
Towards Better Grafting: SCoT and CDDP Analyses for Prediction of the Tomato Rootstocks Performance under Drought Stress

 ,
,  ,
,  ,
,  , , ,
, , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Grafting
2.2. Molecular Markers Analysis
2.2.1. Purification of Tomato DNA
2.2.2. CDDP-PCR Analysis
2.2.3. SCoT-PCR Analysis
2.2.4. Molecular Data Analysis
2.3. Greenhouse Experiment
2.3.1. Growth Conditions
2.3.2. Water Stress Treatments
2.3.3. Plant Growth and Yield
2.3.4. Fruit Quality
2.3.5. Leaf Elements Content
2.3.6. Leaf Proline, Gibberellic Acid and Abscisic Acid Content
2.3.7. Leaf Antioxidant Enzymes Content
2.4. Clustering Analysis
2.5. Statistical Analysis
3. Results
3.1. Analysis of CDDP and SCoT
3.2. Molecular Phylogeny Analysis
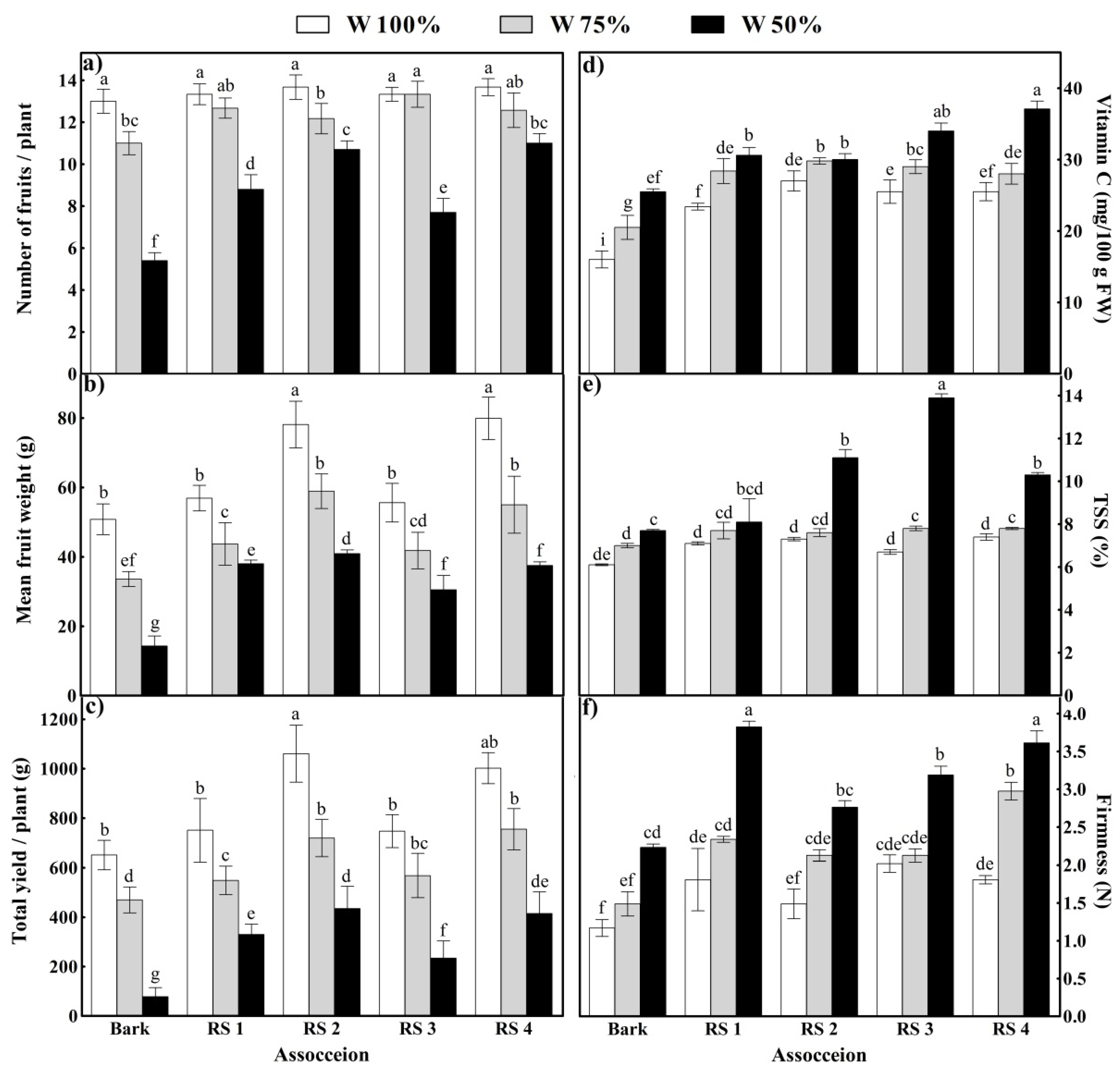
3.3. Plant Growth
3.4. Yield and Quality of Tomato Fruits
3.5. Mineral Content in Tomato Shoots
3.6. Plant Hormones and Antioxidant Enzymes in Tomato Shoots
3.7. Clustering Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ballon-Landa, E.; Parsons, J.K. Nutrition, Physical Activity, and Lifestyle Factors in Prostate Cancer Prevention. Curr. Opin. Urol. 2018, 28, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, D.; Bhardwaj, S.; Landi, M.; Sharma, A.; Ramakrishnan, M.; Sharma, A. The Impact of Drought in Plant Metabolism: How to Exploit Tolerance Mechanisms to Increase Crop Production. Appl. Sci. 2020, 10, 5692. [Google Scholar] [CrossRef]
- Abbasi, T.; Abbasi, S. Biomass Energy and the Environmental Impacts Associated with Its Production and Utilization. Renew. Sustain. Energy Rev. 2010, 14, 919–937. [Google Scholar] [CrossRef]
- Brodersen, C.R.; Roddy, A.B.; Wason, J.W.; McElrone, A.J. Functional Status of Xylem through Time. Annu. Rev. Plant Biol. 2019, 70, 407–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Dindaroglu, T.; Abdul-Wajid, H.H.; Battaglia, M.L. Drought Stress Impacts on Plants and Different Approaches to Alleviate Its Adverse Effects. Plants 2021, 10, 259. [Google Scholar] [CrossRef]
- Food and Agriculture Organization. Future of Food and Agriculture 2018: Alternative Pathways to 2050; Food and Agriculture Organization: Rome, Italy, 2018. [Google Scholar]
- Kumawat, K.R.; Sharma, N. Effect of Drought Stress on Plants Growth. Pop. Kheti 2018, 6, 239–241. [Google Scholar]
- Sharma, A.; Kumar, V.; Shahzad, B.; Ramakrishnan, M.; Singh Sidhu, G.P.; Bali, A.S.; Handa, N.; Kapoor, D.; Yadav, P.; Khanna, K.; et al. Photosynthetic Response of Plants under Different Abiotic Stresses: A Review. J. Plant Growth Regul. 2020, 39, 509–531. [Google Scholar] [CrossRef]
- Ibrahim, M.F.; Elbar, O.H.A.; Farag, R.; Hikal, M.; El-Kelish, A.; El-Yazied, A.A.; Alkahtani, J.; El-Gawad, H.G.A. Melatonin Counteracts Drought Induced Oxidative Damage and Stimulates Growth, Productivity and Fruit Quality Properties of Tomato Plants. Plants 2020, 9, 1276. [Google Scholar] [CrossRef] [PubMed]
- Fathi, A.; Tari, D.B. Effect of Drought Stress and Its Mechanism in Plants. Int. J. Life Sci. 2016, 10, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, M.F.; El-Samad, A.; Ashour, H.; El-Sawy, A.M.; Hikal, M.; Elkelish, A.; El-Gawad, H.A.; El-Yazied, A.A.; Hozzein, W.N.; Farag, R.; et al. Regulation of Agronomic Traits, Nutrient Uptake, Osmolytes and Antioxidants of Maize as Influenced by Exogenous Potassium Silicate under Deficit Irrigation and Semiarid Conditions. Agronomy 2020, 10, 1212. [Google Scholar] [CrossRef]
- Salehi-Lisar, S.Y.; Bakhshayeshan-Agdam, H. Drought stress in plants: Causes, consequences, and tolerance. In Drought Stress Tolerance in Plants; Springer: Berlin/Heidelberg, Germany, 2016; Volume 1, pp. 1–16. [Google Scholar]
- Singh, H.; Kumar, P.; Chaudhari, S.; Edelstein, M. Tomato Grafting: A Global Perspective. HortScience 2017, 52, 1328–1336. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Liu, Y.; Cao, B.; Chen, Z.; Xu, K. The Effectiveness of Grafting to Improve Drought Tolerance in Tomato. Plant Growth Regul. 2020, 91, 157–167. [Google Scholar] [CrossRef]
- Zhang, Z.; Cao, B.; Gao, S.; Xu, K. Grafting Improves Tomato Drought Tolerance through Enhancing Photosynthetic Capacity and Reducing ROS Accumulation. Protoplasma 2019, 256, 1013–1024. [Google Scholar] [CrossRef]
- Aguilera, J.G.; Pessoni, L.A.; Rodrigues, G.B.; Elsayed, A.Y.; Da Silva, D.J.; De Barros, E.G. Genetic Variability by ISSR Markers in Tomato (Solanum Lycopersicon Mill.). Rev. Bras. Ciências Agrárias 2011, 6, 243–252. [Google Scholar] [CrossRef] [Green Version]
- Robinson, J.P.; Harris, S.A. Amplified Fragment Length Polymorphism and Microsatellites: A phylogenetic perspective. In Proceedings of the in Which DNA Marker for Which Purpose? Final Compendium of the Research Project Development, Optimisation and Validation of Stephen P. DiFazio; Citeseer: State College, PA, USA, 1999. [Google Scholar]
- Osuna-Martínez, U.; Reyes-Esparza, J.; Rodríguez-Fragoso, L. Cactus (Opuntia Ficus-Indica): A Review on Its Antioxidants Properties and Potential Pharmacological Use in Chronic Diseases. Nat. Prod. Chem. Res. 2014, 2, 153. [Google Scholar]
- Atia, M.A.; Abdeldaym, E.A.; Abdelsattar, M.; Ibrahim, D.S.; Saleh, I.; Abd Elwahab, M.; Osman, G.H.; Arif, I.A.; Abdelaziz, M.E. Piriformospora Indica Promotes Cucumber Tolerance against Root-Knot Nematode by Modulating Photosynthesis and Innate Responsive Genes. Saudi J. Biol. Sci. 2020, 27, 279–287. [Google Scholar] [CrossRef]
- Collard, B.; Mackill, D. Conserved DNA-Derived Polymorphism (CDDP): A Simple and Novel Method for Generating DNA Markers in Plants. Plant Mol. Biol. Rep. 2009, 27, 558–562. [Google Scholar] [CrossRef]
- Atia, M.A.; Sakr, M.M.; Mokhtar, M.M.; Adawy, S.S. Development of sex-specific PCR-based markers in date palm. In Date Palm Biotechnology Protocols Volume II; Springer: Berlin/Heidelberg, Germany, 2017; pp. 227–244. [Google Scholar]
- Letunic, I.; Bork, P. Interactive Tree Of Life v2: Online Annotation and Display of Phylogenetic Trees Made Easy. Nucleic Acids Res. 2011, 39, W475–W478. [Google Scholar] [CrossRef]
- Abouseadaa, H.H.; Atia, M.A.; Younis, I.Y.; Issa, M.Y.; Ashour, H.A.; Saleh, I.; Osman, G.H.; Arif, I.A.; Mohsen, E. Gene-Targeted Molecular Phylogeny, Phytochemical Profiling, and Antioxidant Activity of Nine Species Belonging to Family Cactaceae. Saudi J. Biol. Sci. 2020, 27, 1649–1658. [Google Scholar] [CrossRef]
- Abdelgawad, K.F.; El-Mogy, M.M.; Mohamed, M.I.A.; Garchery, C.; Stevens, R.G. Increasing Ascorbic Acid Content and Salinity Tolerance of Cherry Tomato Plants by Suppressed Expression of the Ascorbate Oxidase Gene. Agronomy 2019, 9, 51. [Google Scholar] [CrossRef] [Green Version]
- AOAC International. Official Methods of Analysis of the Association of Official Analytical Chemists; AOAC International: Rockville, MD, USA, 1925; Volume 2. [Google Scholar]
- Piper, C.S. Soil and Plant Analysis; Scientific Publishers: Jodhpur, Rajasthan, 2017. [Google Scholar]
- Bates, L.S.; Waldren, R.P.; Teare, I. Rapid Determination of Free Proline for Water-Stress Studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Fales, H.; Jaouni, T.; Babashak, J. Simple Device for Preparing Ethereal Diazomethane without Resorting to Codistillation. Anal. Chem. 1973, 45, 2302–2303. [Google Scholar] [CrossRef]
- Abdelaziz, M.; Abdeldaym, E. Effect of Grafting and Different Ec Levels of Saline Irrigation Water on Growth, Yield and Fruit Quality of Tomato (Lycopersicon Esculentum) in Greenhouse. Plant Arch. 2019, 19, 3021–3027. [Google Scholar]
- Abdelaziz, M.E.; Abdelsattar, M.; Abdeldaym, E.A.; Atia, M.A.; Mahmoud, A.W.M.; Saad, M.M.; Hirt, H. Piriformospora Indica Alters Na+/K+ Homeostasis, Antioxidant Enzymes and LeNHX1 Expression of Greenhouse Tomato Grown under Salt Stress. Sci. Hortic. 2019, 256, 108532. [Google Scholar] [CrossRef]
- Wang, Q.; Zhao, R.; Chen, Q.; da Silva, J.A.T.; Chen, L.; Yu, X. Physiological and Biochemical Responses of Two Herbaceous Peony Cultivars to Drought Stress. HortScience 2019, 54, 492–498. [Google Scholar] [CrossRef] [Green Version]
- García-Limones, C.; Hervás, A.; Navas-Cortés, J.A.; Jiménez-Díaz, R.M.; Tena, M. Induction of an Antioxidant Enzyme System and Other Oxidative Stress Markers Associated with Compatible and Incompatible Interactions between Chickpea (Cicer Arietinum L.) and Fusarium Oxysporum f. sp. Ciceris. Physiol. Mol. Plant Pathol. 2002, 61, 325–337. [Google Scholar] [CrossRef]
- Baier, M.; Noctor, G.; Foyer, C.H.; Dietz, K.-J. Antisense Suppression of 2-Cysteine Peroxiredoxin in Arabidopsis Specifically Enhances the Activities and Expression of Enzymes Associated with Ascorbate Metabolism but Not Glutathione Metabolism. Plant Physiol. 2000, 124, 823–832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metsalu, T.; Vilo, J. ClustVis: A Web Tool for Visualizing Clustering of Multivariate Data Using Principal Component Analysis and Heatmap. Nucleic Acids Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef]
- Shapiro, S.S.; Wilk, M.B. An Analysis of Variance Test for Normality (Complete Samples). Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
- Hartley, H.O. The Maximum F-Ratio as a Short-Cut Test for Heterogeneity of Variance. Biometrika 1950, 37, 308–312. [Google Scholar] [CrossRef]
- Abdeldym, E.A.; El-Mogy, M.M.; Abdellateaf, H.R.; Atia, M.A. Genetic Characterization, Agro-Morphological and Physiological Evaluation of Grafted Tomato under Salinity Stress Conditions. Agronomy 2020, 10, 1948. [Google Scholar] [CrossRef]
- Cardoso, A.A.; Gori, A.; Da-Silva, C.J.; Brunetti, C. Abscisic Acid Biosynthesis and Signaling in Plants: Key Targets to Improve Water Use Efficiency and Drought Tolerance. Appl. Sci. 2020, 10, 6322. [Google Scholar] [CrossRef]
- Ali, S.; Hayat, K.; Iqbal, A.; Xie, L. Implications of Abscisic Acid in the Drought Stress Tolerance of Plants. Agronomy 2020, 10, 1323. [Google Scholar] [CrossRef]
- Yamaguchi, S. Gibberellin Metabolism and Its Regulation. Annu. Rev. Plant Biol. 2008, 59, 225–251. [Google Scholar] [CrossRef]
- Litvin, A.G.; van Iersel, M.W.; Malladi, A. Drought Stress Reduces Stem Elongation and Alters Gibberellin-Related Gene Expression during Vegetative Growth of Tomato. J. Am. Soc. Hortic. Sci. 2016, 141, 591–597. [Google Scholar] [CrossRef]
- Li, J.; Sima, W.; Ouyang, B.; Wang, T.; Ziaf, K.; Luo, Z.; Liu, L.; Li, H.; Chen, M.; Huang, Y.; et al. Tomato SlDREB Gene Restricts Leaf Expansion and Internode Elongation by Downregulating Key Genes for Gibberellin Biosynthesis. J. Exp. Bot. 2012, 63, 6407–6420. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Cao, B.; Li, N.; Chen, Z.; Xu, K. Comparative Transcriptome Analysis of the Regulation of ABA Signaling Genes in Different Rootstock Grafted Tomato Seedlings under Drought Stress. Environ. Exp. Bot. 2019, 166, 103814. [Google Scholar] [CrossRef]
- Shohat, H.; Illouz-Eliaz, N.; Kanno, Y.; Seo, M.; Weiss, D. The Tomato DELLA Protein PROCERA Promotes Abscisic Acid Responses in Guard Cells by Upregulating an Abscisic Acid Transporter. Plant Physiol. 2020, 184, 518–528. [Google Scholar] [CrossRef] [PubMed]
- Ünyayar, S.; Aktoklu, E.; Büyükasik, Y. The Response to Exogenous Abscisic Acid of the Roots of Notabilis and Its Wild-Type Tomato under Drought Stress. Isr. J. Plant Sci. 2004, 52, 294–299. [Google Scholar] [CrossRef]
- Postiglione, A.E.; Muday, G.K. The Role of ROS Homeostasis in ABA-Induced Guard Cell Signaling. Front. Plant Sci. 2020, 11, 968. [Google Scholar] [CrossRef]
- Shivakrishna, P.; Reddy, K.A.; Rao, D.M. Effect of PEG-6000 Imposed Drought Stress on RNA Content, Relative Water Content (RWC), and Chlorophyll Content in Peanut Leaves and Roots. Saudi J. Biol. Sci. 2018, 25, 285–289. [Google Scholar]
- Liang, G.; Liu, J.; Zhang, J.; Guo, J. Effects of Drought Stress on Photosynthetic and Physiological Parameters of Tomato. J. Am. Soc. Hortic. Sci. 2020, 145, 12–17. [Google Scholar] [CrossRef] [Green Version]
- Medyouni, I.; Zouaoui, R.; Rubio, E.; Serino, S.; Ahmed, H.B.; Bertin, N. Effects of Water Deficit on Leaves and Fruit Quality during the Development Period in Tomato Plant. Food Sci. Nutr. 2021, 9, 1949–1960. [Google Scholar] [CrossRef]
- Dalal, V.K.; Tripathy, B.C. Water-Stress Induced Downsizing of Light-Harvesting Antenna Complex Protects Developing Rice Seedlings from Photo-Oxidative Damage. Sci. Rep. 2018, 8, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Abdallah, M.B.; Trupiano, D.; Polzella, A.; De Zio, E.; Sassi, M.; Scaloni, A.; Zarrouk, M.; Youssef, N.B.; Scippa, G.S. Unraveling Physiological, Biochemical and Molecular Mechanisms Involved in Olive (Olea Europaea L. Cv. Chétoui) Tolerance to Drought and Salt Stresses. J. Plant Physiol. 2018, 220, 83–95. [Google Scholar] [CrossRef]
- Singh, J.; Thakur, J.K. Photosynthesis and abiotic stress in plants. In Biotic and Abiotic Stress Tolerance in Plants; Springer: Berlin/Heidelberg, Germany, 2018; pp. 27–46. [Google Scholar]
- Cui, J.; Shao, G.; Lu, J.; Keabetswe, L.; Hoogenboom, G. Yield, Quality and Drought Sensitivity of Tomato to Water Deficit during Different Growth Stages. Sci. Agrícola 2019, 77, e20180390. [Google Scholar] [CrossRef] [Green Version]
- Falchi, R.; Bonghi, C.; Drincovich, M.F.; Famiani, F.; Lara, M.V.; Walker, R.P.; Vizzotto, G. Sugar Metabolism in Stone Fruit: Source-Sink Relationships and Environmental and Agronomical Effects. Front. Plant Sci. 2020, 11, 1820. [Google Scholar] [CrossRef] [PubMed]
- Singh, G.; Kachwaya, D.S.; Kuchi, V.S.; Singh, M.; Singh, J.; Singh, B. Influence of Foliar Application of Mineral Nutrients on Vegetative and Fruit Quality of Strawberry (Fragaria × ananassa Duch.) Cv. Chandler. J. Plant Sci. Res. 2020, 36, 323–328. [Google Scholar] [CrossRef]
- Kıran, S.; Kuşvuran, Ş.; Talhouni, M.; Sonmez, K.; Ellialtıoglu, Ş.; Özkay, F. Studies on Some Biochemical Changes and Ion Regulation in Some Tomato Genotypes Exposed to Drought Stress. In Proceedings of the VI Balkan Symposium on Vegetables and Potatoes. Acta Hortic. 2014, 1142, 369–376. [Google Scholar]
- Rouphael, Y.; Cardarelli, M.; Schwarz, D.; Franken, P.; Colla, G. Effects of drought on nutrient uptake and assimilation in vegetable crops. In Plant Responses to Drought Stress; Springer: Berlin/Heidelberg, Germany, 2012; pp. 171–195. [Google Scholar]
- Bista, D.R.; Heckathorn, S.A.; Jayawardena, D.M.; Mishra, S.; Boldt, J.K. Effects of Drought on Nutrient Uptake and the Levels of Nutrient-Uptake Proteins in Roots of Drought-Sensitive and-Tolerant Grasses. Plants 2018, 7, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dariva, F.D.; Pessoa, H.P.; Copati, M.G.F.; de Almeida, G.Q.; de Castro Filho, M.N.; de Toledo Picoli, E.A.; da Cunha, F.F.; Nick, C. Yield and Fruit Quality Attributes of Selected Tomato Introgression Lines Subjected to Long-Term Deficit Irrigation. Sci. Hortic. 2021, 289, 110426. [Google Scholar] [CrossRef]
- Sivakumar, R.; Srividhya, S. Impact of Drought on Flowering, Yield and Quality Parameters in Diverse Genotypes of Tomato (Solanum Lycopersicum L.). Adv. Hortic. Sci. 2016, 30, 3–11. [Google Scholar]
- Živanović, B.; Milić Komić, S.; Tosti, T.; Vidović, M.; Prokić, L.; Veljović Jovanović, S. Leaf Soluble Sugars and Free Amino Acids as Important Components of Abscisic Acid—Mediated Drought Response in Tomato. Plants 2020, 9, 1147. [Google Scholar] [CrossRef]
- Petrović, I.; Jovanović, Z.; Stikić, R.; Marjanović, M.; Savić, S. Influence of Severe Drought on Leaf Response in ABA Contrasting Tomato Genotypes (Wild Type and Flacca Mutant). Biol. Life Sci. Forum 2020, 41, 8867. [Google Scholar] [CrossRef]
- Ijaz, R.; Ejaz, J.; Gao, S.; Liu, T.; Imtiaz, M.; Ye, Z.; Wang, T. Overexpression of Annexin Gene AnnSp2, Enhances Drought and Salt Tolerance through Modulation of ABA Synthesis and Scavenging ROS in Tomato. Sci. Rep. 2017, 7, 1–14. [Google Scholar]
- Barupal, T.; Meena, M.; Sharma, K. Inhibitory Effects of Leaf Extract of Lawsonia Inermis on Curvularia Lunata and Characterization of Novel Inhibitory Compounds by GC–MS Analysis. Biotechnol. Rep. 2019, 23, e00335. [Google Scholar] [CrossRef]
- Verslues, P.E.; Sharma, S. Proline Metabolism and Its Implications for Plant-Environment Interaction. Arab. Book 2010, 8, e0140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, S.; Fartyal, D.; Agarwal, A.; Shukla, T.; James, D.; Kaul, T.; Negi, Y.K.; Arora, S.; Reddy, M.K. Abiotic Stress Tolerance in Plants: Myriad Roles of Ascorbate Peroxidase. Front. Plant Sci. 2017, 8, 581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mafakheri, A.; Siosemardeh, A.; Bahramnejad, B.; Struik, P.; Sohrabi, Y. Effect of Drought Stress and Subsequent Recovery on Protein, Carbohydrate Contents, Catalase and Peroxidase Activities in Three Chickpea (‘Cicer arietinum’) Cultivars. Aust. J. Crop Sci. 2011, 5, 1255–1260. [Google Scholar]
- Çelik, Ö.; Ayan, A.; Atak, Ç. Enzymatic and Non-Enzymatic Comparison of Two Different Industrial Tomato (Solanum Lycopersicum) Varieties against Drought Stress. Bot. Stud. 2017, 58, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Zhou, R.; Kong, L.; Yu, X.; Ottosen, C.-O.; Zhao, T.; Jiang, F.; Wu, Z. Oxidative Damage and Antioxidant Mechanism in Tomatoes Responding to Drought and Heat Stress. Acta Physiol. Plant. 2019, 41, 20. [Google Scholar] [CrossRef]
- Chang, L.; Sun, H.; Yang, H.; Wang, X.; Su, Z.; Chen, F.; Wei, W. Over-Expression of Dehydroascorbate Reductase Enhances Oxidative Stress Tolerance in Tobacco. Electron. J. Biotechnol. 2017, 25, 1–8. [Google Scholar] [CrossRef]
- Murshed, R.; Lopez-Lauri, F.; Sallanon, H. Effect of Water Stress on Antioxidant Systems and Oxidative Parameters in Fruits of Tomato (Solanum Lycopersicon L., Cv. Micro-Tom). Physiol. Mol. Biol. Plants 2013, 19, 363–378. [Google Scholar] [CrossRef] [Green Version]
- Raja, V.; Qadir, S.U.; Alyemeni, M.N.; Ahmad, P. Impact of Drought and Heat Stress Individually and in Combination on Physio-Biochemical Parameters, Antioxidant Responses, and Gene Expression in Solanum Lycopersicum. 3 Biotech 2020, 10, 1–18. [Google Scholar] [CrossRef]
- Yao, X.; Yang, R.; Zhao, F.; Wang, S.; Li, C.; Zhao, W. An Analysis of Physiological Index of Differences in Drought Tolerance of Tomato Rootstock Seedlings. J. Plant Biol. 2016, 59, 311–321. [Google Scholar] [CrossRef]
- Sánchez-Rodríguez, E.; Leyva, R.; Constán-Aguilar, C.; Romero, L.; Ruiz, J. Grafting under Water Stress in Tomato Cherry: Improving the Fruit Yield and Quality. Ann. Appl. Biol. 2012, 161, 302–312. [Google Scholar] [CrossRef]
- Abdelaziz, M.E.; Atia, M.A.; Abdelsattar, M.; Abdelaziz, S.M.; Ibrahim, T.A.; Abdeldaym, E.A. Unravelling the Role of Piriformospora indica in Combating Water Deficiency by Modulating Physiological Performance and Chlorophyll Metabolism-Related Genes in Cucumis sativus. Horticulturae 2021, 7, 399. [Google Scholar] [CrossRef]
- Mokhtar, M.M.; El Allali, A.; Hegazy, M.E.F.; Atia, M.A.M. PlantPathMarks (PPMdb): An interactive hub for pathways-based markers in plant genomes. Sci. Rep. 2020, 11, 1–12. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Code | Primer Sequence | Gene | Gene Function | Amino Acid Motif | Length | % GC |
|---|---|---|---|---|---|---|
| CDDP-1 | TGGCGSAAGTACGGCCAG | WRKY | Transcription factor for developmental and physiological roles | WRKYGQ | 18 | 67 |
| CDDP-2 | GTGGTTGTGCTTGCC | GKHNH | 15 | 60 | ||
| CDDP-3 | GCCCTCGTASGTSGT | TTYEG | 15 | 67 | ||
| CDDP-4 | GGCAAGGGCTGCCGC | MYB | Implicated in secondary metabolism, abiotic and biotic stresses, cellular morphogenesis | GKSCR | 15 | 80 |
| CDDP-5 | GGCAAGGGCTGCCGG | GKSCR | 15 | 80 | ||
| CDDP-6 | AAGGGSAAGCTSCCSAAG | KNOX | Homeobox genes that function as transcription factors with a unique homeodomain | KGKLPK | 18 | 61 |
| CDDP-7 | CACTGGTGGGAGCTSCAC | HWWELH | 18 | 67 | ||
| CDDP-8 | AAGCGSCACTGGAAGCC | KRHWKP | 17 | 65 | ||
| CDDP-9 | ATGGGCCGSGGCAAGGTGC | MADS | Involved in controlling floral organ initiation and development | MGRGKV | 19 | 74 |
| CDDP-10 | CTSTGCGACCGSGAGGTC | LCDAEV | 18 | 72 | ||
| CDDP-11 | ACSCCSATCCACCGC | ABP1 | Auxin-binding protein | TPIHR | 15 | 73 |
| Primer Code | Primer Sequence | % GC |
|---|---|---|
| SCoT-1 | CAACAATGGCTACCACCA | 50 |
| SCoT-2 | CACCATGGCTACCACCAG | 61 |
| SCoT-3 | CAACAATGGCTACCACCG | 56 |
| SCoT-4 | CAACAATGGCTACCACCT | 50 |
| SCoT-5 | CAACAATGGCTACCACGA | 50 |
| SCoT-6 | CAACAATGGCTACCACGC | 56 |
| SCoT-7 | ACCATGGCTACCACCGGG | 67 |
| SCoT-8 | CAACAATGGCTACCACGT | 50 |
| SCoT-9 | CAACAATGGCTACCAGCA | 50 |
| SCoT-10 | CAACAATGGCTACCAGCC | 56 |
| SCoT-11 | AAGCAATGGCTACCACCA | 50 |
| SCoT-12 | ACGACATGGCGACCAACG | 61 |
| SCoT-13 | ACGACATGGCGACCATCG | 61 |
| SCoT-14 | ACGACATGGCGACCACGC | 67 |
| SCoT-15 | ACGACATGGCGACCGCGA | 67 |
| Code | Number of Bands | % of Polymorphism | |
|---|---|---|---|
| Total | Polymorphic | ||
| CDDP | |||
| CDDP-1 | 18 | 4 | 22.2 |
| CDDP-2 | 18 | 6 | 33.3 |
| CDDP-3 | 17 | 5 | 29.4 |
| CDDP-4 | 16 | 6 | 37.5 |
| CDDP-5 | 18 | 5 | 27.8 |
| CDDP-6 | 19 | 7 | 36.8 |
| CDDP-7 | 17 | 6 | 35.3 |
| CDDP-8 | 18 | 4 | 22.2 |
| CDDP-9 | 18 | 5 | 27.8 |
| CDDP-10 | 21 | 4 | 19.0 |
| CDDP-11 | 15 | 3 | 20.0 |
| Total | 195 | 55 | 28.2 |
| Average | 17.7 | 5.0 | |
| SCoT | |||
| SCoT-1 | 21 | 9 | 42.9 |
| SCoT-2 | 15 | 8 | 53.3 |
| SCoT-3 | 15 | 7 | 46.7 |
| SCoT-4 | 16 | 8 | 50.0 |
| SCoT-5 | 19 | 8 | 42.1 |
| SCoT-6 | 21 | 9 | 42.9 |
| SCoT-7 | 13 | 8 | 61.5 |
| SCoT-8 | 17 | 7 | 41.2 |
| SCoT-9 | 13 | 5 | 38.5 |
| SCoT-10 | 16 | 7 | 43.8 |
| SCoT-11 | 18 | 8 | 44.4 |
| SCoT-12 | 17 | 5 | 29.4 |
| SCoT-13 | 16 | 7 | 43.8 |
| SCoT-14 | 20 | 8 | 40.0 |
| SCoT-15 | 19 | 8 | 42.1 |
| Total | 255 | 112 | 43.9 |
| Average | 17.0 | 7.5 | |
| RS1 | RS2 | RS3 | RS4 | Bark | |
|---|---|---|---|---|---|
| RS1 | 100% | ||||
| RS2 | 80% | 100% | |||
| RS3 | 82% | 80% | 100% | ||
| RS4 | 79% | 86% | 79% | 100% | |
| Bark | 87% | 76% | 81% | 76% | 100% |
| RS1 | RS2 | RS3 | RS4 | Bark | |
|---|---|---|---|---|---|
| RS1 | 100% | ||||
| RS2 | 78% | 100% | |||
| RS3 | 80% | 80% | 100% | ||
| RS4 | 75% | 87% | 77% | 100% | |
| Bark | 82% | 78% | 79% | 76% | 100% |
| Mineral | Treatment | Rootstock | ||||
|---|---|---|---|---|---|---|
| Bark | RS1 | RS2 | RS3 | RS4 | ||
| N (%) | W100% | 3.06 b | 3.67 a | 2.99 b | 2.60 cb | 3.10 b |
| W 75% | 2.19 c | 2.52 cb | 2.51 cb | 2.18 c | 2.59 cb | |
| W50% | 1.18 g | 1.83 de | 1.50 f | 1.88 e | 2.01 d | |
| P (%) | W100% | 0.22 ef | 0.37 a | 0.35 ab | 0.30 cb | 0.25 ed |
| W 75% | 0.15 g | 0.25 de | 0.29 cb | 0.25 de | 0.29 cb | |
| W50% | 0.12 h | 0.22 ef | 0.23 ef | 0.23 ef | 0.22 ef | |
| K (%) | W100% | 2.62 ef | 4.25 a | 3.66 b | 3.55 b | 4.14 a |
| W 75% | 2.21 fg | 3.41 bcd | 3.30 cd | 3.03 de | 3.65 b | |
| W50% | 1.88 g | 2.69 de | 2.86 de | 2.40 efg | 3.63 b | |
| Ca (%) | W100% | 0.65 de | 1.29 a | 0.79 bc | 0.76 bc | 0.81 b |
| W 75% | 0.41 g | 0.72 d | 0.66 de | 0.51 fg | 0.67 de | |
| W50% | 0.29 h | 0.49 g | 0.55 f | 0.54 f | 0.60 ef | |
| Mg (%) | W100% | 0.30 gh | 0.50 b | 0.53 a | 0.35 dc | 0.33 def |
| W 75% | 0.22 f | 0.39 cb | 0.36 cde | 0.33 def | 0.34 def | |
| W50% | 0.17 j | 0.21 f | 0.30 ef | 0.22 f | 0.29 ef | |
| Fe (ppm) | W100% | 47.49 fg | 79.40 a | 57.34 cde | 64.71 b | 62.42 bc |
| W 75% | 40.26 h | 52.21 efg | 52.19 efg | 57.27 cde | 53.74 ef | |
| W50% | 38.00 i | 55.34 de | 52.80 efg | 54.65 de | 56.23 de | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Mogy, M.M.; Atia, M.A.M.; Dhawi, F.; Fouad, A.S.; Bendary, E.S.A.; Khojah, E.; Samra, B.N.; Abdelgawad, K.F.; Ibrahim, M.F.M.; Abdeldaym, E.A. Towards Better Grafting: SCoT and CDDP Analyses for Prediction of the Tomato Rootstocks Performance under Drought Stress. Agronomy 2022, 12, 153. https://doi.org/10.3390/agronomy12010153
El-Mogy MM, Atia MAM, Dhawi F, Fouad AS, Bendary ESA, Khojah E, Samra BN, Abdelgawad KF, Ibrahim MFM, Abdeldaym EA. Towards Better Grafting: SCoT and CDDP Analyses for Prediction of the Tomato Rootstocks Performance under Drought Stress. Agronomy. 2022; 12(1):153. https://doi.org/10.3390/agronomy12010153
Chicago/Turabian StyleEl-Mogy, Mohamed M., Mohamed A. M. Atia, Faten Dhawi, Ahmed S. Fouad, Eslam S. A. Bendary, Ebtihal Khojah, Bassem N. Samra, Karima F. Abdelgawad, Mohamed F. M. Ibrahim, and Emad A. Abdeldaym. 2022. "Towards Better Grafting: SCoT and CDDP Analyses for Prediction of the Tomato Rootstocks Performance under Drought Stress" Agronomy 12, no. 1: 153. https://doi.org/10.3390/agronomy12010153