

Physiological Responses of Chickpea Genotypes to Cold and Heat Stress in Flowering Stage

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
- (1.)
- Add 400 µL of the 80% ETOH solution (for 200 mL: 160 mL 96% ethanol + 40 mL HEPES stock solution) and vortex.
- (2.)
- Shake for 15 min at 80 °C in an Eppendorf Thermo Mixer at 1300 rpm and centrifuge 1–3 min at 12,000 rpm.
- (3.)
- Collect the supernatant in a 2 mL Eppendorf vial, and keep it on ice in the dark.
- (4.)
- Extract the remaining residue again with 400 µL of the 50% ETOH solution (for 50 mL: 25 mL 96% ethanol, 10 mL HEPES stock solution, 15 mL dd H2O).
- (5.)
- Repeat step 2 and collect the supernatant and pool it with the first one.
- (6.)
- Repeat ethanol extraction (heating included) twice with 200 µL of 80% ETOH until the pellet is clear.
- (1.)
- Add to the pellets in the Eppendorf vials 1000 µL dd H2O plus one metal ball and put in the ball mill (Resch, 200 mm) for 2 min at 22 Hz.
- (2.)
- Autoclave the Eppendorf vials for 90 min, cool and vortex them afterwards.
- (3.)
- Place 100 µL sample plus 400 µL buffer enzyme solution (needs to be freshly prepared, 400 µL Na-Acetate, 0.184 mg Amyloglucosidase, 0.16 µL α-amylase) into 2 mL Eppendorf tubes.
- (4.)
- Incubate for 16 h at 37 °C and 750 rpm in the Thermo Mixer.
- (5.)
- Centrifuge the samples for 5 min at 13,000 rpm until the solution is clear
- (6.)
- Diluted and filter samples before analysis of glucose equivalents by ion chromatography.
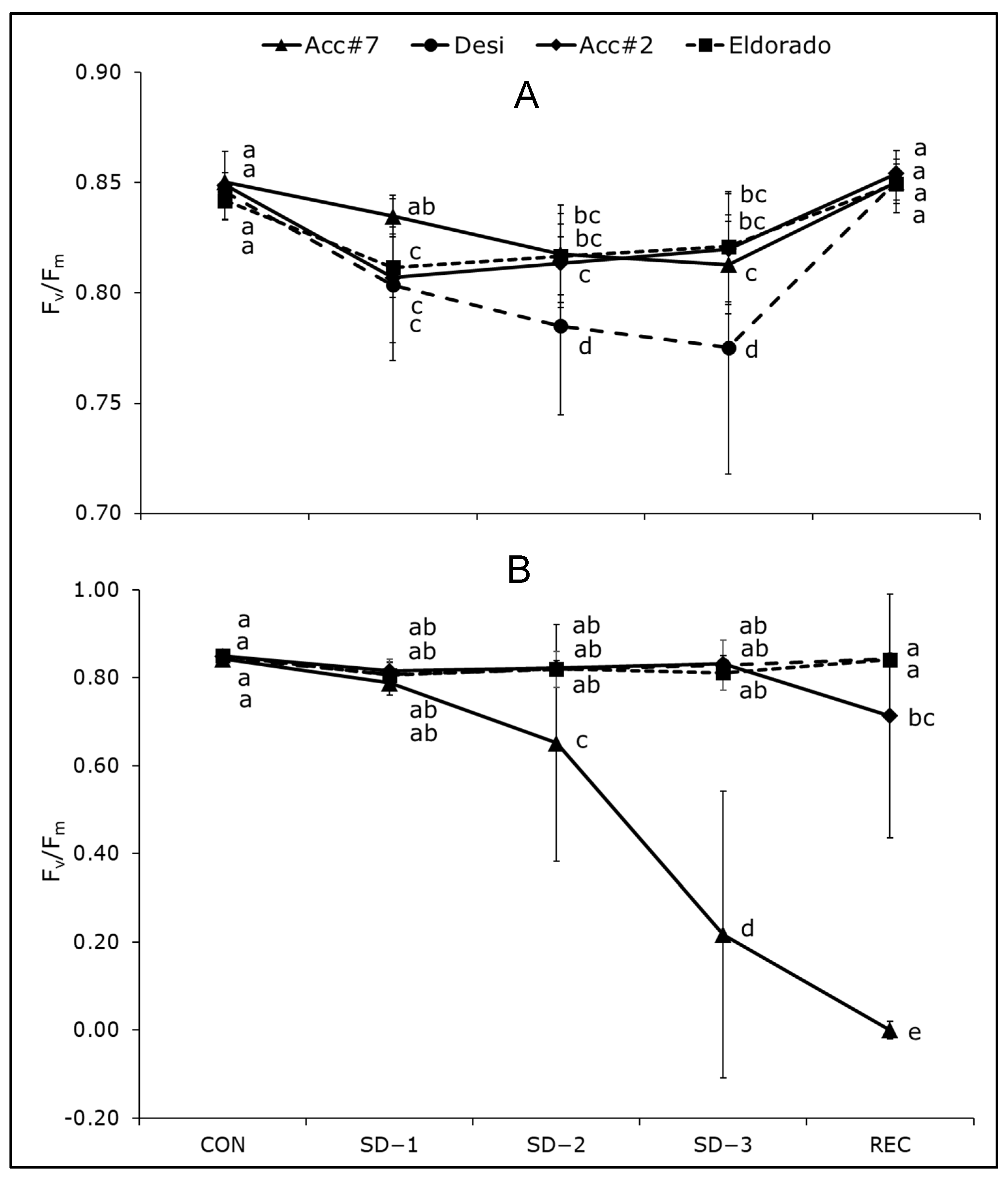
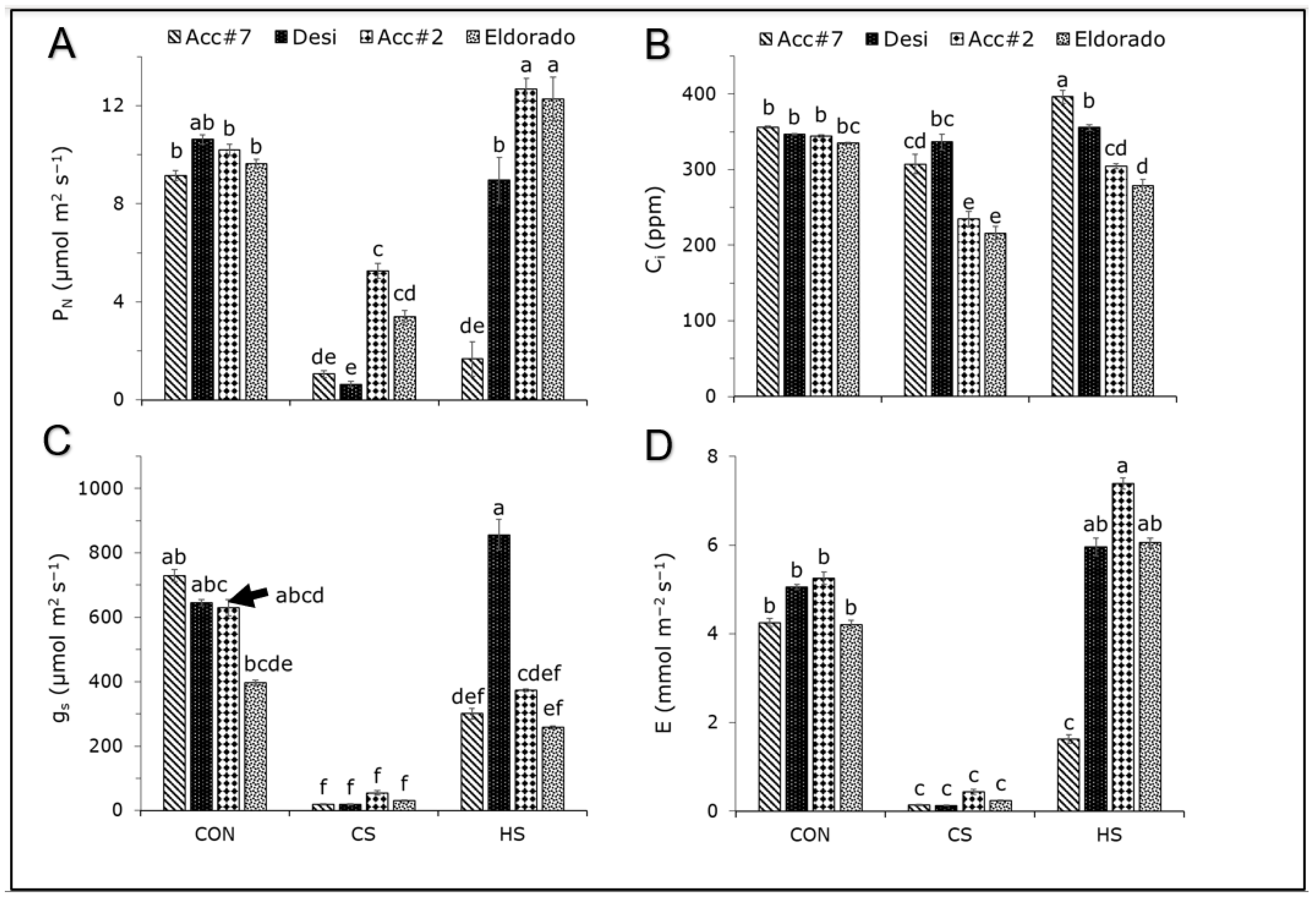
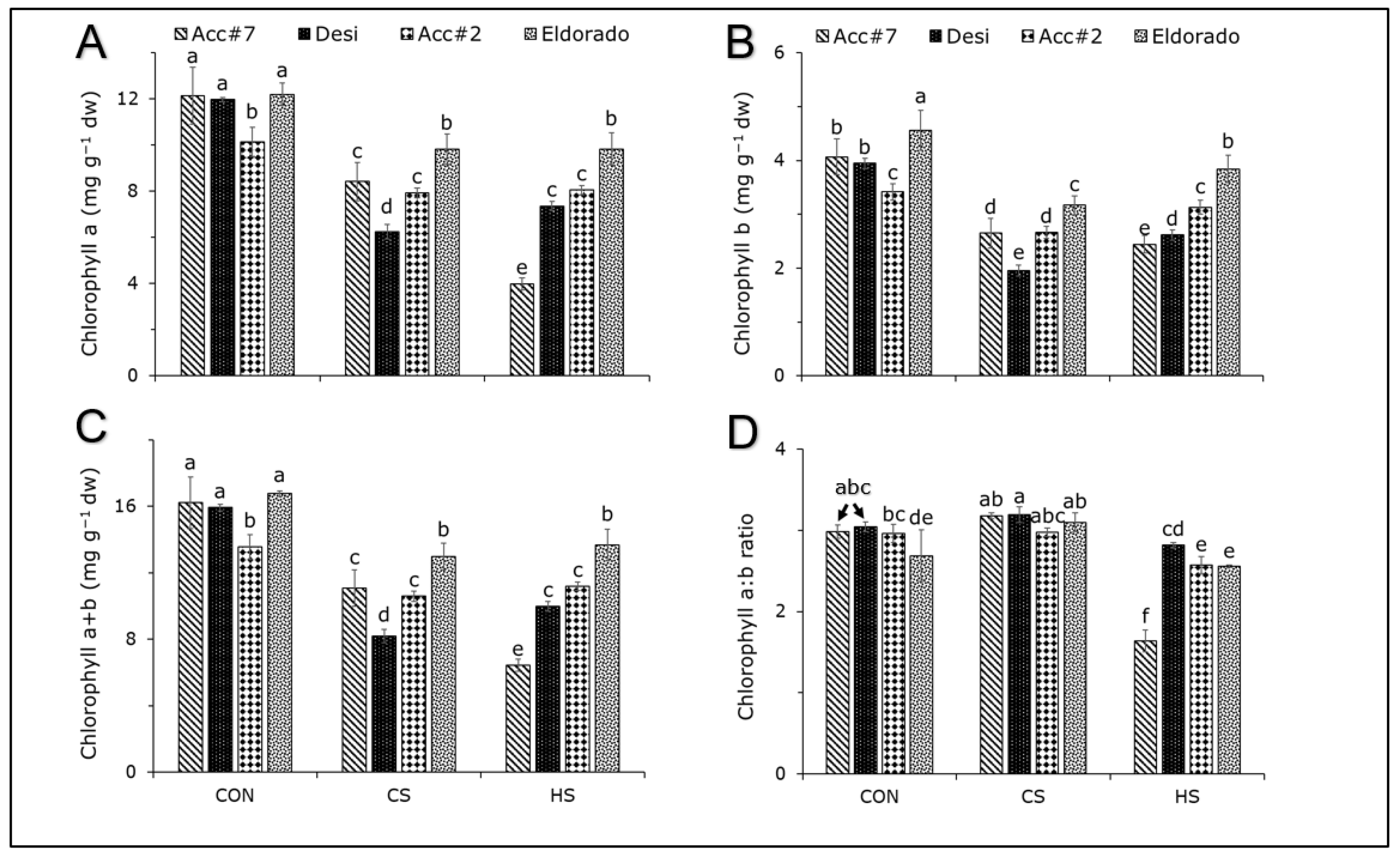
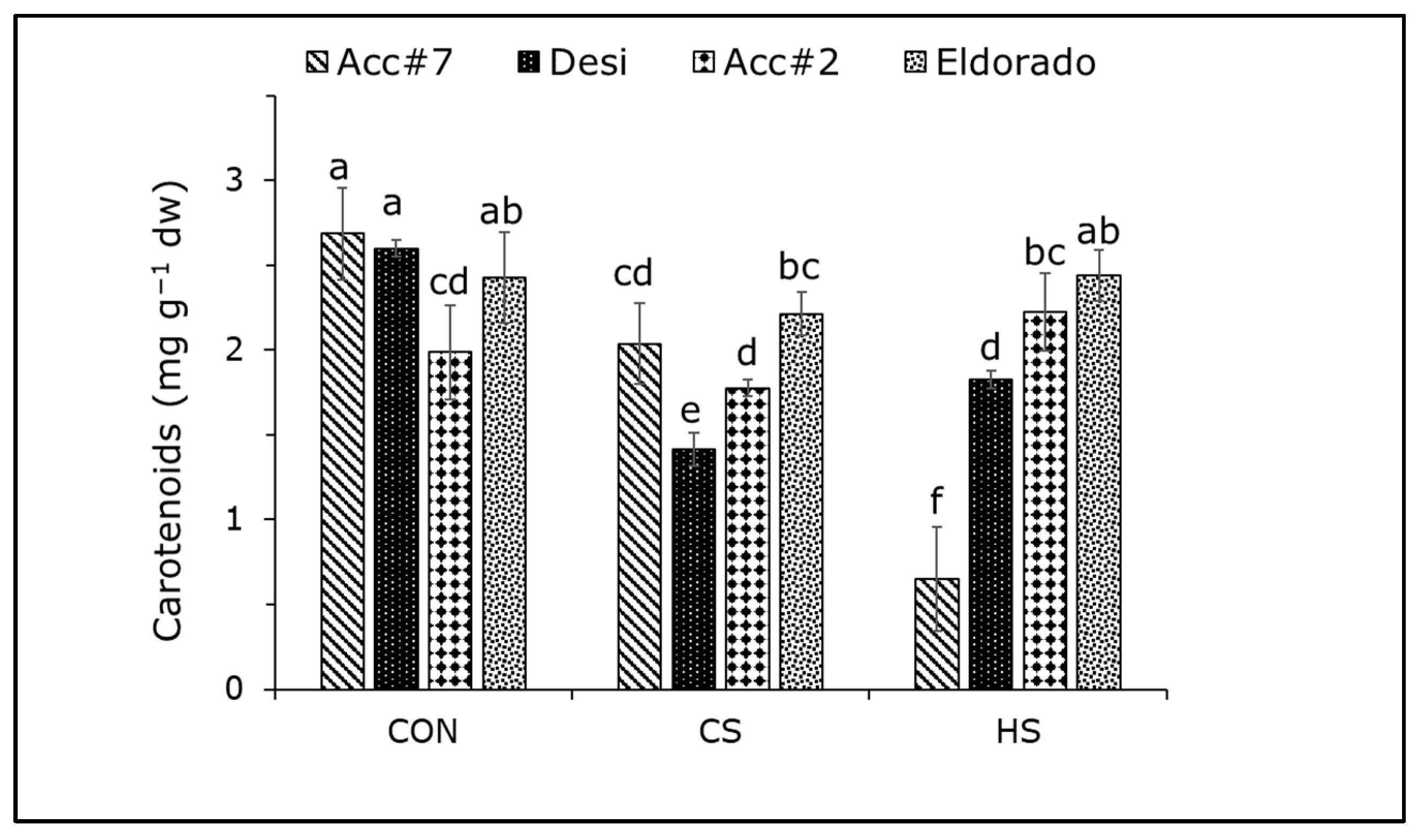
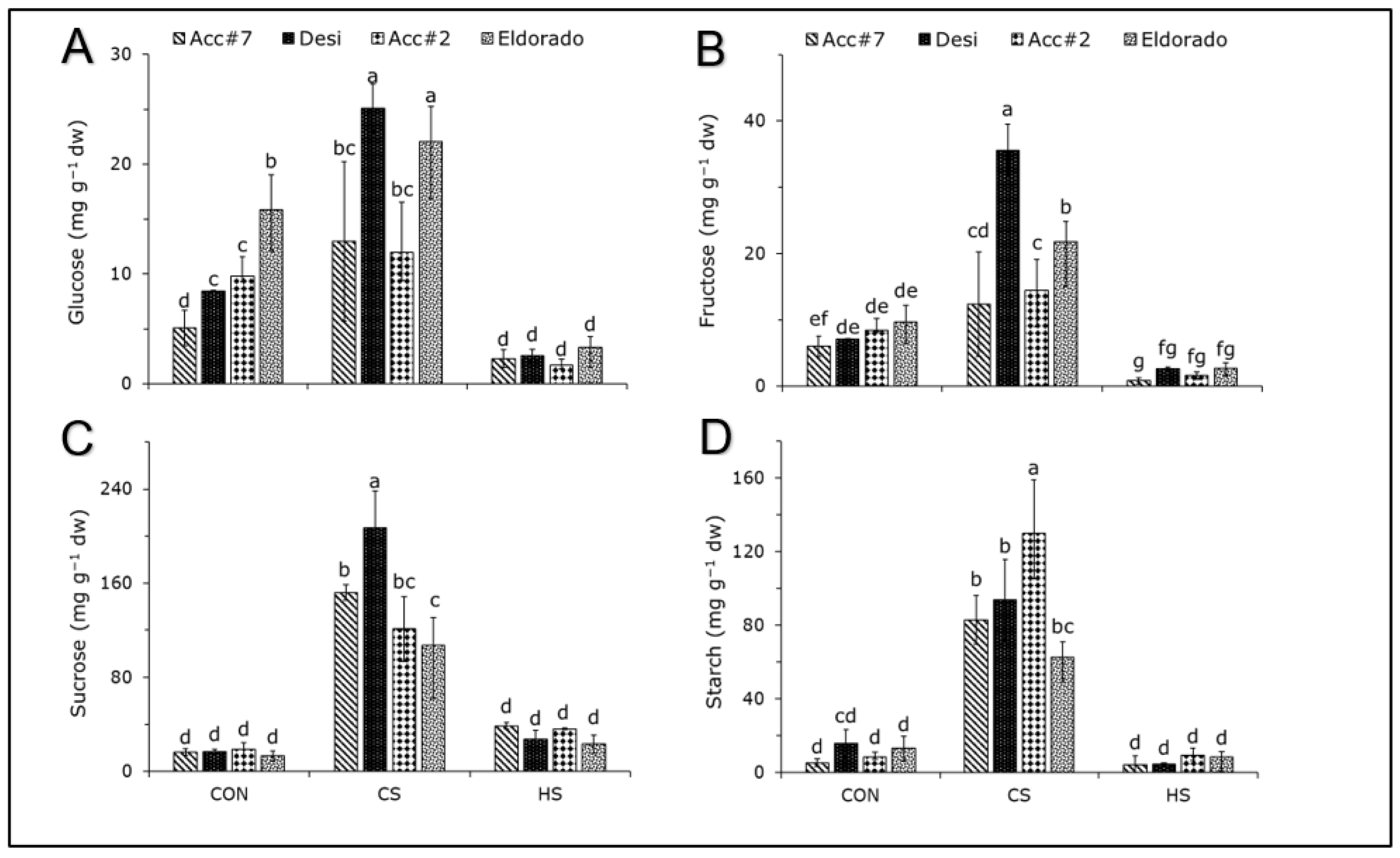
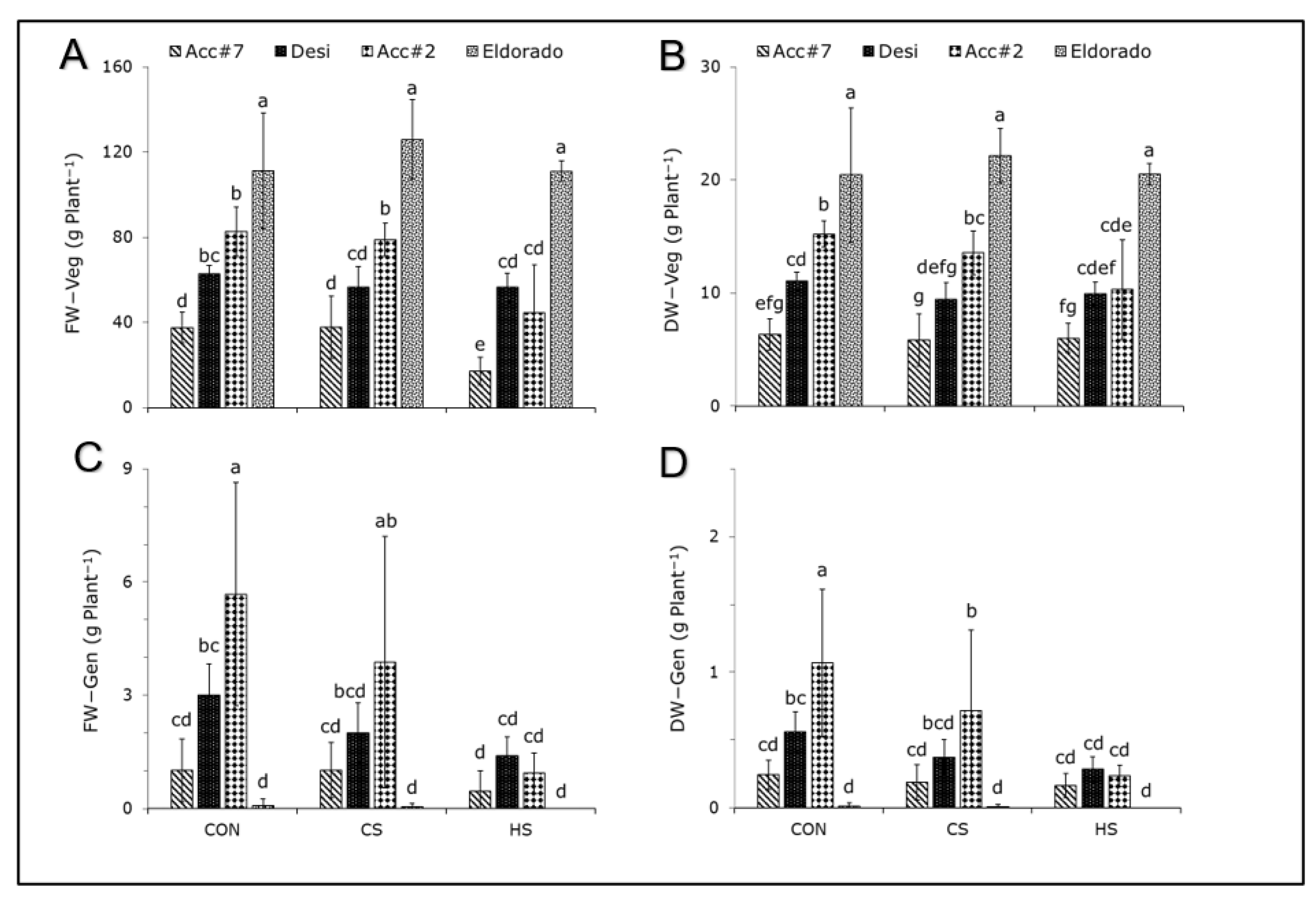
3. Results
4. Discussion
4.1. Effect of Cold Stress on Photosynthetic Apparatus and Carbohydrate Accumulation in Chickpeas
4.2. Photosynthetic Heat Response and Its Strong Dependency on the Chickpea Genotype
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Alcamo, J.; Olesen, J. Life in Europe under Climate Change; John Wiley & Sons: Hoboken, NJ, USA, 2012. [Google Scholar]
- Hopkin, W.; Hüner, N. Introduction to Plant Physiology, 4th ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2009. [Google Scholar]
- Ruelland, E.; Vaultier, M.-N.; Zachowski, A.; Hurry, V. Chapter 2 Cold Signalling and Cold Acclimation in Plants. In Advances in Botanical Research; Elsevier Science & Technology: Amsterdam, The Netherlands, 2009; Volume 49, pp. 35–150. [Google Scholar]
- Hemantaranjan, A.; Malik, C.P.; Bhanu, A.N. Physiology of Heat Stress and Tolerance Mechanisms—An Overview. J. Plant Sci. Res. 2018, 34, 51–64. [Google Scholar] [CrossRef]
- Beck, E.H.; Fettig, S.; Knake, C.; Hartig, K.; Bhattarai, T. Specific and unspecific responses of plants to cold and drought stress. J. Biosci. 2007, 32, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Kaushal, N.; Bhandari, K.; Siddique, K.H.M.; Nayyar, H. Food crops face rising temperatures: An overview of responses, adaptive mechanisms, and approaches to improve heat tolerance. Cogent Food Agric. 2016, 2, 1134380. [Google Scholar] [CrossRef]
- Shanmugam, S.; Kjaer, K.H.; Ottosen, C.O.; Rosenqvist, E.; Kumari Sharma, D.; Wollenweber, B. The Alleviating Effect of Elevated CO2 on Heat Stress Susceptibility of Two Wheat (Triticum aestivum L.) Cultivars. J. Agron. Crop Sci. 2013, 199, 340–350. [Google Scholar] [CrossRef]
- Yadav, S.K. Cold stress tolerance mechanisms in plants. A review. Agron. Sustain. Dev. 2010, 30, 515–527. [Google Scholar] [CrossRef] [Green Version]
- Kumari, P.; Rastogi, A.; Yadav, S. Effects of Heat stress and molecular mitigation approaches in orphan legume, Chickpea. Mol. Biol. Rep. 2020, 47, 4659–4670. [Google Scholar] [CrossRef]
- Merga, B.; Haji, J. Economic importance of chickpea: Production, value, and world trade. Cogent Food Agric. 2019, 5, 1615718. [Google Scholar] [CrossRef]
- Devasirvatham, V.; Tan, D.K.Y. Impact of High Temperature and Drought Stresses on Chickpea Production. Agronomy 2018, 8, 145. [Google Scholar] [CrossRef] [Green Version]
- Stoddard, F.L.; Balko, C.; Erskine, W.; Khan, H.R.; Link, W.; Sarker, A. Screening techniques and sources of resistance to abiotic stresses in cool-season food legumes. Euphytica 2006, 147, 167–186. [Google Scholar] [CrossRef]
- Siddique, K.H.M.; Siddique, K.H.M.; Johansen, C.; Johansen, C.; Turner, N.C.; Turner, N.C.; Jeuffroy, M.-H.; Jeuffroy, M.-H.; Hashem, A.; Hashem, A.; et al. Innovations in agronomy for food legumes. A review. Agron. Sustain. Dev. 2012, 32, 45–64. [Google Scholar] [CrossRef]
- Sita, K.; Sehgal, A.; HanumanthaRao, B.; Nair, R.M.; Vara Prasad, P.V.; Kumar, S.; Gaur, P.M.; Farooq, M.; Siddique, K.H.M.; Varshney, R.K.; et al. Food Legumes and Rising Temperatures: Effects, Adaptive Functional Mechanisms Specific to Reproductive Growth Stage and Strategies to Improve Heat Tolerance. Front. Plant Sci. 2017, 8, 1658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Gan, Y.T.; Clarke, F.; McDonald, C.L. Response of Chickpea Yield to High Temperature Stress during Reproductive Development. Crop Sci. 2006, 46, 2171–2178. [Google Scholar] [CrossRef]
- Kadiyala, M.D.M.; Charyulu, D.K.; Nedumaran, S.; Shyam, D.M.; Gumma, M.K.; Bantilan, M.C.S. Agronomic management options for sustaining chickpea yield under climate change scenario. J. Agrometeorol. 2016, 18, 41. [Google Scholar] [CrossRef]
- Rani, A.; Devi, P.; Jha, U.C.; Sharma, K.D.; Siddique, K.H.M.; Nayyar, H. Developing Climate-Resilient Chickpea Involving Physiological and Molecular Approaches With a Focus on Temperature and Drought Stresses. Front. Plant Sci. 2020, 10, 1759. [Google Scholar] [CrossRef]
- Krishnamurthy, L.; Gaur, P.M.; Basu, P.S.; Chaturvedi, S.K.; Tripathi, S.; Vadez, V.; Rathore, A.; Varshney, R.K.; Gowda, C.L.L. Large genetic variation for heat tolerance in the reference collection of chickpea (Cicer arietinum L.) germplasm. Plant Genet. Resour. 2011, 9, 59–69. [Google Scholar] [CrossRef] [Green Version]
- Srinivasan, A.; Saxena, N.P.; Johansen, C. Cold tolerance during early reproductive growth of chickpea (Cicer arietinum L.): Genetic variation in gamete development and function. Field Crops Res. 1999, 60, 209–222. [Google Scholar] [CrossRef] [Green Version]
- Makonya, G.M.; Ogola, J.B.O.; Muthama Muasya, A.; Crespo, O.; Maseko, S.; Valentine, A.J.; Ottosen, C.-O.; Rosenqvist, E.; Chimphango, S.B.M. Chlorophyll fluorescence and carbohydrate concentration as field selection traits for heat tolerant chickpea genotypes. Plant Physiol. Biochem. 2019, 141, 172–182. [Google Scholar] [CrossRef]
- Zhou, R.; Yu, X.; Huang, S.; Song, X.; Rosenqvist, E.; Ottosen, C.-O. Genotype-dependent responses of chickpea to high temperature and moderately increased light. Plant Physiol. Biochem. 2020, 154, 353–359. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1987; Volume 148, pp. 350–382. [Google Scholar]
- Croser, J.S.; Clarke, H.J.; Siddique, K.H.M.; Khan, T.N. Low-Temperature Stress: Implications for Chickpea (Cicer arietinum L.) Improvement. Crit. Rev. Plant Sci. 2003, 22, 185–219. [Google Scholar] [CrossRef]
- Baker, N.R.; Rosenqvist, E. Applications of chlorophyll fluorescence can improve crop production strategies: An examination of future possibilities. J. Exp. Bot. 2004, 55, 1607–1621. [Google Scholar] [CrossRef]
- Zhou, R.; Kong, L.; Wu, Z.; Rosenqvist, E.; Wang, Y.; Zhao, L.; Zhao, T.; Ottosen, C.-O. Physiological response of tomatoes at drought, heat and their combination followed by recovery. Physiol. Plantarum 2018, 165, 144–154. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.; Hussain, M.; Nawaz, A.; Lee, D.-J.; Alghamdi, S.S.; Siddique, K.H.M. Seed priming improves chilling tolerance in chickpea by modulating germination metabolism, trehalose accumulation and carbon assimilation. Plant Physiol. Biochem. 2017, 111, 274–283. [Google Scholar] [CrossRef] [PubMed]
- Pu-Hai, L.; Gan, Y.; Warkentin, T.; McDonald, C. Morphological plasticity of chickpea in a semiarid environment. Crop Sci. 2003, 43, 426–429. [Google Scholar]
- Nayyar, H.; Bains, T.S.; Kumar, S. Chilling stressed chickpea seedlings: Effect of cold acclimation, calcium and abscisic acid on cryoprotective solutes and oxidative damage. Environ. Exp. Bot. 2005, 54, 275–285. [Google Scholar] [CrossRef]
- Kaushal, N.; Awasthi, R.; Gupta, K.; Gaur, P.; Siddique, K.H.M.; Nayyar, H. Heat-stress-induced reproductive failures in chickpea (Cicer arietinum) are associated with impaired sucrose metabolism in leaves and anthers. Funct. Plant Biol. 2013, 40, 1334–1349. [Google Scholar] [CrossRef]
- Lyu, J.I.; Ramekar, R.; Kim, J.M.; Hung, N.N.; Seo, J.S.; Kim, J.-B.; Choi, I.-Y.; Park, K.-C.; Kwon, S.-J. Unraveling the complexity of faba bean (Vicia faba L.) transcriptome to reveal cold-stress-responsive genes using long-read isoform sequencing technology. Sci. Rep. 2021, 11, 21094. [Google Scholar] [CrossRef] [PubMed]
- Devasirvatham, V.; Tan, D.K.Y.; Gaur, P.M.; Raju, T.N.; Trethowan, R.M. High temperature tolerance in chickpea and its implications for plant improvement. Crop Pasture Sci. 2012, 63, 419–422. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Thakur, P.; Kaushal, N.; Malik, J.A.; Gaur, P.; Nayyar, H. Effect of varying high temperatures during reproductive growth on reproductive function, oxidative stress and seed yield in chickpea genotypes differing in heat sensitivity. Arch. Agron. Soil Sci. 2013, 59, 823–843. [Google Scholar] [CrossRef]
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M.R. Heat tolerance in plants: An overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Eker, T.; Sari, H.; Sari, D.; Canci, H.; Arslan, M.; Aydinoglu, B.; Ozay, H.; Toker, C. Advantage of Multiple Pods and Compound Leaf in Kabuli Chickpea under Heat Stress Conditions. Agronomy 2022, 12, 557. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeitelhofer, M.; Zhou, R.; Ottosen, C.-O. Physiological Responses of Chickpea Genotypes to Cold and Heat Stress in Flowering Stage. Agronomy 2022, 12, 2755. https://doi.org/10.3390/agronomy12112755
Zeitelhofer M, Zhou R, Ottosen C-O. Physiological Responses of Chickpea Genotypes to Cold and Heat Stress in Flowering Stage. Agronomy. 2022; 12(11):2755. https://doi.org/10.3390/agronomy12112755
Chicago/Turabian StyleZeitelhofer, Mareen, Rong Zhou, and Carl-Otto Ottosen. 2022. "Physiological Responses of Chickpea Genotypes to Cold and Heat Stress in Flowering Stage" Agronomy 12, no. 11: 2755. https://doi.org/10.3390/agronomy12112755