
Genotypic and Environmental Influence on Fresh Rhizome Yield of Turmeric (Curcuma longa L.)
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Waimanalo
2.2. Kaua‘i
2.3. Maui
2.3.1. Kula Agricultural Park
2.3.2. Maui Agricultural Research Center
2.4. Statistical Analysis
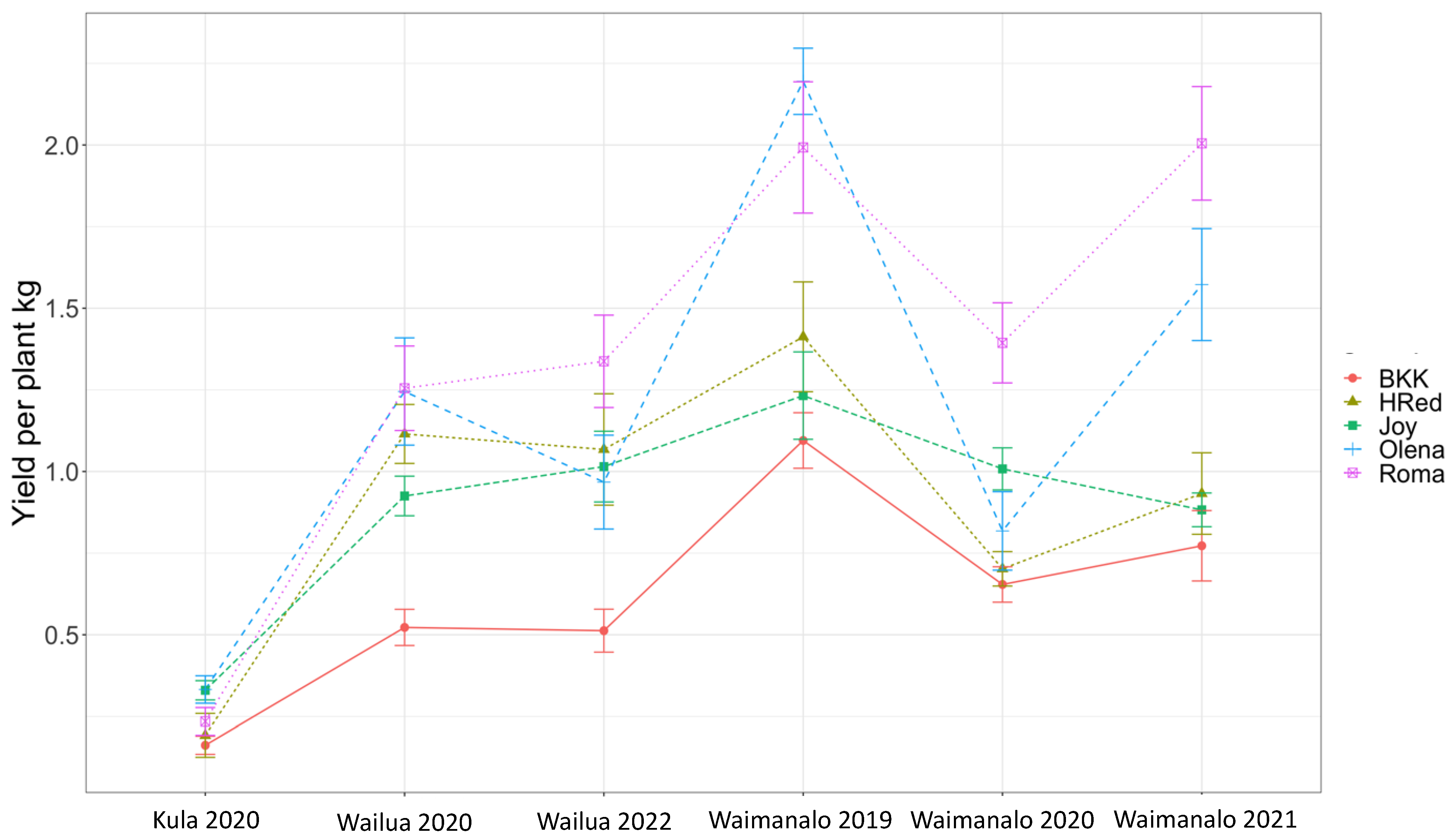
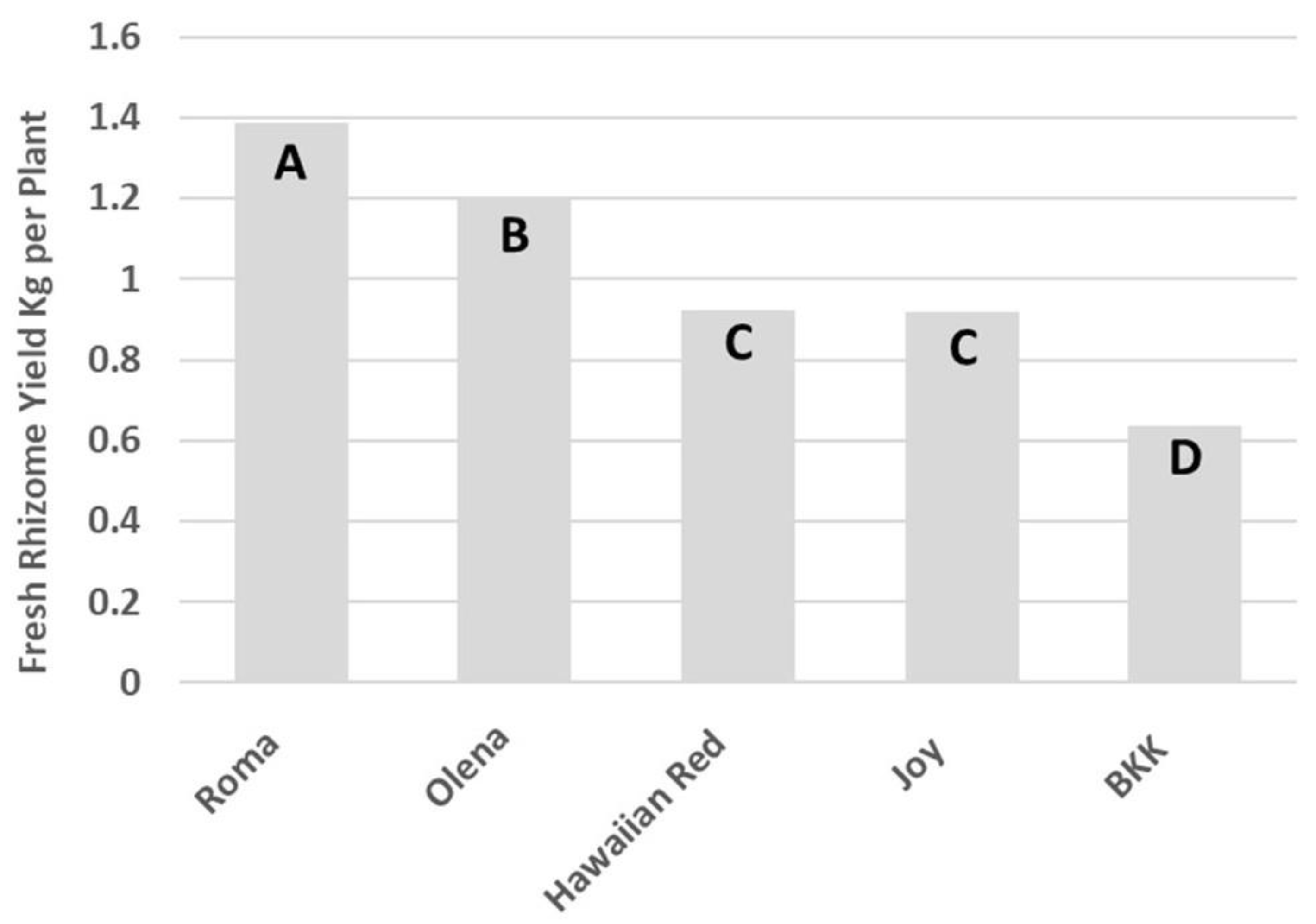
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nair, K.P. Turmeric (Curcuma longa L.) and Ginger (Zingiber officinale Rosc.)-World’s Invaluable Medicinal Spices: The Agronomy and Economy of Turmeric and Ginger; Springer: Cham, Switzerland, 2019. [Google Scholar]
- Prasath, D.; Kandiannan, K.; Leela, N.K.; Aarthi, S.; Sasikumar, B.; Babu, K.N. Turmeric: Botany and production practices. Hortic. Rev. 2019, 46, 99–184. [Google Scholar]
- Choo, B.K.M.; Shaikh, M.F. Mechanism of Curcuma longa and Its Neuroactive Components for the Management of Epileptic Seizures: A Systematic Review. Curr. Neuropharmacol. 2021, 19, 1496–1518. [Google Scholar] [CrossRef] [PubMed]
- Fuloria, S.; Mehta, J.; Chandel, A.; Sekar, M.; Rani, N.; Begum, M.Y.; Subramaniyan, V.; Chidambaram, K.; Thangavelu, L.; Nordin, R.; et al. A Comprehensive Review on the Therapeutic Potential of Curcuma longa Linn. in Relation to its Major Active Constituent Curcumin. Front. Pharmacol. 2022, 13, 820806. [Google Scholar] [CrossRef] [PubMed]
- Statista. Available online: https://www.statista.com/statistics/798314/us-turmeric-imports/ (accessed on 22 August 2022).
- Flores, S.; Retana-Cordero, M.; Fisher, P.R.; Freyre, R.; Gomez, C. Effect of Photoperiod, Propagative Material, and Production Period on Greenhouse-grown Ginger and Turmeric Plants. Hortscience 2021, 56, 1476–1485. [Google Scholar] [CrossRef]
- Retana-Cordero, M.; Flores, S.; Freyre, R.; Gomez, C. Strategies to Reduce Radiation Stress in Open-Field Ginger and Turmeric Production. Agronomy 2022, 12, 1910. [Google Scholar] [CrossRef]
- Retana-Cordero, M.; Fisher, P.R.; Gomez, C. Modeling the Effect of Temperature on Ginger and Turmeric Rhizome Sprouting. Agronomy 2021, 11, 1931. [Google Scholar] [CrossRef]
- Shannon, D.A.; van Santen, E.; Salmasi, S.Z.; Murray, T.J.; Duong, L.T.; Greenfield, J.T.; Gonzales, T.; Foshee, W. Shade, Establishment Method, and Varietal Effects on Rhizome Yield and Curcumin Content in Turmeric in Alabama. Crop Sci. 2019, 59, 2701–2710. [Google Scholar] [CrossRef]
- Radovich, T.J.K. Biology and Classification of Vegetables; John Wiley & Sons, Ltd.: Chichester, UK, 2018; pp. 1–23. [Google Scholar]
- Ravindran, P.N.; Nirmal Babu, K.; Sivaraman, K. Turmeric: The Genus Curcuma; CRC Press: Boca Raton, FL, USA, 2007. [Google Scholar]
- Vinodhini, V.; Selvi, B.S.; Balakrishnan, S.; Muthuragavan, R. Studies on variability and genetic components of yield and quality traits in turmeric (Curcuma longa L.). Electron. J. Plant Breed. 2018, 9, 1060–1066. [Google Scholar] [CrossRef]
- Vimal, V.K.; Singh, P.K.; Pandey, V.P. Assess the genetic diversity for growth yield and quality characters among the genotypes of turmeric. Plant Arch. 2018, 18, 1026–1032. [Google Scholar]
- Rivière, P.; Dawson, J.C.; Goldringer, I.; David, O. Hierarchical Bayesian Modeling for Flexible Experiments in Decentralized Participatory Plant Breeding. Crop Sci. 2015, 55, 1053–1067. [Google Scholar] [CrossRef]
- Chitra, C.; Daniel, G.M.; Ponmurugan, P.; Mathivanan, N. Association of multiple fungal pathogens to rhizome rot disease of turmeric. Arch. Phytopathol. Plant Prot. 2020, 53, 440–458. [Google Scholar] [CrossRef]
- Hall, M.; Lawrence, K.; Shannon, D.; Gonzalez, T.; Newman, M. First report of binucleate Rhizoctonia AG-G on common turmeric (Curcuma longa) in the United States. Plant Dis. 2019, 103, 771. [Google Scholar] [CrossRef]
- Vinayarani, G.; Prakash, H.S. Fungal endophytes of turmeric (Curcuma longa L.) and their biocontrol potential against pathogens Pythium aphanidermatum and Rhizoctonia solani. World J. Microbiol. Biotechnol. 2018, 34, 49. [Google Scholar] [CrossRef]
- Amadi, C.; Olojede, A.O.; Obasi, M. Growth and yield of turmeric in a derived savanna agro-ecology of Nigeria. Int. J. Agric. Policy Res. 2015, 3, 388–395. [Google Scholar]
- Choudhary, A.K.; Rahi, S. Organic cultivation of high yielding turmeric (Curcuma longa L.) cultivars: A viable alternative to enhance rhizome productivity, profitability, quality and resource-use efficiency in monkey-menace areas of north-western Himalayas. Ind. Crops Prod. 2018, 124, 495–504. [Google Scholar] [CrossRef]
- Harish, B.S.; Umesha, K.; Venugopalan, R.; Prasad, B.N.M. Photo-selective nets influence physiology, growth, yield and quality of turmeric (Curcuma longa L.). Ind. Crops Prod. 2022, 186, 115202. [Google Scholar] [CrossRef]
- Bahadur, M.; Azad, A.; Hakim, M.; Hossain, S.; Sikder, S. Effect of different spacing and potassium levels on the growth and yield of turmeric var. sinduri. Pak. J. Biol. Sci. 2000, 3, 593–595. [Google Scholar] [CrossRef] [Green Version]
- Patel, B.N.; Patel, R.K.; Bhalerao, P.P. Effect of rhizome size on growth and yield of turmeric CV. GNT-1. Int. J. Trop. Agric. 2018, 36, 771–777. [Google Scholar]
- Vithya, K.; Venkatesan, K.; Selvi, B.S.; Manonmani, S.; Kokiladevi, E. Per se performance and diversity analysis in turmeric (Curcuma longa L.) genotypes for plant and rhizome yield characters. Electron. J. Plant Breed. 2021, 12, 1314–1320. [Google Scholar] [CrossRef]
- Sivakumar, V.; Chandrasekar, R.C.; Bhagavan, B.V.K.; Ravindrakumar, K. Genetic variability, heritability and genetic advance studies in turmeric (Curcuma longa L.) under high altitudearea of Andhra Pradesh. Environ. Ecol. 2021, 39, 697–701. [Google Scholar]
- Mohamed, M.; Mahfouz, S.; Yosef, A. Effect of spacing and varieties on growth, yield and chemical constituents of Turmeric plants. Int. J. Med. Aromat. Plants 2014, 4, 34–40. [Google Scholar]
- Aswathi, A.P.; Raghav, S.B.; Prasath, D. Assessment of genetic variation in turmeric (Curcuma longa L.) varieties based on morphological and molecular characterization. Genet. Resour. Crop Evol. 2022, 1–12. [Google Scholar] [CrossRef]
- Suresh, R.; Ramar, A.; Balakrishnan, S.; Rajeswari, S.; Kumaravadivel, N. Performance and evaluation of turmeric (Curcuma longa L.) genotypes based on quantitative traits for tropical regions of Tamil Nadu. Electron. J. Plant Breed. 2020, 11, 735–741. [Google Scholar] [CrossRef]
- Vinodhini, V.; Selvi, B.S.; Balakrishnan, S.; Suresh, R. Studies on growth analysis and framing selection criteria for high yield and quality traits in turmeric (Curcuma longa L.). Electron. J. Plant Breed. 2022, 13, 519–525. [Google Scholar] [CrossRef]
- Mekonnen, B.; Garedew, W. Growth, yield, and quality responses of turmeric (Curcuma longa L.) to nitrogen fertilizer rate and timing of its application. Acta Agrobot. 2019, 72, 1–11. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Harvest Year | Location | Cultivar | Fingers | Mothers | Estimated Finger Yield |
|---|---|---|---|---|---|
| kg plant−1 | t ha−1 | ||||
| 2019 | Maui Ag. Park | BKK | 1.03 b | 0.30 b | 22.89 b |
| Hawaiian Red | 1.65 a | 0.47 a | 36.59 a | ||
| Joy | 1.58 a | 0.53 a | 35.18 a | ||
| Mystic | 1.50 a | 0.50 a | 33.33 a | ||
| Olena | 1.37 ab | 0.30 b | 30.37 ab | ||
| 2019 | Waimānalo | BKK | 1.10 b | 0.11 c | 24.36 b |
| Caribbean | 1.15 b | 0.27 b | 25.64 b | ||
| Hawaiian Red | 1.41 b | 0.28 b | 31.35 b | ||
| Joy | 1.23 b | 0.33 b | 27.41 b | ||
| Mystic | 1.22 b | 0.35 ab | 27.17 b | ||
| Olena | 2.19 a | 0.33 b | 48.77 a | ||
| Roma | 1.99 a | 0.46 a | 44.23 a | ||
| 2020 | Kaua‘i | BKK | 0.52 d | 0.18 d | 11.61 d |
| Hawaiian Red | 1.12 ab | 0.31 bc | 24.78 ab | ||
| Joy | 0.93 bc | 0.39 ab | 20.56 bc | ||
| Olena | 1.25 a | 0.22 cd | 27.67 a | ||
| Roma | 1.26 a | 0.42 a | 27.89 a | ||
| Wailua Gold | 0.70 cd | 0.17 d | 15.50 cd | ||
| 2020 | MARC | BKK | 0.16 b | 0.07 c | 3.64 b |
| Hawaiian Red | 0.26 ab | 0.13 b | 5.69 ab | ||
| Joy | 0.33 a | 0.17 a | 7.29 a | ||
| Olena | 0.33 a | 0.08 c | 7.35 a | ||
| Roma | 0.23 b | 0.05 c | 5.20 b | ||
| 2020 | Waimānalo | 18-007 | 1.13 bcd | 0.27 ab | 25.11 bcd |
| 18-008 | 1.19 bc | 0.23 abc | 26.53 bc | ||
| 18-012 | 0.83 cd | 0.21 bc | 18.44 cd | ||
| 18-013 | 1.32 b | 0.21 bc | 29.33 b | ||
| 18-014 | 1.12 bcd | 0.20 c | 24.80 bcd | ||
| BKK | 0.80 d | 0.19 c | 17.87 d | ||
| Hawaiian Red | 0.97 bcd | 0.28 a | 21.64 bcd | ||
| Joy | 1.29 b | 0.22 abc | 28.58 b | ||
| Olena | 1.10 bcd | 0.27 a | 24.40 bcd | ||
| Roma | 1.87 a | 0.24 abc | 41.51 a | ||
| 2021 | Waimānalo | 18-007 | 1.48 bcd | 0.24 ab | 32.89 bcd |
| 18-008 | 1.20 d | 0.22 bc | 26.61 d | ||
| 18-013 | 1.50 bcd | 0.27 a | 33.33 bcd | ||
| 18-014 | 1.98 b | 0.20 bcd | 43.89 b | ||
| 18-023 | 1.36 cd | 0.18 cd | 30.11 cd | ||
| BKK | 0.92 d | 0.16 de | 20.39 d | ||
| Hawaiian Red | 1.14 d | 0.18 cd | 25.33 d | ||
| Joy | 1.10 d | 0.20 bcd | 24.39 d | ||
| Olena | 1.80 bc | 0.13 e | 40.06 bc | ||
| Roma | 2.58 a | 0.22 abc | 57.22 a | ||
| 2022 | Kaua‘i | 18-013 | 1.48 a | 0.36 a | 32.89 a |
| 18-023 | 1.26 ab | 0.28 ab | 27.89 ab | ||
| BKK | 0.51 d | 0.09 e | 11.39 d | ||
| Hawaiian Red | 1.07 bc | 0.17 cde | 23.72 bc | ||
| Joy | 1.02 bc | 0.20 bcd | 22.56 bc | ||
| Olena | 0.97 c | 0.13 de | 21.50 c | ||
| Roma | 1.34 a | 0.26b c | 29.72 a | ||
| Wailua Gold | 0.97 d | 0.11 e | 21.50 d | ||
| Iteration (I) | p < 0.001 | p < 0.001 | p < 0.001 | ||
| Genotype (G) | p < 0.001 | p < 0.001 | p < 0.001 | ||
| I × G | p < 0.001 | p < 0.001 | p < 0.001 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tavares, K.; Kirk, E.; Motomura-Wages, S.; Calpito, J.; Bingham, J.-P.; Ahmad, A.A.; Flanagan, K.; Uyeda, J.; Kantar, M.B.; Radovich, T.J.K. Genotypic and Environmental Influence on Fresh Rhizome Yield of Turmeric (Curcuma longa L.). Agronomy 2022, 12, 2703. https://doi.org/10.3390/agronomy12112703
Tavares K, Kirk E, Motomura-Wages S, Calpito J, Bingham J-P, Ahmad AA, Flanagan K, Uyeda J, Kantar MB, Radovich TJK. Genotypic and Environmental Influence on Fresh Rhizome Yield of Turmeric (Curcuma longa L.). Agronomy. 2022; 12(11):2703. https://doi.org/10.3390/agronomy12112703
Chicago/Turabian StyleTavares, Kylie, Emilie Kirk, Sharon Motomura-Wages, Justin Calpito, Jon-Paul Bingham, Amjad A. Ahmad, Kevin Flanagan, Jensen Uyeda, Michael B. Kantar, and Theodore J. K. Radovich. 2022. "Genotypic and Environmental Influence on Fresh Rhizome Yield of Turmeric (Curcuma longa L.)" Agronomy 12, no. 11: 2703. https://doi.org/10.3390/agronomy12112703