
Earthworm Lumbricus terrestris Contributes Nitrous Oxide Emission from Temperate Agricultural Soil Regardless of Applied Mineral Nitrogen Fertilizer Doses

Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil Samples and Earthworms Collection
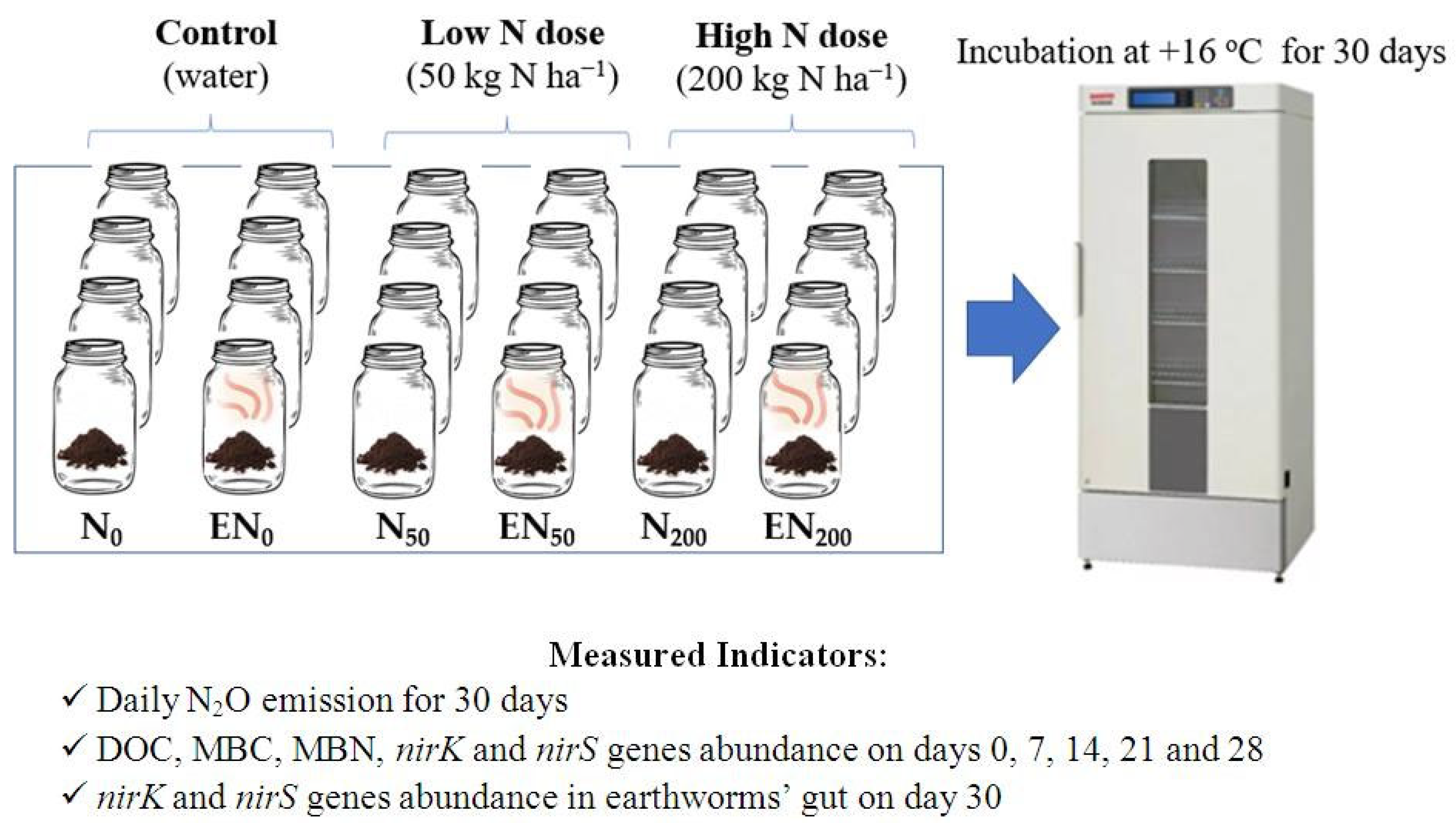
2.2. The Experiment Design
2.3. Soil Analyses
2.4. Earthworms Gut Isolation
2.5. DNA Extraction and Quantitative PCR Analysis of Genes Abundance
2.6. Statistical Analyses
3. Results
3.1. Earthworms Survival and Growth
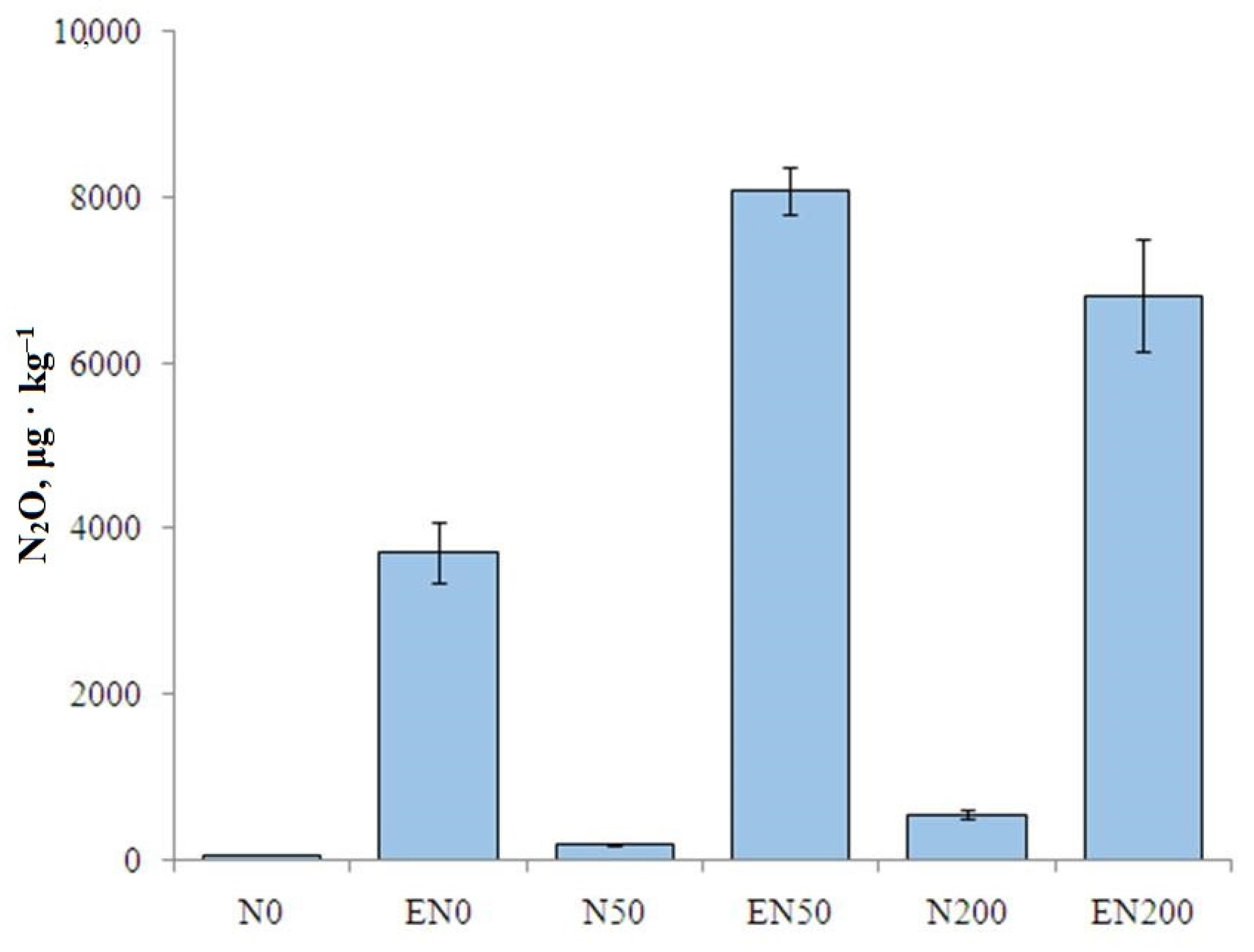
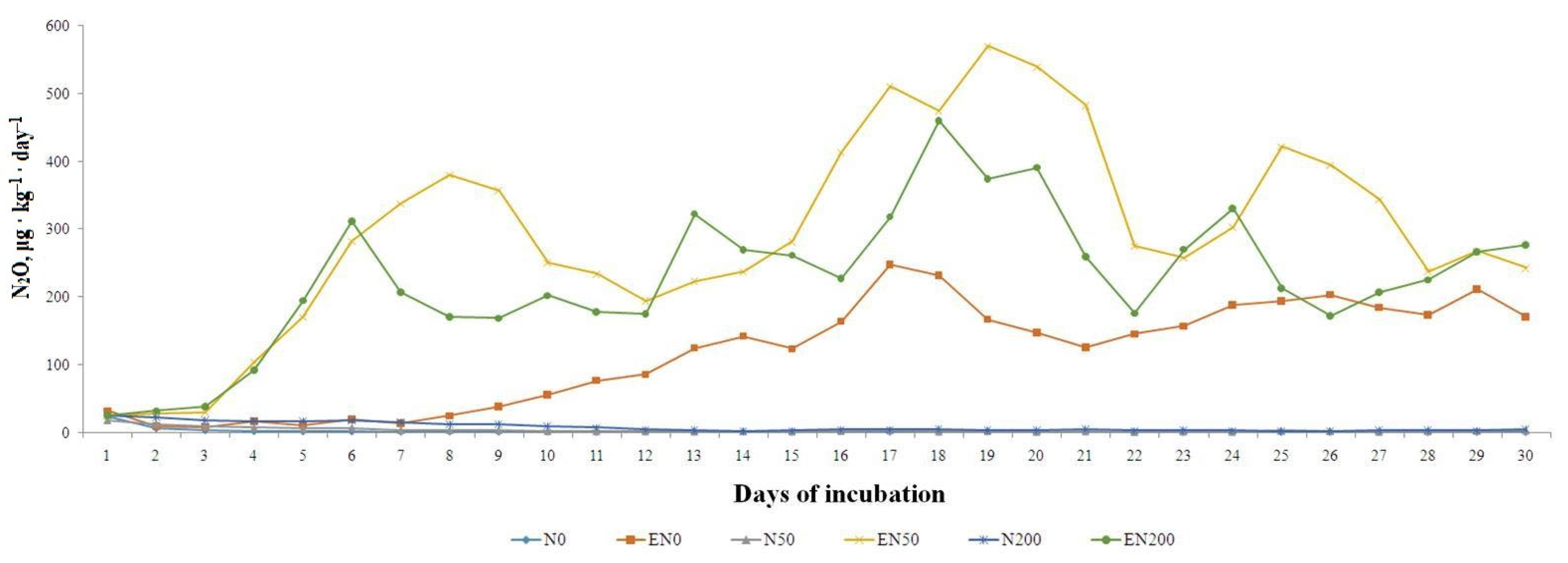
3.2. Soil Nitrous Oxide Emission
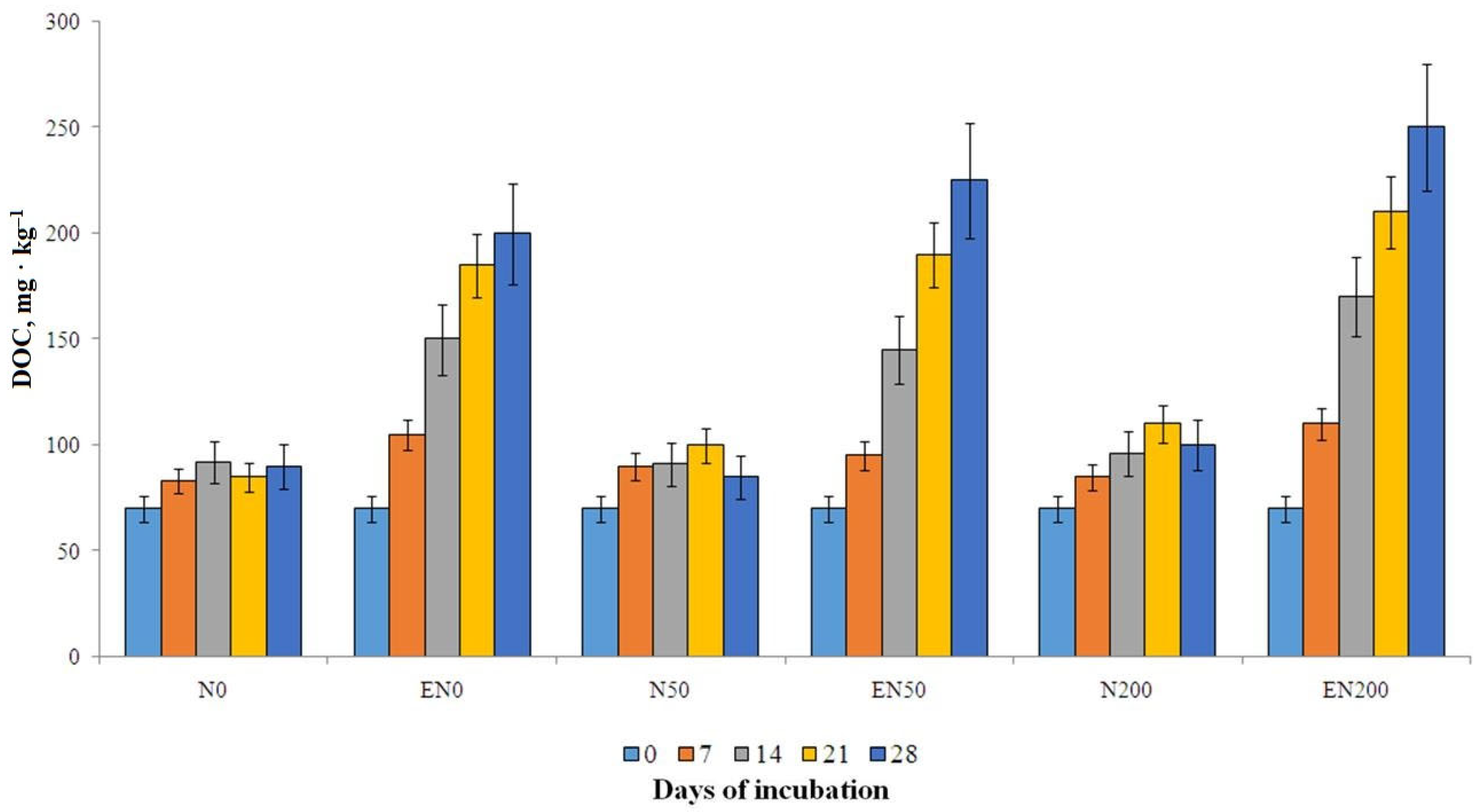
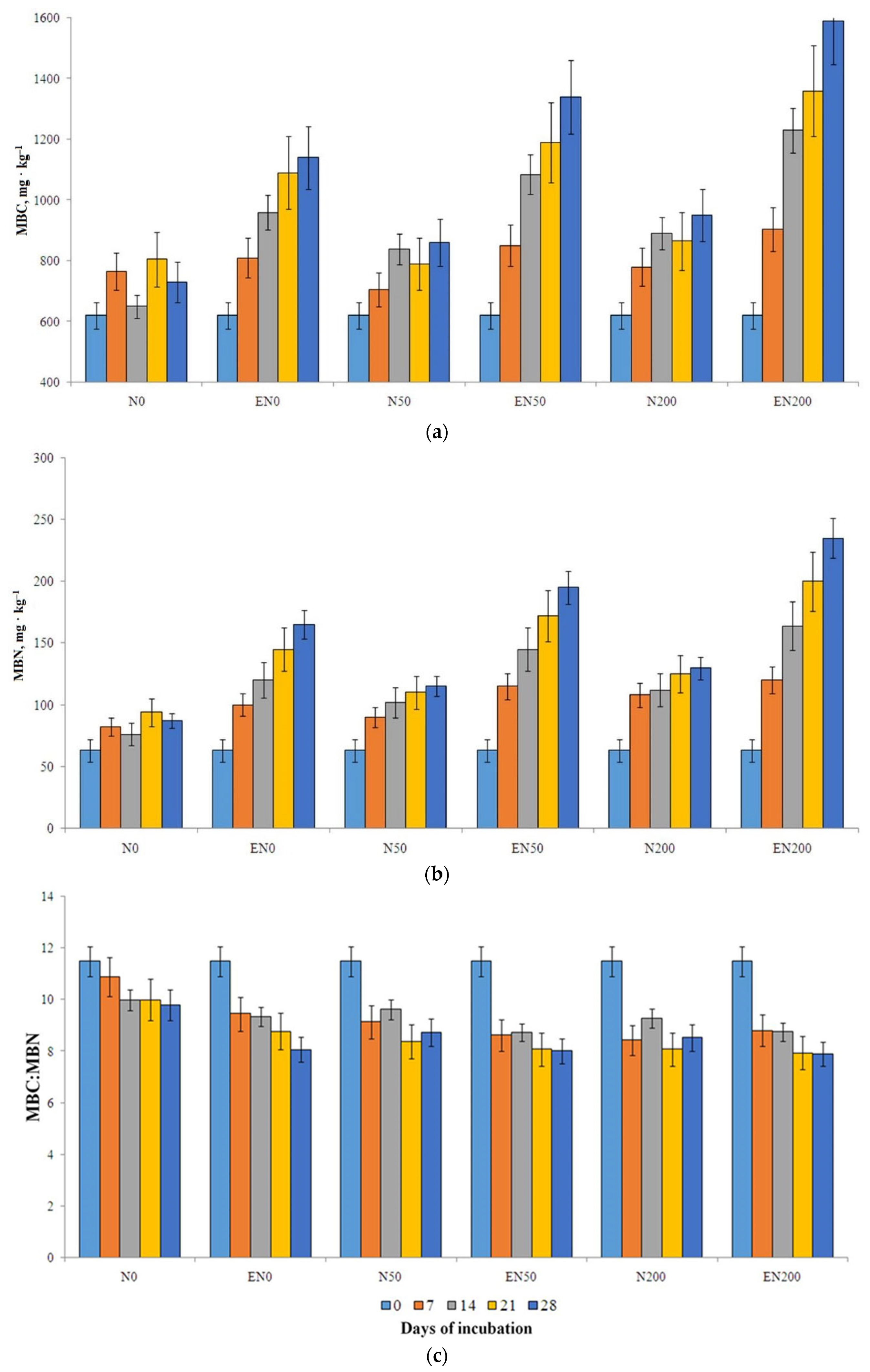
3.3. The Content of DOC, MBC and MBN in Soil
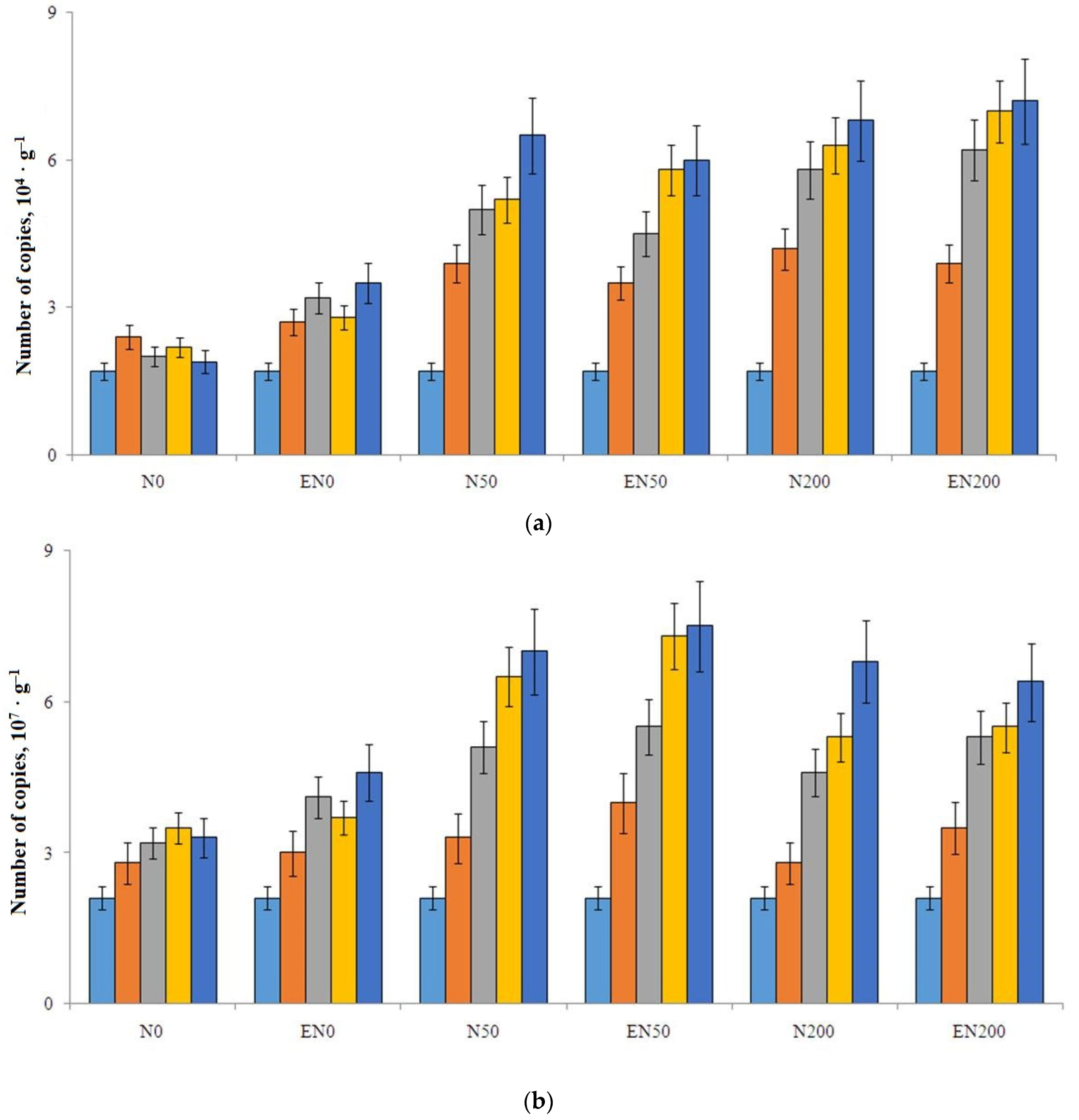
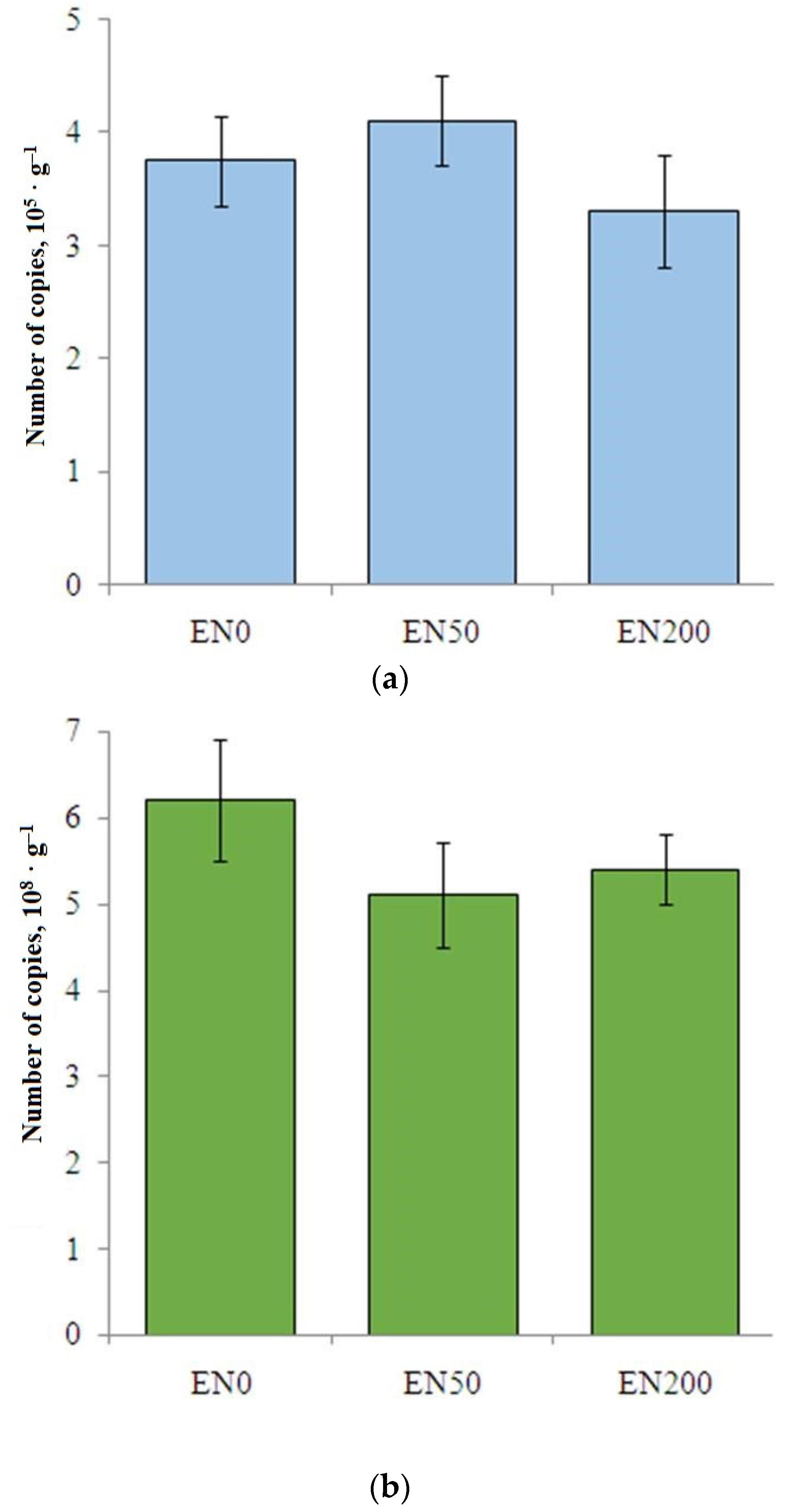
3.4. Abundance of nirK and nirS Genes in Soil and Earthworm Gut
4. Discussion
4.1. Influence of Earthworms on Nitrous Oxide Emission from Soil
4.2. How Earthworms Affect the Soil N2O Emission
4.3. Influence of the Mineral N Fertilizer Dose on the Soil N2O Emission
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Solomon, S.; Qin, M.; Manning, M. Climate change 2007: The physical science basis. In Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Ravishankara, A.R.; Daniel, J.S.; Portmann, R.W. Nitrous oxide (N2O): The dominant ozone-depleting substance emitted in the 21st century. Science 2009, 326, 123–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reay, D.S.; Davidson, E.A.; Smith, K.A.; Smith, P.; Melillo, J.M.; Dentener, F.; Crutzen, P.J. Global agriculture and nitrous oxide emissions. Nat. Clim. Chang. 2012, 2, 410–416. [Google Scholar] [CrossRef]
- Kudeyarov, V.N. Nitrous oxide emission from fertilized soils: An analytical review. Eurasian Soil Sci. 2020, 53, 1396–1407. [Google Scholar] [CrossRef]
- Thilakarathna, S.K.; Hernandez-Ramirez, G.; Puurveen, D.; Kryzanowski, L.; Lohstraeter, G.; Powers, L.; Quan, N.; Tenuta, M. Nitrous oxide emissions and nitrogen use efficiency in wheat: Nitrogen fertilization timing and formulation, soil nitrogen, and weather effects. Soil Sci. Soc. Am. J. 2020, 84, 1910–1927. [Google Scholar] [CrossRef]
- Minikaev, D.; Zurgel, U.; Tripler, E.; Gelfand, I. Effect of increasing nitrogen fertilization on soil nitrous oxide emissions and nitrate leaching in a young date palm (Phoenix dactylifera L., cv. Medjool) orchard. Agric. Ecosyst. Environ. 2021, 319, 107569. [Google Scholar] [CrossRef]
- Wrage, N.; Velthof, G.; van Beusichem, M.; Oenema, O. Role of nitrifier denitrification in the production of nitrous oxide. Soil Biol. Biochem. 2001, 33, 1723–1732. [Google Scholar] [CrossRef]
- Kool, D.M.; Dolfing, J.; Wrage, N.; van Groenigen, J.W. Nitrifier denitrification as a distinct and significant source of nitrous oxide from soil. Soil Biol. Biochem. 2011, 43, 174–178. [Google Scholar] [CrossRef]
- Dobrovol’skaya, T.G.; Zvyagintsev, D.G.; Chernov, I.Y.; Golovchenko, A.V.; Zenova, G.M.; Lysak, L.V.; Manucharova, N.A.; Marfenina, O.E.; Polyanskaya, L.M.; Stepanov, A.L.; et al. The role of microorganisms in the ecological functions of soils. Eurasian Soil Sci. 2015, 48, 959–967. [Google Scholar] [CrossRef]
- Hallin, S.; Philippot, L.; Loffler, F.E. Genomics and ecology of novel N2O reducing microorganisms. Trends Microbiol. 2018, 26, 43–55. [Google Scholar] [CrossRef]
- Stehfest, E.; Bouwman, L. N2O and NO emission from agricultural fields and soils under natural vegetation: Summarizing available measurement data and modeling of global annual emissions. Nutr. Cycl. Syst. 2006, 74, 207–228. [Google Scholar] [CrossRef]
- Lin, S.; Hernandez-Ramirez, G.; Kryzanowski, L.; Wallace, T.; Grant, R.; Degenhardt, R. Timing of manure injection and nitrification inhibitors impacts on nitrous oxide emissions and nitrogen transformations in a barley crop. Soil Sci. Soc. Am. J. 2017, 81, 1595–1605. [Google Scholar] [CrossRef] [Green Version]
- Maslov, M.N.; Pozdnyakov, L.A. Application of crop residues in combination with a mineral nitrogen fertilizer to Albic Retisols and their effect on the nitrous oxide production. Mosc. Univ. Soil Sci. Bull. 2022, 1, 46–52. [Google Scholar] [CrossRef]
- Blouin, M.; Hodson, M.E.; Delgado, E.A.; Baker, G.; Brussaard, L.; Butt, K.R.; Dai, J.; Dendooven, L.; Peres, G.; Tondoh, J.E.; et al. A review of earthworm impact on soil function and ecosystem services. Eur. J. Soil Sci. 2013, 64, 161–182. [Google Scholar] [CrossRef]
- Kim, Y.N.; Robinson, B.; Boyer, S.; Zhong, H.T.; Dickinson, N. Interactions of native and introduced earthworms with soils and plant rhizospheres in production landscapes of New Zealand. Appl. Soil Ecol. 2015, 96, 141–150. [Google Scholar] [CrossRef]
- Chapuis-Lardy, L.; Brauman, A.; Bernard, L.; Pablo, A.L.; Toucet, J.; Mano, M.J.; Weber, L.; Brunet, D.; Razafimbelo, T.; Chotte, J.L.; et al. Effect of the endogeic earthworm Pontoscolex corethrurus on the microbial structure and activity related to CO2 and N2O fluxes from a tropical soil (Madagascar). Appl. Soil Ecol. 2010, 45, 201–208. [Google Scholar] [CrossRef]
- Moore, J.C.; de Ruiter, P.C. Energetic food webs. In An Analysis of Real and Model Ecosystems; Oxford University Press: Oxford, UK, 2012. [Google Scholar]
- Huang, K.; Xia, H.; Cui, G.; Li, F. Effects of earthworms on nitrification and ammonia oxidizers in vermicomposting systems for recycling of fruit and vegetable wastes. Sci. Total Envir. 2017, 578, 337–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medina-Sauza, R.M.; Álvarez-Jiménez, M.; Delhal, A.; Reverchon, F.; Blouin, M.; Guerrero-Analco, J.A.; Cerdán, C.R.; Guevara, R.; Villain, L.; Barois, I. Earthworms building up soil microbiota, a Review. Front. Envir. Sci. 2019, 7, 81. [Google Scholar] [CrossRef] [Green Version]
- Lubbers, I.M.; van Groenigen, K.J.; Fonte, S.J.; Six, J.; Brussaard, L.; van Groenigen, J.W. Greenhouse-gas emissions from soils increased by earthworms. Nat. Clim. Chang. 2013, 3, 187–194. [Google Scholar] [CrossRef]
- Amador, J.; Avizinis, E. Response of nitrous oxide flux to addition of anecic earthworms to an agricultural field. Open J. Soil Sci. 2013, 3, 100–106. [Google Scholar] [CrossRef] [Green Version]
- Nieminen, M.; Hurme, T.; Mikola, J.; Regina, K.; Nuutinen, V. Impact of earthworm Lumbricus terrestris living sites on the greenhouse gas balance of no-till arable soil. Biogeosciences 2015, 12, 5481–5493. [Google Scholar] [CrossRef]
- Wu, D.; Liu, M.; Song, X.; Jiao, J.; Li, H.; Hu, F. Earthworm ecosystem service and dis-service in an N-enriched agroecosystem: Increase of plant production leads to no effects on yield-scaled N2O emissions. Soil Biol. Biochem. 2015, 82, 1–8. [Google Scholar] [CrossRef]
- Abail, Z.; Whalen, J.K. Earthworm contributions to soil nitrogen supply in corn-soybean agroecosystems in Quebec, Canada. Pedosphere 2021, 31, 405–412. [Google Scholar] [CrossRef]
- Parkin, T.B.; Berry, E. Microbial nitrogen transformations in earthworm burrows. Soil Biol. Biochem. 1999, 31, 1765–1771. [Google Scholar] [CrossRef]
- Brown, G.G.; Barois, I.; Lavelle, P. Regulation of soil organic matter dynamics and microbial activity in the drilosphere and the role of interactions with other edaphic functional domains. Eur. J. Soil Biol. 2000, 36, 177–198. [Google Scholar] [CrossRef]
- Drake, H.L.; Horn, M.A. As the worm turns: The earthworm gut as a transient habitat for soil microbial biomes. Annu. Rev. Microbiol. 2007, 61, 169–189. [Google Scholar] [CrossRef]
- Wu, Y.; Shaaban, M.; Zhao, J.; Hao, R.; Hu, R. Effect of the earthworm gut-stimulated denitrifiers on soil nitrous oxide emissions. Eur. J. Soil Biol. 2015, 70, 104–110. [Google Scholar] [CrossRef]
- Ihssen, J.; Horn, M.A.; Matthies, C.; Gossner, A.; Schramm, A.; Drake, H.L. N2O-producing microorganisms in the gut of the earthworm Aporrectodea caliginosa are indicative of ingested soil bacteria. Appl. Environ. Microbiol. 2003, 69, 1655–1661. [Google Scholar] [CrossRef] [Green Version]
- Giannopoulos, G.; Pulleman, M.M.; van Groenigen, J.W. Interactions between residue placement and earthworm ecological strategy affect aggregate turnover and N2O dynamics in agricultural soil. Soil Biol. Biochem. 2010, 42, 618–625. [Google Scholar] [CrossRef]
- Nebert, L.D.; Bloem, J.; Lubbers, I.M.; van Groenigen, J.W. Association of earthworm-denitrifier interactions with increased emission of nitrous oxide from soil mesocosms amended with crop residue. Appl. Environ. Microbiol. 2011, 77, 4097–4104. [Google Scholar] [CrossRef] [Green Version]
- IUSS Working Group WRB. World Reference Base for Soil Resources 2014, update 2015 International Soil Classification System for Naming Soils and Creating Legends for Soil Maps. World Soil Resources Report. 2015, p. 192. Available online: https://www.fao.org/3/i3794en/I3794en.pdf (accessed on 20 September 2022).
- Berry, E.C.; Jordan, D. Temperature and soil moisture content effects on the growth of Lumbricus terrestris (Oligochaeta: Lumbricidae) under laboratory conditions. Soil Biol. Biochem. 2001, 33, 133–136. [Google Scholar] [CrossRef]
- Butt, K.R.; Frederickson, J.; Morris, R.M. The intensive production of Lumbricus terrestris L. for soil amelioration. Soil Biol. Biochem. 1992, 24, 1321–1325. [Google Scholar] [CrossRef]
- Daniel, O.; Kohli, L.; Bieri, M. Weight gain and weight loss of the earthworm Lumbricus terrestris L. at different temperatures and body weights. Soil Biol. Biochem. 1996, 28, 1235–1240. [Google Scholar] [CrossRef]
- Castellano-Hinojosa, A.; Correa-Galeote, D.; González-López, J.; Bedmar, E.J. Effect of nitrogen fertilisers on nitrous oxide emission, nitrifier and denitrifier abundance and bacterial diversity in closed ecological systems. Appl. Soil Ecol. 2019, 145, 103380. [Google Scholar] [CrossRef]
- Brookes, P.C.; Landman, A.; Pruden, G.; Jenkinson, D.S. Chloroform fumigation and the release of soil nitrogen: A rapid direct extraction method to measure microbial biomass nitrogen in soil. Soil Biol. Biochem. 1985, 17, 837–842. [Google Scholar] [CrossRef]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Makarov, M.I.; Malysheva, T.I.; Maslov, M.N.; Kuznetsova, E.Y.; Menyailo, O.V. Determination of carbon and nitrogen in microbial biomass of southern-taiga soils by different methods. Eurasian Soil Sci. 2016, 49, 685–695. [Google Scholar] [CrossRef]
- Makarov, M.I.; Kuznetsova, E.Y.; Malysheva, T.I.; Maslov, M.N.; Menyailo, O.V. Effect of the storage conditions of soil samples on carbon and nitrogen extractability. Eurasian Soil Sci. 2017, 50, 549–558. [Google Scholar] [CrossRef]
- Maslov, M.N.; Maslova, O.A.; Tokareva, O.A. Changes in labile and microbial pools of carbon and nitrogen in forest litter samples under different methods of storage. Eurasian Soil Sci. 2019, 52, 747–755. [Google Scholar] [CrossRef]
- Astaykina, A.; Streletskii, R.; Maslov, M.; Krasnov, G.; Gorbatov, V. Effects of three pesticides on the earthworm Lumbricus terrestris gut microbiota. Front. Microbiol. 2022, 13, 853535. [Google Scholar] [CrossRef]
- Henry, S.; Baudoin, E.; López-Gutiérrez, J.C.; Martin-Laurent, F.; Brauman, A.; Philippot, L. Quantification of denitrifying bacteria in soils by nirK gene targeted real-time PCR. J. Microbiol. Methods 2004, 59, 327–335. [Google Scholar] [CrossRef]
- Throbäck, N.; Enwall, K.; Jarvis, A.; Hallin, S. Reassessing PCR primers targeting nirS, nirK and nosZ genes for community surveys of denitrifying bacteria with DGGE. FEMS Microbiol. Ecol. 2004, 49, 401–417. [Google Scholar] [CrossRef] [PubMed]
- Augustenborg, C.A.; Hepp, S.; Kammann, C.; Hagan, D.; Schmidt, O.; Müller, C. Biochar and earthworm effects on soil nitrous oxide and carbon dioxide emissions. J. Environ. Qual. 2012, 41, 1203. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Hu, Z.; Xu, X.; Jiang, X.; Zheng, B.; Liu, X.; Pan, X.; Kardol, P. Emissions of ammonia and greenhouse gases during combined pre-composting and vermicomposting of duck manure. Waste Manag. 2014, 34, 1546–1552. [Google Scholar] [CrossRef] [PubMed]
- Lv, B.; Zhang, D.; Cui, Y.; Yin, F. Effects of C/N ratio and earthworms on greenhouse gas emissions during vermicomposting of sewage sludge. Bioresour. Technol. 2018, 268, 408–414. [Google Scholar] [CrossRef]
- Lv, B.; Cui, Y.; Wei, H.; Chen, Q.; Zhang, D. Elucidating the role of earthworms in N2O emission and production pathway during vermicomposting of sewage sludge and rice straw. J. Hazard. Mater. 2020, 400, 123215. [Google Scholar] [CrossRef]
- Giannopoulos, G.; van Groenigen, J.W.; Pulleman, M.M. Earthworm-induced N2O emissions in a sandy soil with surface-applied crop residues. Pedobiologia 2011, 54, 103–111. [Google Scholar] [CrossRef]
- Wang, C.; Butterbach-Bahl, K.; He, N.; Wang, Q.; Xing, X.; Han, X. Nitrogen addition and mowing affect microbial nitrogen transformations in a C4 grassland in northern China. Eur. J. Soil Sci. 2015, 66, 485–495. [Google Scholar] [CrossRef]
- Chen, C.; Whalen, J.K.; Guo, X. Earthworms reduce soil nitrous oxide emissions during drying and rewetting cycles. Soil Biol. Biochem. 2014, 68, 117–124. [Google Scholar] [CrossRef]
- Bertora, C.; van Vliet, P.C.J.; Hummelink, E.W.J.; van Groenigen, J.W. Do earthworms increase N2O emissions in ploughed grassland? Soil Biol. Biochem. 2007, 39, 632–640. [Google Scholar] [CrossRef]
- Rizhiya, E.; Bertora, C.; van Vliet, P.C.J.; Kuikman, P.J.; Faber, J.H.; van Groenigen, J.W. Earthworm activity as a determinant for N2O emission from crop residue. Soil Biol. Biochem. 2007, 39, 2058–2069. [Google Scholar] [CrossRef]
- Speratti, A.B.; Whalen, J.K. Carbon dioxide and nitrous oxide fluxes from soil as influenced by anecic and endogeic earthworms. Appl. Soil Ecol. 2008, 38, 27–33. [Google Scholar] [CrossRef]
- Depkat-Jakob, P.S.; Brown, G.G.; Tsai, S.M.; Horn, M.A.; Drake, H.L. Emission of nitrous oxide and dinitrogen by diverse earthworm families from Brazil and resolution of associated denitrifying and nitrate-dissimilating taxa. FEMS Microbiol. Ecol. 2012, 83, 375–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Speratti, A.B.; Whalen, J.K.; Rochette, P. Earthworm influence on carbon dioxide and nitrous oxide fluxes from an unfertilized corn agroecosystem. Biol. Fertil. Soils 2007, 44, 405–409. [Google Scholar] [CrossRef]
- Marhan, S.; Auber, J.; Poll, C. Additive effects of earthworms, nitrogen-rich litter and elevated soil temperature on N2O emission and nitrate leaching from an arable soil. Appl. Soil Ecol. 2015, 86, 55–61. [Google Scholar] [CrossRef]
- Braga, L.P.P.; Yoshiura, C.A.; Borges, C.D.; Horn, M.A.; Brown, G.G.; Drake, H.L.; Tsai, S.M. Disentangling the influence of earthworms in sugarcane rhizosphere. Sci. Rep. 2016, 6, 38923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulz, K.; Hunger, S.; Brown, G.G.; Tsai, S.M.; Cerri, C.C.; Conrad, R.; Drake, H.L. Methanogenic food web in the gut contents of methane-emitting earthworm Eudrilus eugeniae from Brazil. ISME J. 2015, 9, 1778–1792. [Google Scholar] [CrossRef]
- Horn, M.A.; Schramm, A.; Drake, H.L. The earthworm gut: An ideal habitat for ingested N2O-producing microorganisms. Appl. Environ. Microbiol. 2003, 69, 1662–1669. [Google Scholar] [CrossRef] [Green Version]
- Kuiper, I.; De Deyn, G.B.; Thakur, M.P.; van Groenigen, J.W. Soil invertebrate fauna affect N2O emissions from soil. Glob. Chang. Biol. 2013, 19, 2814–2825. [Google Scholar] [CrossRef]
- Zhou, M.; Zhu, B.; Brüggemann, N.; Bergmann, J.; Wang, Y.; Butterbach-Bahl, K. N2O and CH4 emissions, and NO3– leaching on a crop-yield basis from a subtropical rain-fed wheat-maize rotation in response to different types of nitrogen fertilizer. Ecosystems 2014, 17, 286–301. [Google Scholar] [CrossRef]
- Wu, H.; Lu, M.; Lu, X.; Guan, Q.; He, X. Interactions between earthworms and mesofauna has no significant effect on emissions of CO2 and N2O from soil. Soil Biol. Biochem. 2015, 88, 294–297. [Google Scholar] [CrossRef]
- Kharin, S.A.; Kurakov, A.V. Transformation of nitrogen compounds and dynamics of microbial biomass in fresh casts of Aporrectodea caliginosa. Eurasian Soil Sci. 2009, 42, 75–81. [Google Scholar] [CrossRef]
- Amador, J.A.; Görres, J.H. Microbiological characterization of the structures built by earthworms and ants in an agricultural field. Soil Biol. Biochem. 2007, 39, 2070–2077. [Google Scholar] [CrossRef]
- Majeed, M.Z.; Miambi, E.; Barois, I.; Blanchart, E.; Brauman, A. Emissions of nitrous oxide from casts of tropical earthworms belonging to different ecological categories. Pedobiologia 2013, 56, 49–58. [Google Scholar] [CrossRef]
- Mori, A.; Hojito, M. Effect of combined application of manure and fertilizer on N2O fluxes from a grassland soil in Nasu, Japan. Agric. Ecosyst. Environ. 2012, 160, 40–50. [Google Scholar] [CrossRef]
- Paul, B.K.; Lubbers, I.M.; Groenigen, J.W. Residue incorporation depth is a controlling factor of earthworm-induced nitrous oxide emissions. Glob. Chang. Biol. 2011, 18, 1141–1151. [Google Scholar] [CrossRef]
- Linton, N.F.; Ferrari Machado, P.V.; Deen, B.; Wagner-Riddle, C.; Dunfield, K. Long-term diverse rotation alters nitrogen cycling bacterial groups and nitrous oxide emissions after nitrogen fertilization. Soil Biol. Biochem. 2020, 149, 107917. [Google Scholar] [CrossRef]
- Clark, I.M.; Buchkina, N.; Jhurreea, D.; Goulding, K.W.T.; Hirsch, P.R. Impacts of nitrogen application rates on the activity and diversity of denitrifying bacteria in the Broadbalk Wheat Experiment. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 1235–1244. [Google Scholar] [CrossRef] [Green Version]
- Jones, C.M.; Spor, A.; Brennan, F.P.; Breuil, M.-C.; Bru, D.; Lemanceau, P.; Griths, B.; Hallin, S.; Philippot, L. Recently identified microbial guild mediates soil N2O sink capacity. Nat. Clim. Chang. 2014, 4, 801–805. [Google Scholar] [CrossRef]
- Philippot, L.; Andert, J.; Jones, C.M.; Bru, D.; Hallin, S. Importance of denitrifiers lacking the genes encoding the nitrous oxide reductase for N2O emissions from soil. Glob. Chang. Biol. 2010, 17, 1497–1504. [Google Scholar] [CrossRef]
- Levy-Booth, D.J.; Prescott, C.E.; Grayston, S.J. Microbial functional genes involved in nitrogen fixation, nitrification and denitrification in forest ecosystems. Soil Biol. Biochem. 2014, 75, 11–25. [Google Scholar] [CrossRef]
- Hu, H.-W.; Chen, D.; He, J.-Z. Microbial regulation of terrestrial nitrous oxide formation: Understanding the biological pathways for prediction of emission rates. FEMS Microbiol. Rev. 2015, 39, 729–749. [Google Scholar] [CrossRef]
- Zumft, W.G. Cell biology and molecular basis of denitrification. Microbiol. Mol. Biol. Rev. 1997, 61, 533–616. [Google Scholar]
- Coyotzi, S.; Doxey, A.C.; Clark, I.D.; Lapen, D.R.; van Cappellen, P.; Neufeld, J.D. Agricultural soil denitrifiers possess extensive nitrite reductase gene diversity. Environ. Microbiol. 2017, 19, 1189–1208. [Google Scholar] [CrossRef]
- Prieme, A.; Braker, G.; Tiedje, J.M. Diversity of nitrite reductase (nirK and nirS) gene fragments in forested upland and wetland soils. Appl. Environ. Microbiol. 2002, 68, 1893–1900. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, Y.; Evans, S.E.; Friesen, M.L.; Tiemann, L.K. Effect of nitrogen fertilization on the abundance of nitrogen cycling genes in agricultural soils: A meta-analysis of field studies. Soil Biol. Biochem. 2018, 127, 71–78. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Value * |
|---|---|
| Ctot, % | 2.0 ± 0.2 |
| Ntot, % | 0.3 ± 0.1 |
| C:N | 7.8 ± 0.5 |
| pHH2O | 6.0 ± 0.2 |
| Density, g cm−3 | 1.15 ± 0.05 |
| Primer Name | Primer Sequence | Thermal Conditions | Reference |
|---|---|---|---|
| nirK876 nirK1040 | ATY GGC GGV CAY GGC GA GCC TCG ATC AGR TTR TGG TT | 95 °C, 15 min, 1 cycle 95 °C for 15 s, 63 to 58 °C for 30 s (−1 °C by cycle), 72 °C for 30 s, 80 °C for 15 s, 6 cycles 95 °C for 15 s, 60 °C for 30 s, 72 °C for 30 s, 80 °C for 15 s, 40 cycles, 95 °C for 15 s, 60 to 95 °C, 1 cycle | [43] |
| nirSCd3aFm nirSR3cdm | AAC GYS AAG GAR ACS GG GAS TTC GGR TGS GTC TTS AYG AA | 95 °C, 15 min, 1 cycle 95 °C for 15 s, 65 to 60 °C for 30 s (−1 °C by cycle), 72 °C for 30 s, 80 °C for 15 s, 6 cycles 95 °C for 15 s, 60 °C for 30 s, 72 °C for 30 s, 80 °C for 15 s, 40 cycles 95 °C for 15 s, 60 to 95 °C, 1 cycle | [44] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maslov, M.; Astaykina, A.; Pozdnyakov, L. Earthworm Lumbricus terrestris Contributes Nitrous Oxide Emission from Temperate Agricultural Soil Regardless of Applied Mineral Nitrogen Fertilizer Doses. Agronomy 2022, 12, 2745. https://doi.org/10.3390/agronomy12112745
Maslov M, Astaykina A, Pozdnyakov L. Earthworm Lumbricus terrestris Contributes Nitrous Oxide Emission from Temperate Agricultural Soil Regardless of Applied Mineral Nitrogen Fertilizer Doses. Agronomy. 2022; 12(11):2745. https://doi.org/10.3390/agronomy12112745
Chicago/Turabian StyleMaslov, Mikhail, Angelika Astaykina, and Lev Pozdnyakov. 2022. "Earthworm Lumbricus terrestris Contributes Nitrous Oxide Emission from Temperate Agricultural Soil Regardless of Applied Mineral Nitrogen Fertilizer Doses" Agronomy 12, no. 11: 2745. https://doi.org/10.3390/agronomy12112745