
Absorption and Distribution of Calcium (45Ca) Applied to the Surface of Orange (Citrus sinensis) Fruits at Different Developmental Stages

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site and Plant Material
2.2. 45Ca Measurements
2.3. Tissue Ca Concentrations
2.4. Electron Microscopy Observations
2.5. Statistical Analysis
3. Results
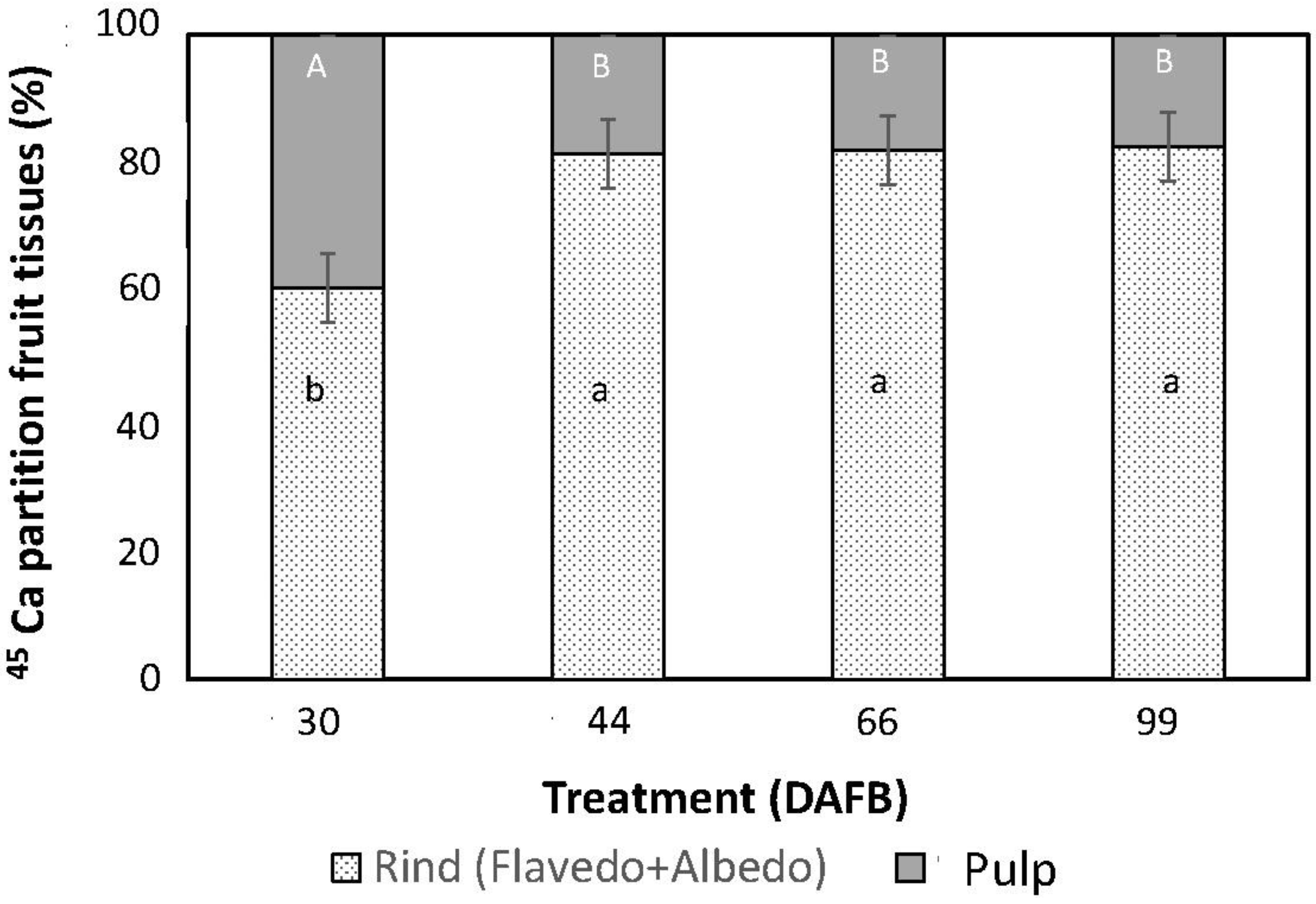
3.1. Absorption and Movement of 45Ca
3.2. Fruit Dry Weight and Ca Concentration
3.3. Fruit Anatomy
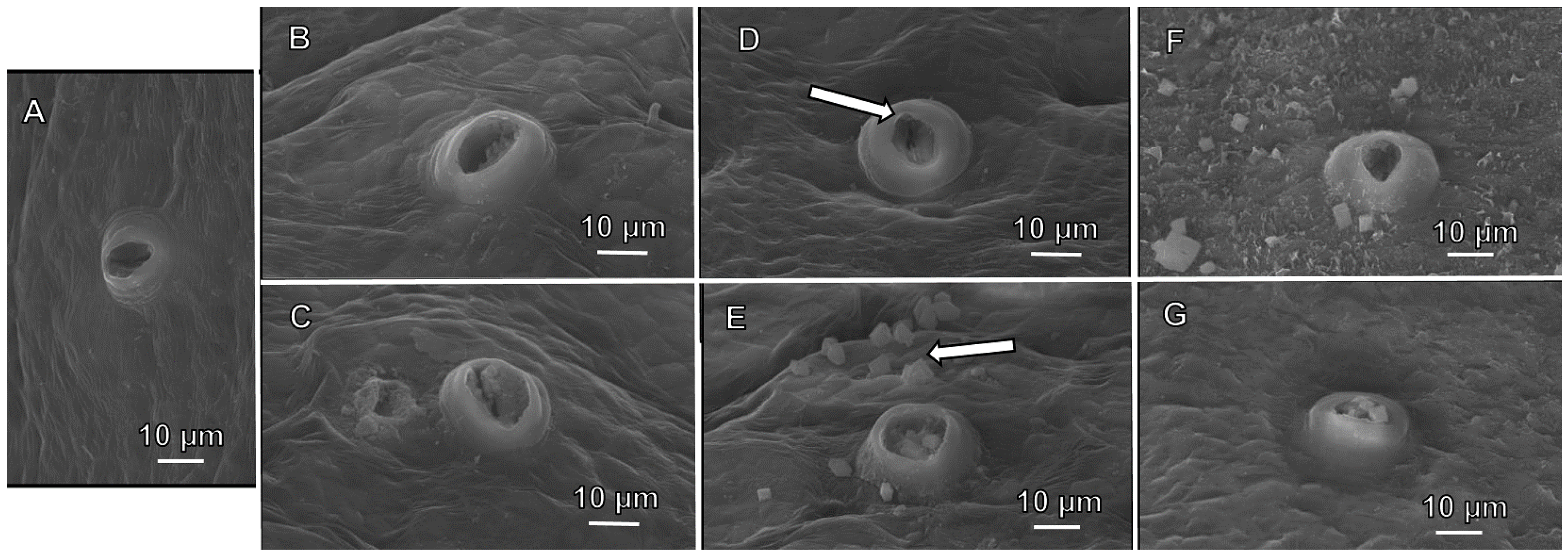
3.3.1. Rind Surface Topography
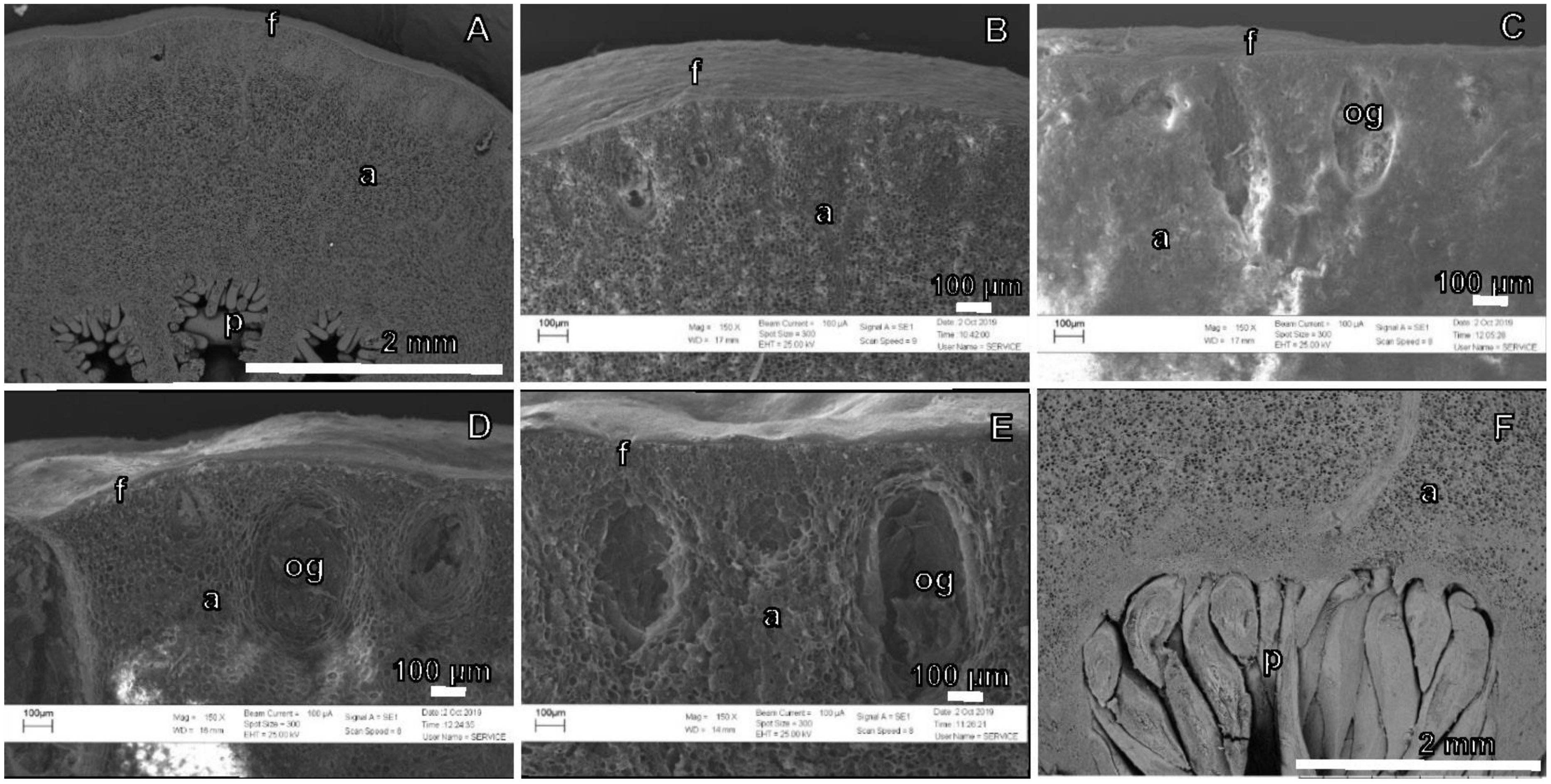
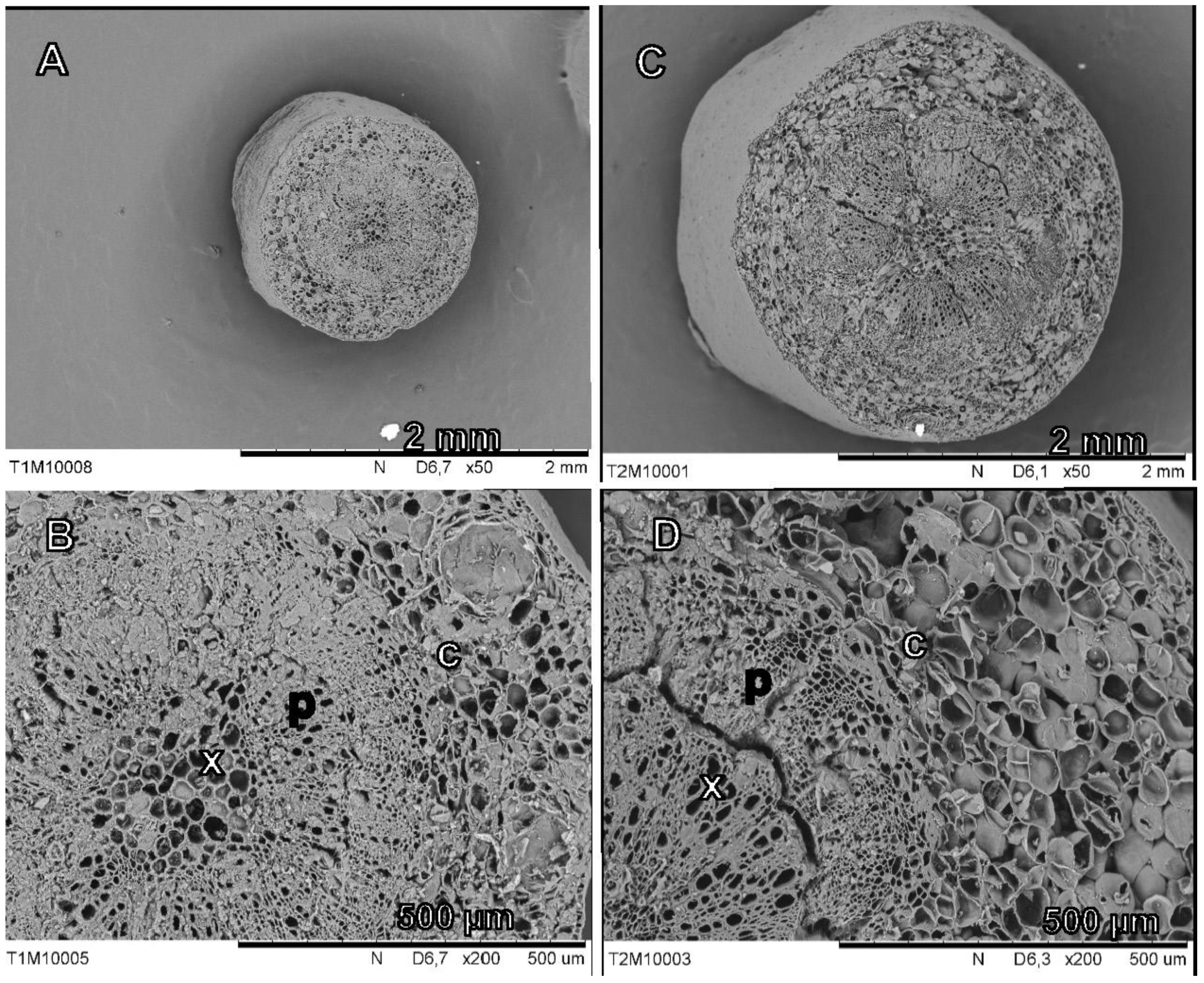
3.3.2. Fruit Cross-Section Anatomy
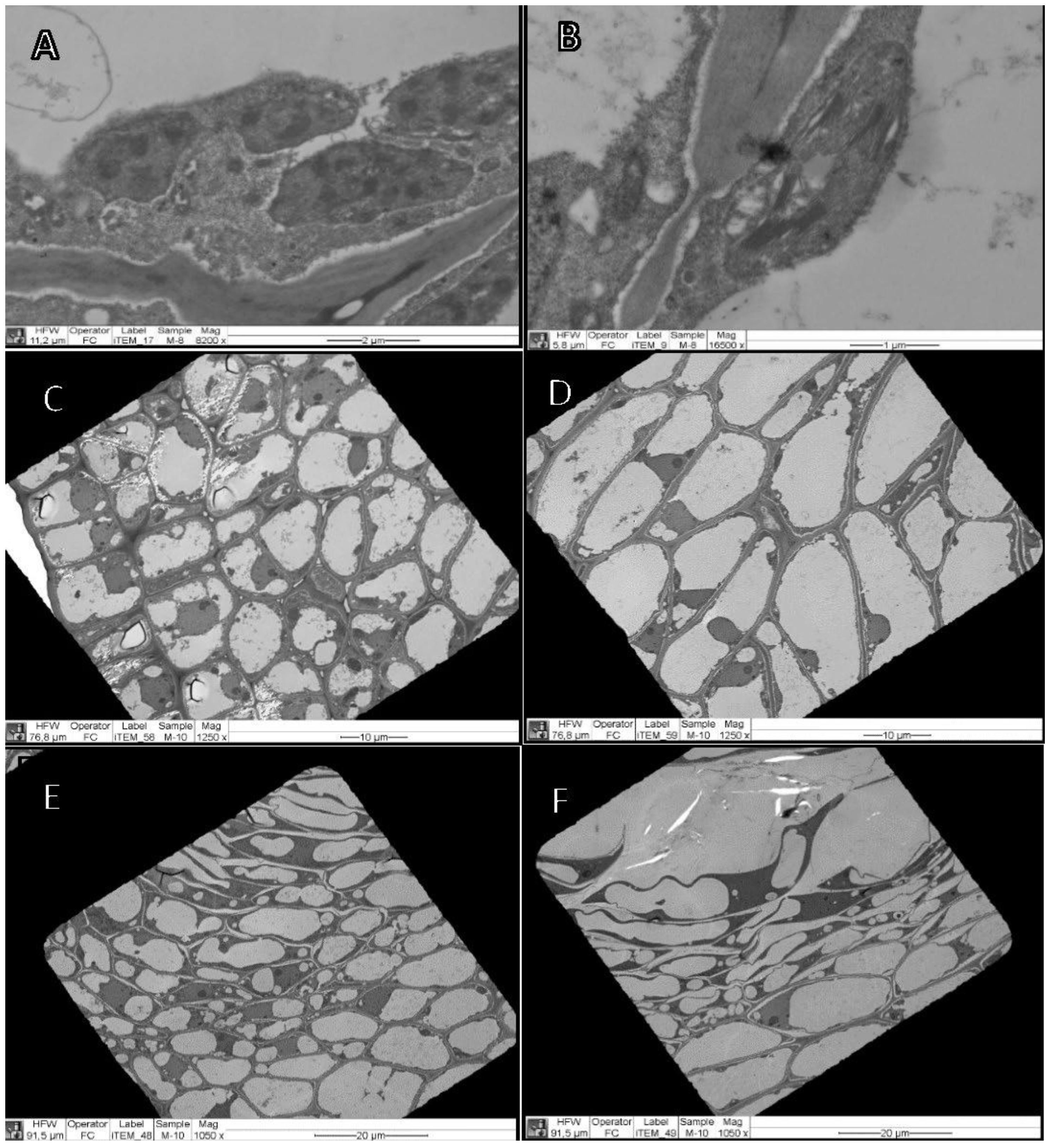
3.3.3. Calcium Oxalate Crystals
3.3.4. Pedicel Cross-Sections
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- White, P.J.; Broadley, M.R. Calcium in plants. Ann. Bot. 2003, 92, 487–511. [Google Scholar] [CrossRef]
- Hocking, B.; Tyerman, S.D.; Burton, R.A.; Gilliham, M. Fruit calcium: Transport and physiology. Front. Plant Sci. 2016, 7, 569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Chen, J. Citrus Fruit-Cracking: Causes and Occurrence. Hortic. Plant J. 2017, 3, 255–260. [Google Scholar] [CrossRef]
- Montanaro, Y.; Dichio, B.; Xiloyannis, C. Significance of fruit transpiration on calcium nutrition in developing apricot fruit. J. Plant Nutr. Soil Sci. 2010, 173, 618–622. [Google Scholar] [CrossRef]
- Montanaro, G.; Dichio, B.; Lang, A.; Mininni, A.N.; Nuzzo, V.; Clearwater, M.J.; Xiloyannis, C. Internal versus external control of calcium nutrition in kiwifruit. J. Plant Nutr. Soil Sci. 2014, 177, 819–830. [Google Scholar] [CrossRef]
- Storey, R.; Treeby, M.T. Nutrient uptake into navel oranges during fruit development. J. Hortic. 2002, 77, 91–99. [Google Scholar] [CrossRef]
- de Freitas, S.T.; Mitcham, E.J. Factors involved in fruit calcium deficiency disorders. Hortic. Rev. (Am. Soc. Hortic. Sci). 2012, 40, 107–146. [Google Scholar] [CrossRef]
- Saure, M.C. Why calcium deficiency is not the cause of blossom-end rot in tomato and pepper fruit-a reappraisal. Sci. Hortic. 2014, 174, 151–154. [Google Scholar] [CrossRef]
- Saure, M.C. Calcium translocation to fleshy fruit: Its mechanism and endogenous control. Sci. Hortic. 2005, 105, 65–89. [Google Scholar] [CrossRef]
- Winkler, A.; Knoche, M. Calcium and the physiology of sweet cherries: A review. Sci. Hortic. 2019, 245, 107–115. [Google Scholar] [CrossRef]
- Treeby, M.T.; Storey, R. Calcium-spray treatments for ameliorating albedo breakdown in navel oranges. Aust. J. Exp. Agric. 2002, 42, 495–502. [Google Scholar] [CrossRef]
- Agustí, M.; Martínez-Fuentes, A.; Mesejo, C. Citrus fruit quality. physiological basis and techniques of improvement. Agrociencia 2002, 6, 1–16. [Google Scholar]
- Bonomelli, C.; Fernández, V.; Martiz, J.; Videla, X.; Arias, M.I.; Rojas-Silva, X.; Nario, A. Absorption and distribution of root, fruit and foliar applied 45Ca in “Clemenules” mandarin trees. J. Sci. Food Agric. 2020, 100, 4643–4650. [Google Scholar] [CrossRef]
- Fernández, V.; Sotiropoulus, T.; Brown, P. Foliar Fertilization: Scientific Principles and Field Practices, 1st ed.; IFA: Paris, France, 2013. [Google Scholar] [CrossRef]
- Li, C.; Wu, J.; Blamey, F.; Wang, L.; Zhou, L.; Paterson, D.J.; Van der Ent, A.; Fernández, V.; Lombi, E.; Wang, Y.; et al. Non-glandular trichomes of sunflower are important in the absorption and translocation of foliar-applied Zn fertilizer. J. Exp. Bot. 2021, 72, 5079–5092. [Google Scholar] [CrossRef] [PubMed]
- Eichert, T.; Kurtz, A.; Steiner, U.; Goldbach, H.E. Size exclusion limits and lateral heterogeneity of the stomatal foliar uptake pathway for aqueous solutes and water-suspended nanoparticles. Physiol. Plant 2008, 134, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Fernández, V.; Gil-Pelegrín, E.; Eichert, T. Foliar water and solute absorption: An update. Plant J. 2021, 105, 870–883. [Google Scholar] [CrossRef]
- Schönherr, J. Calcium chloride penetrates plant cuticles via aqueous pores. Planta 2000, 212, 112–118. [Google Scholar] [CrossRef]
- Wang, J.; Sun, L.; Xie, L.; He, Y.; Luo, T.; Sheng, L.; Luo, Y.; Zeng, Y.; Xu, J.; Deng, X.; et al. Regulation of cuticle formation during fruit development and ripening in “Newhall” navel orange (Citrus sinensis Osbeck) revealed by transcriptomic and metabolomic profiling. Plant Sci. 2016, 243, 131–144. [Google Scholar] [CrossRef]
- Hiratsuka, S.; Suzuki, M.; Nishimura, H.; Nada, K. Fruit photosynthesis in Satsuma mandarin. Plant Sci. 2015, 241, 65–69. [Google Scholar] [CrossRef]
- Storey, R.; Treeby, M.T. The morphology of epicuticular wax and albedo cells of orange fruit in relation to albedo breakdown. J. Hortic. Sci. 1994, 69, 329–338. [Google Scholar] [CrossRef]
- Agustí Fonfría, M. Citricultura, 1st ed.; Mundi-Prensa: Madrid, España, 2000. [Google Scholar]
- Alquezar, B.; Mesejo, C.; Alferez, F.; Agustí, M.; Zacarias, L. Morphological and ultrastructural changes in peel of ‘Navelate’ oranges in relation to variations in relative humidity during postharvest storage and development of peel pitting. Postharvest Biol. Technol. 2010, 56, 163–170. [Google Scholar] [CrossRef]
- Tadeo, F.R.; Moya, J.L.; Iglesias, D.; Talón, M.; Primo-Millo, E. Histología y Citología de Cítricos, 1st ed.; Generalitat Valenciana: Valencia, España, 2003. [Google Scholar]
- Spiegel-Roy, P.; Goldschmidt, E. The Biology of Citrus, 1st ed.; Cambridge University Press: Cambridge, UK, 1996. [Google Scholar]
- Iglesias, D.J.; Cercós, M.; Colmenero-Flores, J.M.; Naranjo, M.A.; Ríos, G.; Carrera, E.; Ruiz-Rivero, O.; Lliso, I.; Morillon, R.; Tadeo, F.R.; et al. Physiology of citrus fruiting. Braz. J. Plant Physiol. 2007, 19, 333–362. [Google Scholar] [CrossRef]
- Bonomelli, C.; Bravo, K.; Vega, A.; Ruiz, R.; Montenegro, G. Use of fura 2 fluorescent dyes as indicator for studying calcium distribution in several plant tissues. Commun. Soil Sci. Plant Anal. 2010, 41, 1061–1072. [Google Scholar] [CrossRef]
- Franceschi, V.R.; Nakata, P.A. Calcium oxalate in plants: Formation and Function. Annu. Rev. Plant Biol. 2005, 56, 41–71. [Google Scholar] [CrossRef]
- Storey, R.; Treeby, M.T. Cryo-SEM study of the early symptoms of peteca in “Lisbon” lemons. J. Hortic. Sci. Biotechnol. 2002, 77, 551–556. [Google Scholar] [CrossRef]
- Undurraga, P.L.; Olaeta, J.A.; Retamales, J.B.; Escobar, J.; Toso, A.M. Effect of maturity and storage temperature on the development of peteca in lemons (Citrus limon (L.) Burm. F.) cv. Eureka. Sci. Hortic. 2009, 122, 56–61. [Google Scholar] [CrossRef]
- Kostman, T.A.; Franceschi, V.R.; Nakata, P.A. Endoplasmic reticulum sub-compartments are involved in calcium sequestration within raphide crystal idioblasts of Pistia stratiotes L. Plant Sci. 2003, 165, 205–212. [Google Scholar] [CrossRef]
- Nakata, P.A. Plant calcium oxalate crystal formation, function, and its impact on human health. Front. Biol. 2012, 7, 254–266. [Google Scholar] [CrossRef]
- Webb, M.A. Cell-Mediated Crystallization of Calcium Oxalate in Plants. Plant Cell 1999, 11, 751. [Google Scholar] [CrossRef] [Green Version]
- Ho, L.C.; Belda, R.; Brown, M.; Andrews, J.; Adams, P. Uptake and transport of calcium and the possible causes of blossom-end rot in tomato. J. Exp. Bot. 1993, 44, 509–518. [Google Scholar] [CrossRef]
- Li, W.; Xu, F.; Chen, S.; Zhang, Z.; Zhao, Y.; Jin, Y.; Li, M.; Zhu, Y.; Liu, Y.; Yang, Y.; et al. A comparative study on Ca content and distribution in two Gesneriaceae species reveals distinctive mechanisms to cope with high rhizospheric soluble calcium. Front. Plant Sci. 2014, 5, 647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lötscher, M.; Hay, M.J.M. Distribution of phosphorus and calcium from nodal roots of Trifolium repens: The relative importance of transport via xylem or phloem. New Phytol. 1996, 133, 445–452. [Google Scholar] [CrossRef]
- Volk, G.M.; Goss, L.J.; Franceschi, V.R. Calcium channels are involved in calcium oxalate crystal formation in specialized cells of Pistia stratiotes L. Ann. Bot. 2004, 93, 741–753. [Google Scholar] [CrossRef] [Green Version]
- Bonomelli, C.; Alcalde, C.; Aguilera, C.; Videla, X.; Rojas-Silva, X.; Nario, A.; Fernández, V. Absorption and mobility of radio-labelled calcium in chili pepper plants and sweet cherry trees. Sci. Agric. 2021, 78, 1–7. [Google Scholar] [CrossRef]
- Kalcsits, L.; van der Heijden, G.; Reid, M.; Mullin, K. Calcium absorption during fruit development in ‘honeycrisp’ apple measured using 44ca as a stable isotope tracer. HortScience 2017, 52, 1804–1809. [Google Scholar] [CrossRef] [Green Version]
- Videla, X.; Rojas-Silva, X.; Nario, A.; Arias, M.I.; Bonomelli, C. Calcination Method of 45 Ca Samples for Isotope Ratio Analysis via Liquid Scintillation. Commun. Soil Sci. Plant Anal. 2019, 50, 412–420. [Google Scholar] [CrossRef]
- Cao, H.; Zhang, J.; Xu, J.; Ye, J.; Yun, Z.; Xu, Q.; Xu, J.; Deng, X. Comprehending crystalline β-carotene accumulation by comparing engineered cell models and the natural carotenoid-rich system of citrus. J. Exp. Bot. 2012, 63, 4403–4417. [Google Scholar] [CrossRef]
- Rueden, C.T.; Schindelin, J.; Hiner, M.C.; DeZonia, B.E.; Walter, A.E.; Arena, E.T.; Eliceiri, K.W. ImageJ2: ImageJ for the next generation of scientific image data. BMC Bioinform. 2017, 18, 1–26. [Google Scholar] [CrossRef]
- Di Renso, J.A.; Casanoves, F.; Balzarini, M.G.; Gonzalez, L.; Tablada, M.; Robledo, C.W. InfoStat Versión 2011; Grupo InfoStat FCA: Córdoba, Argentina, 2011. [Google Scholar]
- Bonomelli, C.; Gil, P.M.; Schaffer, B. Effect of soil type on calcium absorption and partitioning in young avocado (Persea americana Mill.) Trees. Agronomy 2019, 9, 837. [Google Scholar] [CrossRef] [Green Version]
- Madani, B.; Wall, M.; Mirshekari, A.; Bah, A.; Mohamed, M.T.M. Influence of Calcium Foliar Fertilization on Plant Growth, Nutrient Concentrations, and Fruit Quality of Papaya. Am. Soc. Hort. Sci. 2015, 25, 496–504. [Google Scholar] [CrossRef] [Green Version]
- Cronje, P.J.; Barry, G.H.; Huysamer, M. Fruiting position during development of ‘Nules Clementine’ mandarin affects the concentration of K, Mg and Ca in the flavedo. Sci. Hortic. 2011, 130, 829–837. [Google Scholar] [CrossRef]
- Voogt, W.; Blok, C.; Eveleens, B.; Marcelis, L.; Bindraban, P.S. Foliar fertilizer application. In VFRC Report 2013/2; Virtual Fertilizer Research Center: Washington, DC, USA, 2013; Volume 43. [Google Scholar] [CrossRef]
- Knight, T.G.; Klieber, A.; Sedgley, M. The relationship between oil gland and fruit development in Washington Navel orange (Citrus sinensis L. Osbeck). Ann. Bot. 2001, 88, 1039–1047. [Google Scholar] [CrossRef] [Green Version]
- Hetherington, A.M.; Woodward, F.I. The role of stomata in sensing and driving environmental change. Nature 2003, 424, 901–908. [Google Scholar] [CrossRef] [PubMed]
- Hieke, S.; Menzel, C.M.; Ludders, P. Effects of leaf, shoot and fruit development on photosynthesis of lychee trees (Litchi chinensis). Tree Physiol. 2002, 22, 955–961. [Google Scholar] [CrossRef] [Green Version]
- Fernández, V.; Bahamonde, H.A.; Javier Peguero-Pina, J.; Gil-Pelegrín, E.; Sancho-Knapik, D.; Gil, L.; Goldbach, H.E.; Eichert, T. Physico-chemical properties of plant cuticles and their functional and ecological significance. J. Exp. Bot. 2017, 68, 5293–5306. [Google Scholar] [CrossRef]
- Agustí, M.; Almela, V.; Juan, M.; Alferez, F.; Tadeo, F.R.; Zacarías, L. Histological and physiological characterization of rind breakdown of “Navelate” sweet orange. Ann. Bot. 2001, 88, 415–422. [Google Scholar] [CrossRef] [Green Version]
- El-Otmani, M.; Arpaia, M.L.; Coggins, C.W.; Pehrson, J.E.; O’Connell, N.V. Developmental changes in “Valencia” orange fruit epicuticular wax in relation to fruit position on the tree. Sci. Hortic. 1989, 41, 69–81. [Google Scholar] [CrossRef]
- Wang, J.; Hao, H.; Liu, R.; Ma, Q.; Xu, J.; Chen, F.; Cheng, Y.; Deng, X. Comparative analysis of surface wax in mature fruits between Satsuma mandarin (Citrus unshiu) and “Newhall” navel orange (Citrus sinensis) from the perspective of crystal morphology, chemical composition and key gene expression. Food Chem. 2014, 153, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.C.; Zeng, Q.; Ji, Q.X.; Liu, C.F.; Liu, S.B.; Liu, Y. A comparison of the ultrastructure and composition of fruits’ cuticular wax from the wild-type “Newhall” navel orange (Citrus sinensis [L.] Osbeck cv. Newhall) and its glossy mutant. Plant Cell Rep. 2012, 31, 2239–2246. [Google Scholar] [CrossRef]
- Laskowski, L.E.; Monerri, C.; García-Luis, A.; Guardiola, J.L. Vascularización del pedicelo y crecimiento del fruto de Citrus sinensis var. Salustiana y su relación con el contenido de ácido indolacético. Bioagro 2008, 20, 11–20. [Google Scholar]
- Miqueloto, A.; do Amarante, C.V.T.; Steffens, C.A.; dos Santos, A.; Mitcham, E. Relationship between xylem functionality, calcium content an the incidence of bitter pit in apple fruit. Sci. Hortic. 2014, 165, 319–323. [Google Scholar] [CrossRef]
- Song, W.; Yi, J.; Kurniadinata, O.F.; Wang, H.; Huang, X. Linking fruit ca uptake capacity to fruit growth and pedicel anatomy, a cross-species study. Front. Plant Sci. 2018, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Mao, H.; Liu, X.; Qiao, L.; Guo, R. Calcium oxalate crystallization in the presence of amphiphilic phosphoproteins. Cryst. Eng. Comm. 2014, 16, 8841–8851. [Google Scholar] [CrossRef]
- Mathonnet, M.; Dessombz, A.; Bazin, D.; Weil, R.; Frédéric, T.; Pusztaszeri, M.; Daudon, M. Chemical diversity of calcifications in thyroid and hypothetical link to disease. Comptes Rendus Chim. 2016, 19, 1672–1678. [Google Scholar] [CrossRef]
- Saffo, M.B.; Lowenstam, H.A. Calcareous deposits in the renal sac of a molgulid tunicate. Science 1978, 200, 1166–1168. [Google Scholar] [CrossRef] [PubMed]
- Sturm, E.V.; Frank-Kamenetskaya, O.; Vlasov, D.; Zelenskaya, M.; Sazanova, K.; Rusakov, A.; Kniep, R. Crystallization of calcium oxalate hydrates by the interaction of calcite marble with fungus Aspergillus Niger. Am. Mineral. 2015, 100, 2559–2565. [Google Scholar] [CrossRef]
- Storey, R.; Treeby, M.T. Seasonal changes in nutrient concentrations of navel orange fruit. Sci. Hortic. 2000, 84, 67–82. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment DAFB | Date | Development Stage | Fruit Diameter (mm) | Relative Humidity (%) | Air Temperature (°C) |
|---|---|---|---|---|---|
| 30 | 11/14/18 | Fruit Set | 7.6 ± 0.9 | 69.5 | 13.5 |
| 44 | 11/28/18 | Stage I | 15.4 ± 1.9 | 66.5 | 16.0 |
| 66 | 12/20/18 | End Stage I | 29.2 ± 1.7 | 71.2 | 11.6 |
| 99 | 01/22/19 | Mid Stage II | 48.8 ± 1.7 | 64.9 | 17.8 |
| D.W. (%) | Ca Concentration (% D.W.) | ||||
|---|---|---|---|---|---|
| Developmental Stage | DAFB | Fruit | Pedicel | Fruit | Pedicel |
| Fruit Set | 30 | 33.60 ± 1.58 a | 27.44 ± 0.15 a | 0.31 ± 0.05 a | 2.72 ± 0.15 a |
| Stage I | 44 | 32.50 ± 1.36 a | 26.89 ± 0.40 a | 0.48 ± 0.06 b | 2.74 ± 0.10 a |
| End stage I | 66 | 26.47 ± 3.01 b | 32.48 ± 5.91 a | 0.80 ± 0.21 c | 2.49 ± 0.40 a |
| Mid stage II | 99 | 21.23 ± 2.62 b | 49.05 ± 9.33 b | 0.87 ± 0.06 d | 1.73 ± 0.46 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bonomelli, C.; Fernández, V.; Capurro, F.; Palma, C.; Videla, X.; Rojas-Silva, X.; Nario, A.; Mártiz, J. Absorption and Distribution of Calcium (45Ca) Applied to the Surface of Orange (Citrus sinensis) Fruits at Different Developmental Stages. Agronomy 2022, 12, 150. https://doi.org/10.3390/agronomy12010150
Bonomelli C, Fernández V, Capurro F, Palma C, Videla X, Rojas-Silva X, Nario A, Mártiz J. Absorption and Distribution of Calcium (45Ca) Applied to the Surface of Orange (Citrus sinensis) Fruits at Different Developmental Stages. Agronomy. 2022; 12(1):150. https://doi.org/10.3390/agronomy12010150
Chicago/Turabian StyleBonomelli, Claudia, Victoria Fernández, Franco Capurro, Carola Palma, Ximena Videla, Ximena Rojas-Silva, Adriana Nario, and Johanna Mártiz. 2022. "Absorption and Distribution of Calcium (45Ca) Applied to the Surface of Orange (Citrus sinensis) Fruits at Different Developmental Stages" Agronomy 12, no. 1: 150. https://doi.org/10.3390/agronomy12010150