Physiol. 2017, 143, 214–223. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.J.; Lu, Y.C.; Zhang, J.J.; Tan, L.R.; Yang, H. Accumulation and toxicological response of atrazine in rice crops. Ecotoxicol. Environ. Saf. 2014, 102, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Boulahia, K.; Carol, P.; Planchais, S.; Abrous-Belbachir, O. Phaseolus Vulgaris L. seedlings exposed to prometryn herbicide contaminated soil trigger an oxidative stress response. J. Agric. Food Chem. 2016, 64, 3150–3160. [Google Scholar] [CrossRef] [PubMed]
- Singh, H.; Singh, N.B.; Singh, A.; Hussain, I.; Yadav, V. Physiological and biochemical effects of salicylic acid on pisum sativum exposed to isoproturon. Arch. Agron. Soil Sci. 2016, 62, 1425–1436. [Google Scholar] [CrossRef]
- Zhang, J.; Kirkham, M.B. Drought-stress-induced changes in activities of superoxide dismutase, catalase, and peroxidase in wheat species. Plant Cell Physiol. 1994, 35, 785–791. [Google Scholar] [CrossRef]
- Kumar, S.; Trivedi, P.K. Glutathione S-Transferases: Role in combating abiotic stresses including arsenic detoxification in plants. Front. Plant Sci. 2018, 9, 751. [Google Scholar] [CrossRef] [Green Version]
- Shi, C.; Dai, Y.A.; Xia, B.; Xu, X.; Xie, Y.; Liu, Q. The purification and spectral properties of polyphenol oxidase i from Nicotiana tabacum. Plant Mol. Biol. Rep. 2001, 19, 381–382. [Google Scholar] [CrossRef]
- Sen, S.; Mukherji, S. Season-controlled changes in biochemical constituents and oxidase enzyme activities in tomato (Lycopersicon esculentum Mill). J. Environ. Biol. 2009, 30, 479–483. [Google Scholar]
- Mittler, R. ROS are good. Trends Plant Sci. 2017, 22, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Piotrowska, A.; Bajguz, A.; Godlewska-Zyłkiewicz, B.; Czerpak, R.; Kamińska, M. Jasmonic acid as modulator of lead toxicity in aquatic plant Wolffia Arrhiza (Lemnaceae). Environ. Exp. Bot. 2009, 66, 507–513. [Google Scholar] [CrossRef]
- Kadioglu, A.; Saruhan, N.; Sağlam, A.; Terzi, R.; Acet, T. Exogenous salicylic acid alleviates effects of long term drought stress and delays leaf rolling by inducing antioxidant system. Plant Growth Regul. 2011, 64, 27–37. [Google Scholar] [CrossRef]
- Qiu, Z.; Guo, J.; Zhu, A.; Zhang, L.; Zhang, M. Exogenous jasmonic acid can enhance tolerance of wheat seedlings to salt stress. Ecotoxicol. Environ. Saf. 2014, 104, 202–208. [Google Scholar] [CrossRef]
- Agami, R.A.; Mohamed, G.F. Exogenous treatment with indole-3-acetic acid and salicylic acid alleviates cadmium toxicity in wheat seedlings. Ecotoxicol. Environ. Saf. 2013, 94, 164–171. [Google Scholar] [CrossRef]
- Kawano, T. Roles of the reactive oxygen species-generating peroxidase reactions in plant defense and growth induction. Plant Cell Rep. 2003, 21, 829–837. [Google Scholar] [CrossRef]
- Cui, J.; Zhang, R.; Wu, G.L.; Zhu, H.M.; Yang, H. Salicylic acid reduces napropamide toxicity by preventing its accumulation in rapeseed (Brassica Napus L.). Arch. Environ. Contam. Toxicol. 2009, 59, 100–108. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khatami, S.A.; Barmaki, M.; Alebrahim, M.T.; Bajwa, A.A. Salicylic Acid Pre-Treatment Reduces the Physiological Damage Caused by the Herbicide Mesosulfuron-methyl + Iodosulfuron-methyl in Wheat (Triticum aestivum). Agronomy 2022, 12, 3053. https://doi.org/10.3390/agronomy12123053
Khatami SA, Barmaki M, Alebrahim MT, Bajwa AA. Salicylic Acid Pre-Treatment Reduces the Physiological Damage Caused by the Herbicide Mesosulfuron-methyl + Iodosulfuron-methyl in Wheat (Triticum aestivum). Agronomy. 2022; 12(12):3053. https://doi.org/10.3390/agronomy12123053
Chicago/Turabian StyleKhatami, Seyedeh Asieh, Morteza Barmaki, Mohammad Taghi Alebrahim, and Ali Ahsan Bajwa. 2022. "Salicylic Acid Pre-Treatment Reduces the Physiological Damage Caused by the Herbicide Mesosulfuron-methyl + Iodosulfuron-methyl in Wheat (Triticum aestivum)" Agronomy 12, no. 12: 3053. https://doi.org/10.3390/agronomy12123053