
Seasonal, Landscape, and Attractant Effects on Lesser Grain Borer, Rhyzopertha dominica (F.), Captures in Northeast Kansas


Abstract
:1. Introduction
2. Materials and Methods
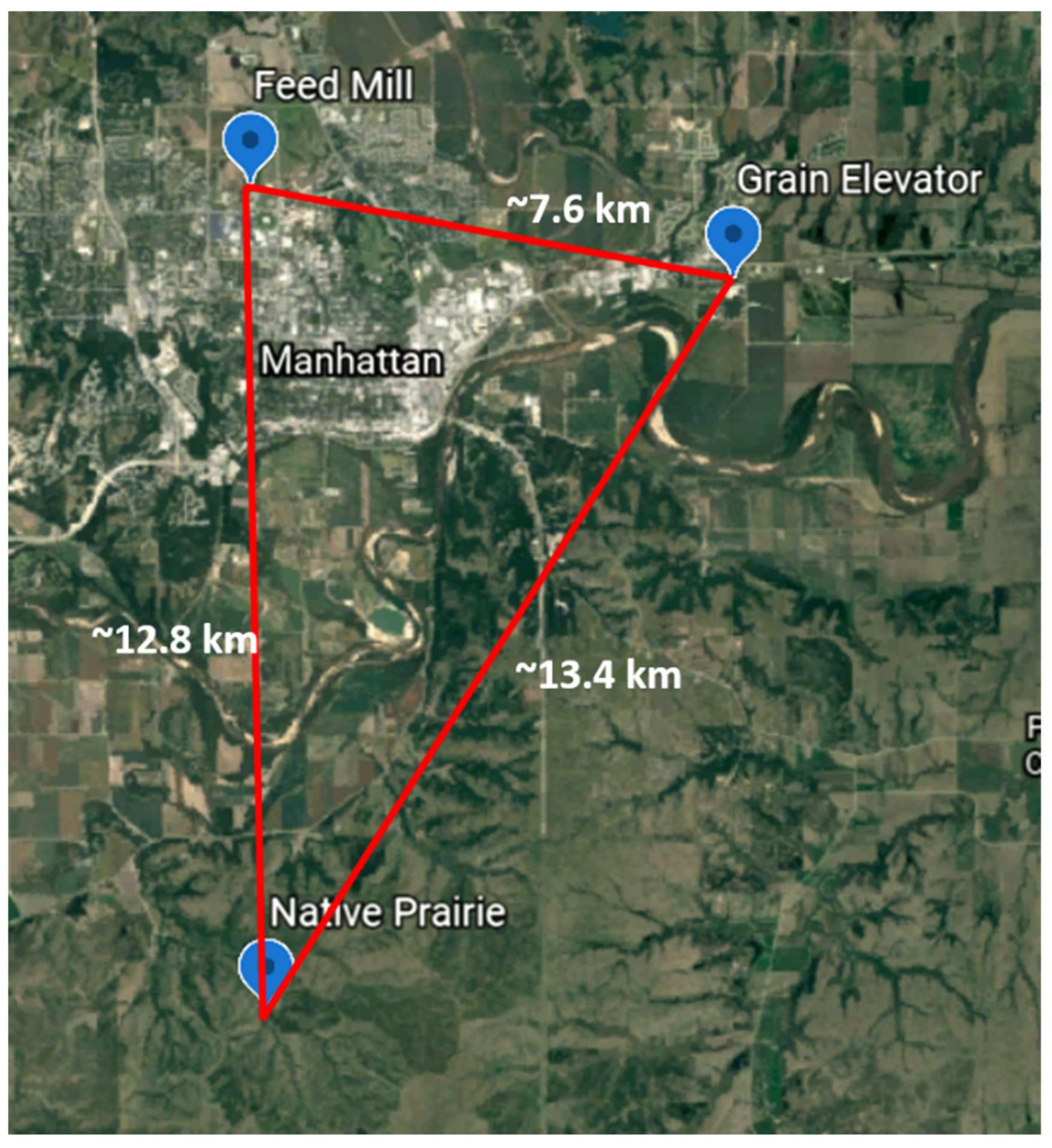
2.1. Study Sites
2.2. Delta Traps
2.3. Wheat Preparation
2.4. Lindgren Traps
3. Results
3.1. Delta Trapping
3.2. Lindgren Trapping
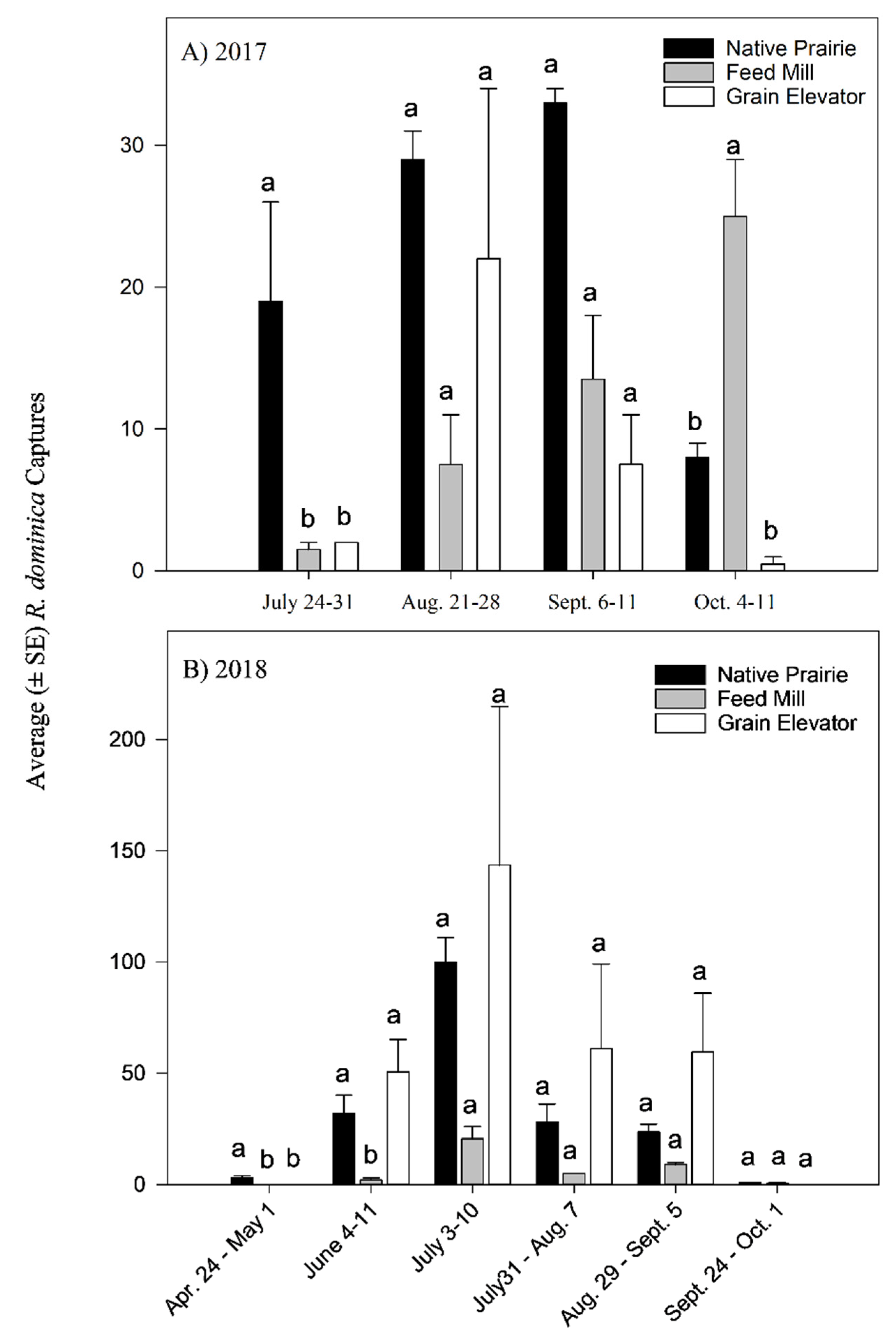
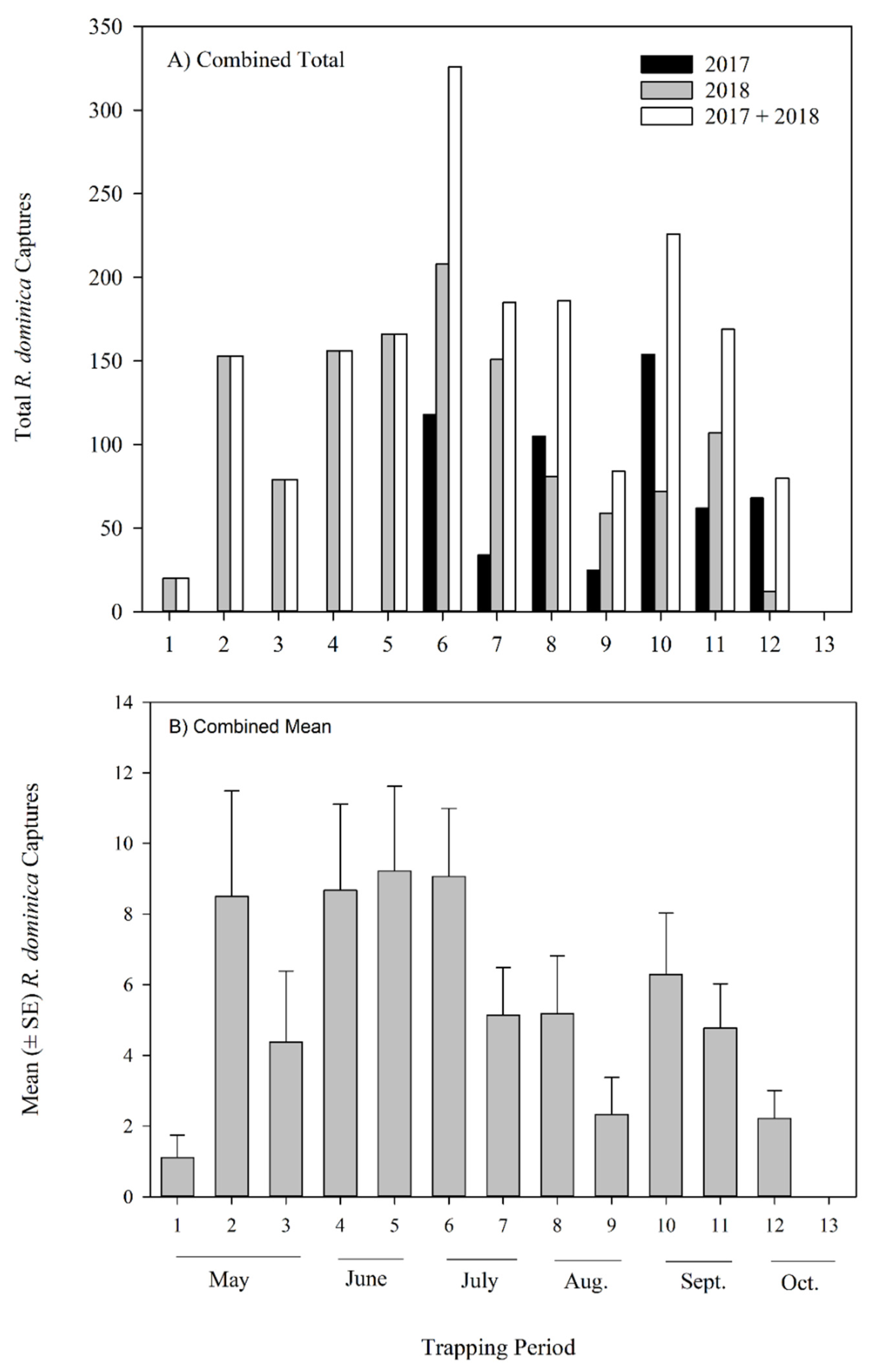
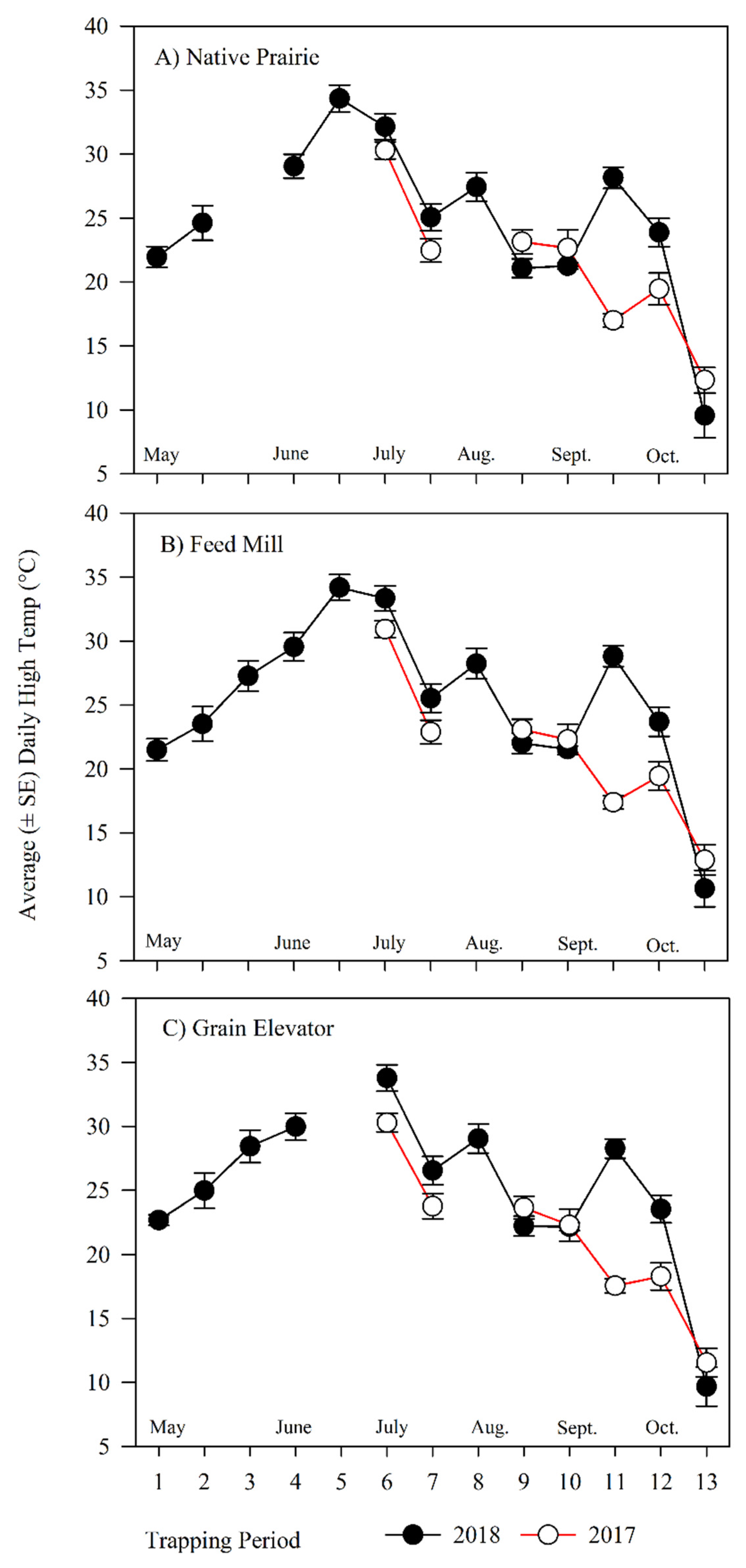
3.2.1. Seasonal Effect on Rhyzopertha dominica Captures
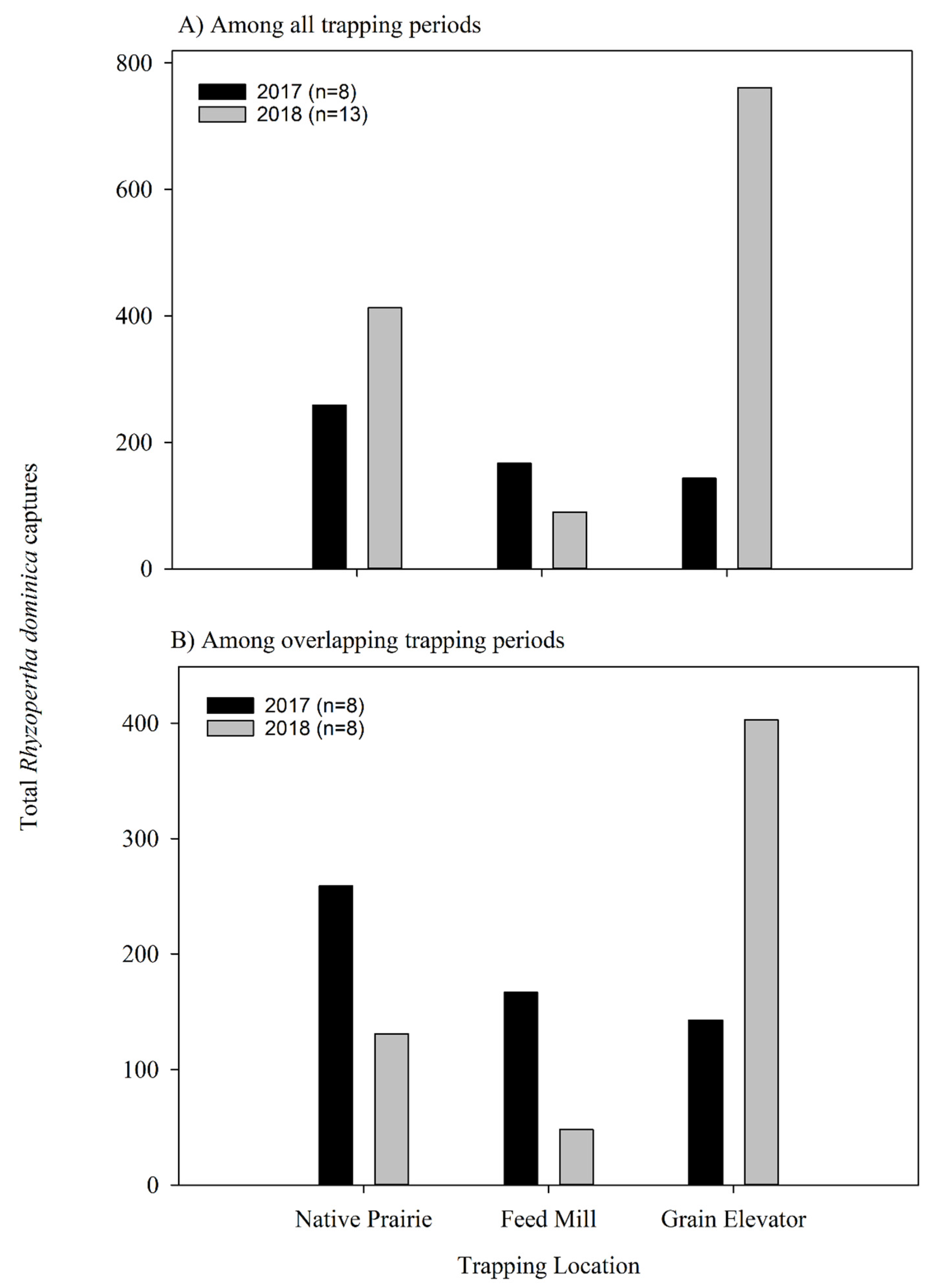
3.2.2. Influence of Ecological Landscape on Rhyzopertha dominica Captures
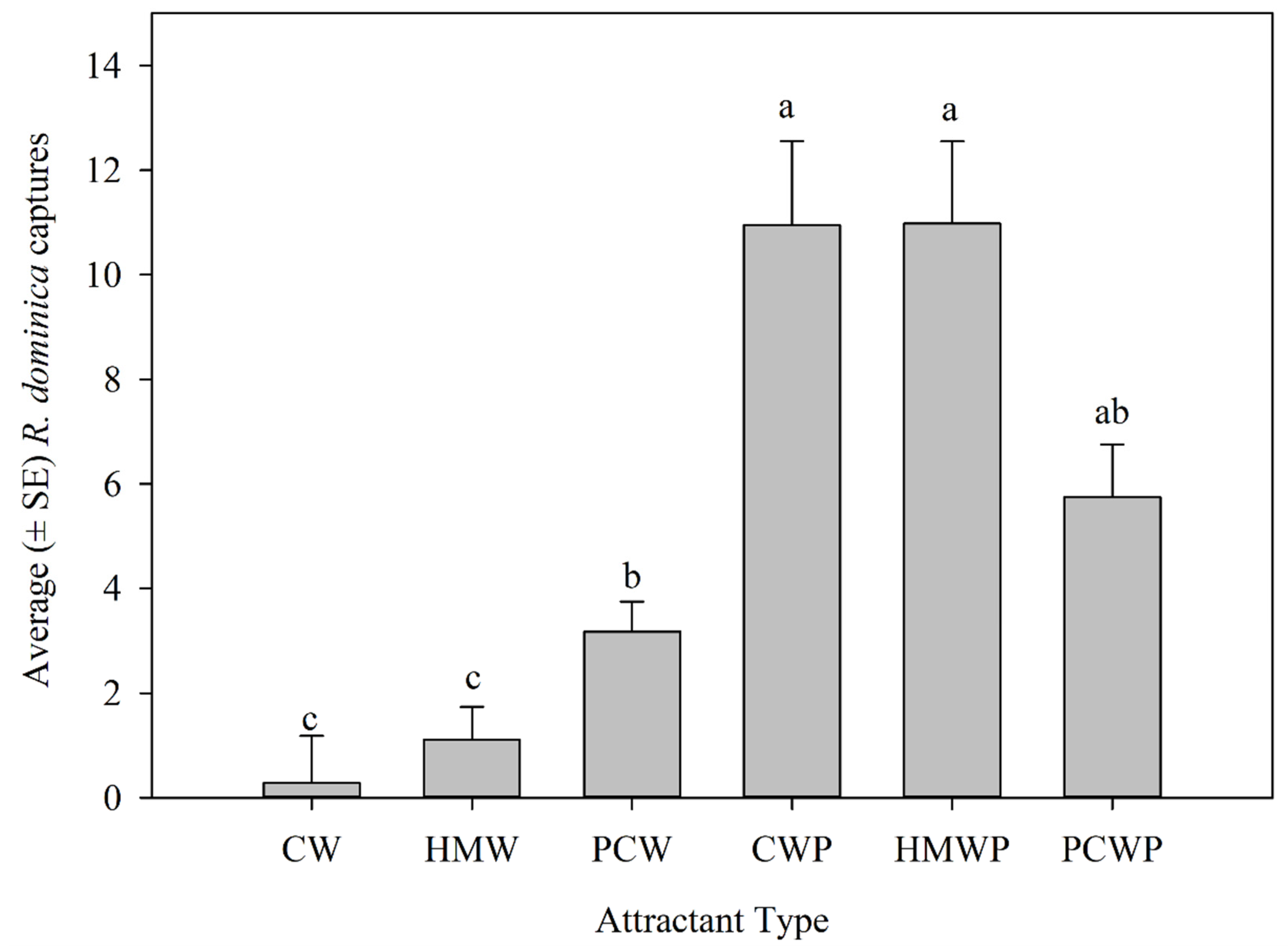
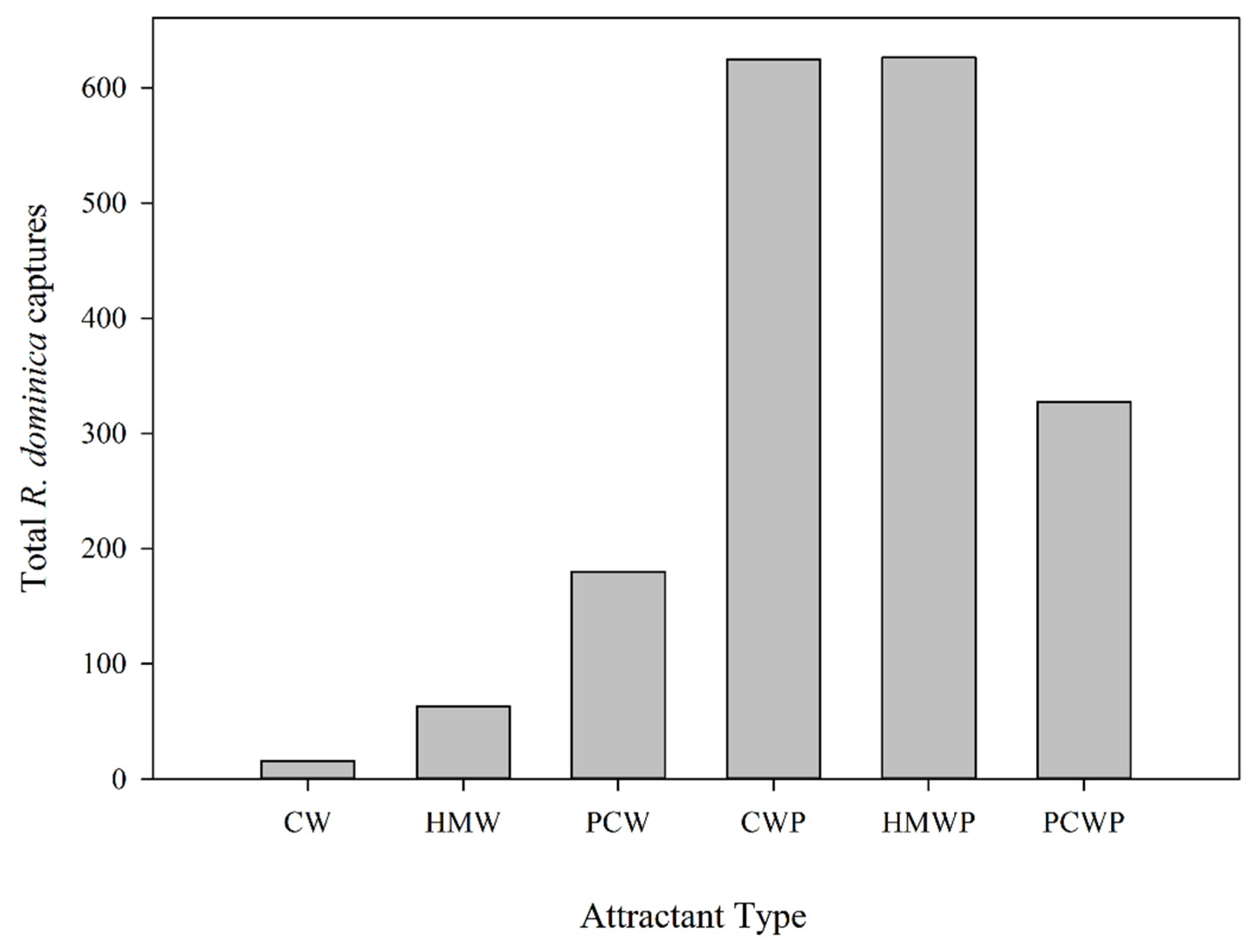
3.2.3. Effect of Attractant Type on Rhyzopertha dominica Captures
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Potter, C. The biology and distribution of Rhyzopertha dominica (Fab.). Trans. R. Ent. Soc. Lond. 1935, 83, 449–482. [Google Scholar] [CrossRef]
- Hagstrum, D.W. Infestation by Cryptolestes ferrugineus (Coleoptera: Cucujidae) of newly harvested wheat stored on three Kansas farms. J. Econ. Entomol. 1989, 82, 655–659. [Google Scholar] [CrossRef]
- Mahroof, R.M.; Edde, P.A.; Robertson, B.; Puckette, J.A.; Phillips, T.W. Dispersal of Rhyzopertha dominica (Coleoptera: Bostrichidae) in different habitats. Environ. Entomol. 2010, 39, 930–938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, H.J.; Silverstein, R.M.; Burkholder, W.E.; Khorramshahi, A. Dominicalure 1 and 2: Components of aggregation pheromone from male lesser grain borer Rhyzopertha dominica (F.) (Coleoptera: Tenebrionidae). J. Chem. Ecol. 1981, 7, 759–781. [Google Scholar] [CrossRef] [PubMed]
- Birch, L.C. The mortality of the immature stages of Calandra oryzae L. (small strain) and Rhyzopertha dominica Fab. in wheat of different moisture contents. Aust. J. Exp. Biol. Med. Sci. 1945, 23, 141–145. [Google Scholar] [CrossRef]
- Dowdy, A.K.; McGughey, W.H. Seasonal activity of stored-product insects in and around farm-stored wheat. J. Econ. Entomol. 1994, 87, 1351–1358. [Google Scholar] [CrossRef]
- Wright, J.E.; Morton, R. Daily flight activity of Trogoderma variabile (Coleoptera: Dermestidae) and Rhyzopertha dominica (Coleoptera: Bostrichidae). J. Stored Prod. Res. 1995, 31, 177–184. [Google Scholar] [CrossRef]
- Cox, P.D.; Wakefield, M.E.; Jacob, T.A. The effects of temperature on flight initiation in a range of moths, beetles, and parasitoids associated with stored products. J. Stored Prod. Res. 2007, 43, 111–117. [Google Scholar] [CrossRef]
- Edde, P.A.; Phillips, T.W.; Nansen, C.; Payton, M.E. Flight activity of the lesser grain borer, Rhyzopertha dominica F. (Coleoptera: Bostrichidae), in relation to weather. Environ. Entomol. 2006, 35, 616–624. [Google Scholar] [CrossRef] [Green Version]
- Leos-Martinez, J.; Granovsky, T.A.; Williams, H.J.; Vinson, B.; Burkholder, W.E. Estimation of aerial density of the lesser grain borer (Coleoptera: Bostrichidae) in a warehouse using dominicalure traps. J. Econ. Entomol. 1986, 79, 1134–1138. [Google Scholar] [CrossRef]
- Barrer, P.M.; Starick, N.T.; Morton, R.; Wright, J.E. Factors influencing initiation of flight by Rhyzopertha dominica (F.) (Coleoptera: Bostrichidae). J. Stored Prod. Res. 1993, 29, 1–5. [Google Scholar] [CrossRef]
- Aslam, M.; Hagstrum, D.W.; Bover, B.A. The effect of phtoperiod on the flight activity and biology of Rhyzopertha dominica (Coleoptera: Bostrichidae). J. Kans. Entomol. Soc. 1994, 67, 107–115. [Google Scholar]
- Dowdy, A.K. Flight initiation of lesser grain borer (Coleoptera: Bostrichidae) as influenced by temperature, humidity, and light. J. Econ. Entomol. 1994, 87, 1714–1717. [Google Scholar] [CrossRef]
- Perez-Mendoza, J.; Dover, D.D.; Hagstrum, D.W.; Baker, J.E. Flight activity of Rhyzopertha dominica (Coleoptera: Bostrichidae) in response to feeding damage and accumulation of waste. J. Econ. Entomol. 1998, 91, 1445–1448. [Google Scholar] [CrossRef]
- Perez-Mendoza, J.; Dover, B.A.; Hagstrum, D.W.; Hopkins, T.L. Effect of crowding, food deprivation, and diet on flight initiation and lipid reserves of the lesser grain borer, Rhyzopertha dominica. Entomol. Exp. Appl. 1999, 91, 317–326. [Google Scholar] [CrossRef] [Green Version]
- Perez-Mendoza, J.; Hagstrum, D.W.; Dover, B.A.; Hopkins, T.L.; Bake, J.E. Flight response, body weight, and lipid content of Rhyzopertha dominica (F.) (Coleoptera: Bostrichidae) as influenced by strain, season, and phenotype. J. Stored Prod. Res. 1999, 35, 183–196. [Google Scholar] [CrossRef]
- Toews, M.D.; Campbell, J.F.; Arthur, F.H.; Ramaswamy, S.B. Outdoor flight activity and immigration of Rhyzopertha dominica into seed wheat warehouses. Entomol. Exp. Appl. 2006, 121, 73–85. [Google Scholar] [CrossRef]
- Throne, J.E.; Cline, D.L. Seasonal flight activity and seasonal abundance of selected stored-product Coleoptera around grain storages in South Carolina. J. Agric. Entomol. 1994, 11, 321–338. [Google Scholar]
- Sinclair, E.R.; Haddrell, R.L. Flight of stored products beetles over a grain farming area in southern Queensland. J. Aust. Ent. Soc. 1985, 24, 9–15. [Google Scholar] [CrossRef]
- Ridley, A.W.; Hereward, J.P.; Daglish, G.J.; Raghu, S.; McCulloch, G.A.; Walter, G.H. Flight of Rhyzopertha dominica (Coleoptera: Bostrichidae)—a spatio-temporal analysis with pheromone trapping and population genetic. J. Econ. Entomol. 2016, 109, 2561–2571. [Google Scholar] [CrossRef] [Green Version]
- Daglish, G.J.; Ridley, A.W.; Reid, R.; Walter, G.H. Testing the consistency of spatio-temporal patterns of flight activity in the stored grains beetles Tribolium castaneum (Herbst) and Rhyzopertha dominica (F.). J. Stored Prod. Res. 2017, 72, 68–74. [Google Scholar] [CrossRef]
- Fields, P.G.; Van Loon, J.; Donlinski, M.G.; Harris, J.L.; Burkholder, W.E. The distribution of Rhyzopertha dominica (F.) in Western Canada. Can. Entomol. 1993, 125, 317–328. [Google Scholar] [CrossRef]
- Rajan, S.T.; Muralitharan, V.; Daglish, G.J.; Mohankumar, S.; Rafter, M.A.; Chandrasekaran, S.; Mohan, S.; Vimal, D.; Srivastava, C.; Loganathan, M.; et al. Flight of three major insect pests of stored grain in the monsoonal tropics of India, by latitude, season, and habitat. J. Stored Prod. Res. 2018, 76, 43–50. [Google Scholar] [CrossRef]
- Buonocore, E.; Monaco, D.L.; Russo, A.; Aberlenc, H.P.; Garzia, G.T. Rhyzopertha dominica (F., 1792) (Coleoptera: Bostrichidae): A stored grain pest on olive trees in Sicily. Bull OEPP/EPPO 2017, 47, 263–268. [Google Scholar] [CrossRef]
- McKay, T.; White, A.L.; Starkus, L.A.; Arthur, F.H.; Campbell, J.F. Seasonal patterns of stored-product insects at a rice mill. J. Econ. Entomol. 2017, 110, 1366–1376. [Google Scholar] [CrossRef]
- Ching’oma, G.P. Spatial Distribution and Movement of the Lesser Grain Borer, Rhyzopertha dominica F. Ph.D. Thesis, Kansas State University, Manhattan, KS, USA, 2006. [Google Scholar]
- Mahroof, R.M.; Phillips, T.W. Stable isotopes as markers to investigate host use by Rhyzopertha dominica. Entomol. Exp. Appl. 2007, 125, 205–213. [Google Scholar] [CrossRef]
- Edde, P.A. A review of the biology and control of Rhyzopertha dominica (F.) the lesser grain borer. J. Stored Prod. Res. 2012, 48, 1–18. [Google Scholar] [CrossRef]
- Jia, F.; Toews, M.D.; Campbell, J.F.; Ramaswamy, S.B. Survival and reproduction of lesser grain borer, Rhyzopertha dominica (F.) (Coleoptera: Bostrichidae) on flora associated with native habitats in Kansas. J. Stored Prod. Res. 2008, 44, 366–372. [Google Scholar] [CrossRef]
- Wright, V.F.; Fleming, E.E.; Post, D. Survival of Rhyzopertha dominica (Coleoptera: Bostrichidae) on fruits and seeds collected from woodrat nests in Kansas. J. Kans. Entomol. Soc. 1990, 63, 344–347. [Google Scholar]
- Edde, P.A.; Phillips, T.W. Potential host affinities for the lesser grain borer, Rhyzopertha dominica: Behavioral responses to host odors and pheromones and reproductive ability on non-grain hosts. Entomol. Exp. Appl. 2006, 119, 255–263. [Google Scholar] [CrossRef]
- Zar, J.H. Biostatistical Analysis, 5th ed.; Pearson Prentice-Hall: Upper Saddle River, NJ, USA, 2010. [Google Scholar]
- Cereals & Grains Association. AACC Approved Methods of Analysis, 11th ed.; Method 26-95.01; Experimental Milling: Temper Table; Cereals & Grains Association: St. Paul, MN, USA, 1999. [Google Scholar]
- Nguyen, D.T.; Hodges, R.J.; Belmain, S.R. Do walking Rhyzopertha dominica (F.) locate cereal hosts by chance? J. Stored Prod. Res. 2008, 44, 90–99. [Google Scholar] [CrossRef]
- Lindgren, B.S. A multiple funnel trap for scolytid beetles (Coleoptera). Can. Entomol. 1983, 115, 299–302. [Google Scholar] [CrossRef]
- Edde, P.A.; Phillips, T.W.; Toews, M.D. Responses of Rhyzopertha dominica (Coleoptera: Bostrichidae) to its aggregation pheromones as influenced by trap design, trap height, and habitat. Environ. Entomol. 2005, 34, 1549–1557. [Google Scholar] [CrossRef] [Green Version]
- Hagstrum, D.W. Immigration of insects into bins storing newly harvested wheat on 12 Kansas farms. J. Stored Prod. Res. 2001, 37, 221–229. [Google Scholar] [CrossRef]
- Holloway, J.C.; Daglish, G.J.; Mayer, D.G. Spatial distribution and flight patterns of two grain storage insect pests, Rhyzopertha dominica (Bostrichidae) and Tribolium castaneum (Tenebrionidae): Implications for pest management. Insects 2020, 11, 715. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Field Site Location | Site Description | ||
|---|---|---|---|
| Konza Prairie Biological Research Station (Native Prairie) | Trap 1 | 39°5′30.6″ N; 96°35′15.0″ W | 3487 ha native tallgrass prairie preserve located in the Flint Hills region of Northeast Kansas. Areas, where traps were placed, are grazed by bison and burned yearly. Traps were placed along a dirt roadway near low underbrush. |
| Trap 6 | 39°5′28.9″ N; 96°35′13.5″ W | ||
| KSU Feed Mill (Feed Mill) | Trap 1 | 39°12′23.8″ N; 96°35′25.0″ W | A pilot-scale feed mill was used for training, extension programs, and which produced small lots of animal feed. Traps were placed on the north side of the feed mill near the underbrush. |
| Trap 6 | 39°12′25.4″ N; 96°35′23.5″ W | ||
| Commercial Grain CO-OP Elevator (Grain Elevator) | Trap 1 | 39°11′37.0″ N; 96°30′12.3″ W | Commercial grain cooperative (CO-OP) grain elevator located in Northeast Kansas. Traps were placed between concrete storage silos and an active railroad line. |
| Trap 6 | 39°11′36.7″ N; 96°30′14.3″ W | ||
| Trial Number | Lindgren Trapping Dates | Delta Trapping Dates | ||
|---|---|---|---|---|
| 2017 * | 2018 | 2017 | 2018 | |
| 1 | - | 1–3 May | - | 24 April–1 May |
| 2 | - | 16–18 May | - | - |
| 3 | - | 30 May–1 June | - | - |
| 4 | - | 11–13 June | - | 4–11 June |
| 5 | - | 27–29 June | - | - |
| 6 | 17–19 July | 11–13 July | 24–31 July | 3–10 July |
| 7 | 31 July–2 August | 24–26 July | - | - |
| 8 | 16–18 August | 8–10 August | 21–28 August | 31 July–7 August |
| 9 | 28–30 August | 21–23 August | - | - |
| 10 | 11–13 September | 5–7 September | 6–11 September | 29 August–5 September |
| 11 | 25–28 September | 17–19 September | - | - |
| 12 | 11–13 October | 1–3 October | 4–11 October | 24 September–1 October |
| 13 | 23–25 October | 15–17 October | - | - |
| Trapping Year | Correlation Coefficient | p-Value |
|---|---|---|
| 2017 | 0.561 | 0.1906 |
| 2018 | 0.766 | 0.0022 |
| 2017 and 2018 | 0.729 | 0.0003 |
| Factor | F | df | p |
|---|---|---|---|
| Trap location | 9.44 | 2, 342 | <0.0001 |
| Attractant type | 30.72 | 5, 342 | <0.0001 |
| Year | 0.94 | 1, 342 | 0.33187 |
| Trap location × Attractant type | 1.10 | 10, 342 | 0.3623 |
| Trap location × Year | 9.11 | 2, 342 | 0.0001 |
| Attractant type × Year | 0.37 | 5, 342 | 0.8670 |
| Trap location × Attractant type × Year | 1.77 | 10, 342 | 0.0647 |
| Trapping Location | 2017 (n = 7) | 2018 (n = 12) | ||
|---|---|---|---|---|
| Total Captures | Mean (±SE) Captures a | Total Captures | Mean (±SE) Captures b | |
| Feed Mill | 167 | 23.9 ± 6.3 | 90 | 7.5 ± 2.4 b |
| Native Prairie | 259 | 37.0 ± 13.0 | 431 | 34.4 ± 9.1 a |
| Grain Elevator | 143 | 20.4 ± 6.6 | 761 | 63.4 ± 10.9 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scheff, D.S.; Campbell, J.F.; Arthur, F.H. Seasonal, Landscape, and Attractant Effects on Lesser Grain Borer, Rhyzopertha dominica (F.), Captures in Northeast Kansas. Agronomy 2022, 12, 99. https://doi.org/10.3390/agronomy12010099
Scheff DS, Campbell JF, Arthur FH. Seasonal, Landscape, and Attractant Effects on Lesser Grain Borer, Rhyzopertha dominica (F.), Captures in Northeast Kansas. Agronomy. 2022; 12(1):99. https://doi.org/10.3390/agronomy12010099
Chicago/Turabian StyleScheff, Deanna S., James F. Campbell, and Franklin H. Arthur. 2022. "Seasonal, Landscape, and Attractant Effects on Lesser Grain Borer, Rhyzopertha dominica (F.), Captures in Northeast Kansas" Agronomy 12, no. 1: 99. https://doi.org/10.3390/agronomy12010099