
Effect of Banana Bunchy Top Virus on the Heat Shock Protein Genes of Pentalonia nigronervosa during Temperature Susceptibility and Its Effect on Virus Transmission

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Insect Rearing
2.2. Observation of Developmental Stages
2.3. Thermal Stress on Pentalonia nigronervosa
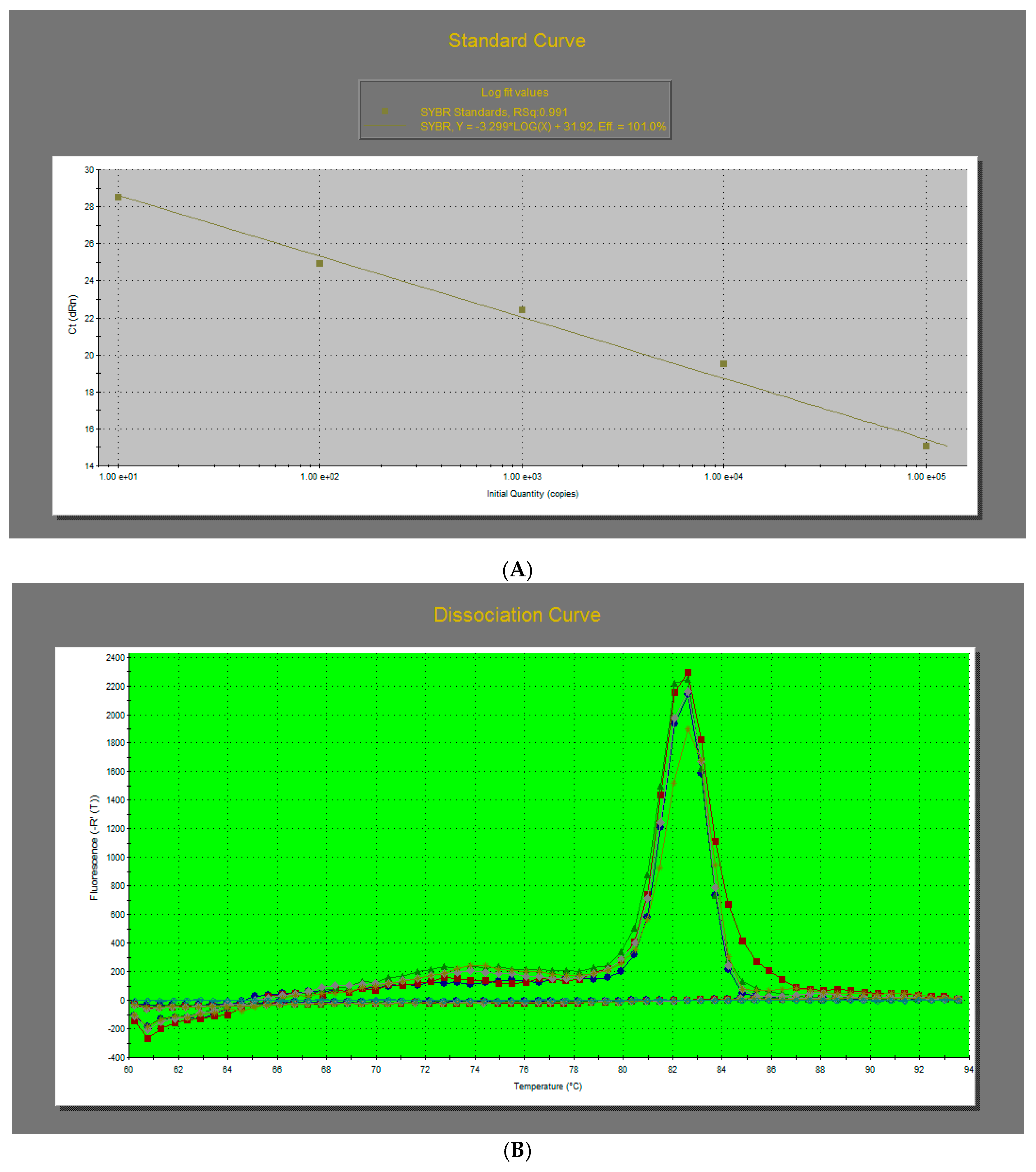
2.4. Quantitative Real-Time PCR (qPCR) Analysis
2.5. Effect of Temperature Stress on Transmission of BBTV
2.6. Preparation of Standard DNA Templates for Standard Curve Construction
2.7. Absolute Quantification of the Viral Titer through qRT-PCR Assay
2.8. Statistical Analysis
3. Results
3.1. Comparison of the Lifespans of Non-Viruliferous and Viruliferous P. nigronervosa
3.2. Effects of Temperature Stress on Adult Mortality
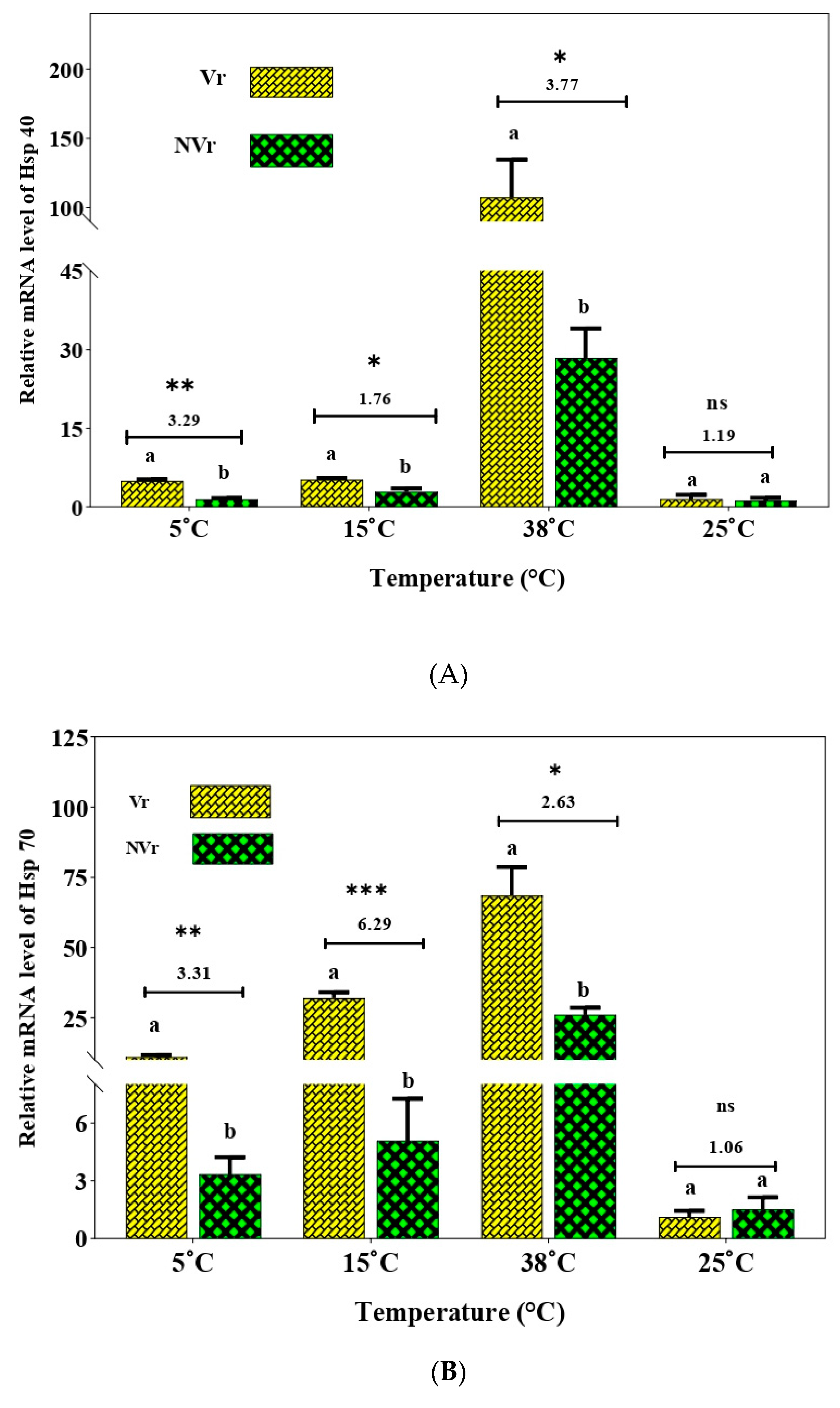
3.3. Effect of Temperature Stress on the Expression of Heat Shock Protein Genes
3.4. Effect of Temperature Stress on Virus Transmission
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stout, M.J.; Thaler, J.S.; Thomma, B.P. Plant-mediated interactions between pathogenic microorganisms and herbivorous arthropods. Annu. Rev. Èntomol. 2006, 51, 663–689. [Google Scholar] [CrossRef]
- Belliure, B.; Janssen, A.; Maris, P.C.; Peters, D.; Sabelis, M.W. Herbivore arthropods benefit from vectoring plant viruses. Ecol. Lett. 2004, 8, 70–79. [Google Scholar] [CrossRef]
- Hatcher, P.E. Three-way interactions between plant pathogenic fungi, herbivorous insects and their host plants. Biol. Rev. 1995, 70, 639–694. [Google Scholar] [CrossRef]
- Hodge, S.; Powell, G. Do plant viruses facilitate their aphid vectors by inducing symptoms that alter behavior and performance? Environ. Entomol. 2008, 37, 1573–1581. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.; Douglas, A.E.; Woodward, S.; Hartley, S.E. Microbial impacts on plant-herbivore interactions: The indirect effects of a birch pathogen on a birch aphid. Oecologia 2003, 134, 388–396. [Google Scholar] [CrossRef] [PubMed]
- Hogenhout, S.A.; Ammar, E.D.; Whitfield, A.E.; Redinbaugh, M.G. Insect vector interactions with persistently trans-mitted viruses. Annu. Rev. Phytopathol. 2008, 46, 327–359. [Google Scholar] [CrossRef] [Green Version]
- Ng, J.C.K.; Perry, K.L. Transmission of plant viruses by aphid vectors. Mol. Plant Pathol. 2004, 5, 505–511. [Google Scholar] [CrossRef]
- Miller, J.W.; Coon, B.F. The Effect of Barley Yellow Dwarf Virus on the Biology of Its Vector the English Grain Aphid, Macrosiphum granarium12. J. Econ. Èntomol. 1964, 57, 970–974. [Google Scholar] [CrossRef]
- Wijkamp, I.; Goldbach, R.; Peters, D. Propagation of tomato spotted wilt virus in Frankliniella occidentalis does neither result in pathological effects nor in transovarial passage of the virus. Èntomol. Exp. Appl. 1996, 81, 285–292. [Google Scholar] [CrossRef]
- Fereres, A.; Kampmeier, G.E.; Irwin, M.E. Aphid attraction and preference for soybean and pepper plants infected with Po-tyviridae. Ann. Entomol. Soc. 1999, 92, 542–548. [Google Scholar] [CrossRef]
- Medina-Ortega, K.J.; Bosque-Perez, N.A.; Ngumbi, E.; Jimenez-Martinez, E.S.; Eigenbrode, S.D. Rhopalosiphum padi (Hemip-tera: Aphididae) responses to volatile cues from barley yellow dwarf-virus infected wheat. Environ. Entomol. 2009, 38, 836–845. [Google Scholar] [CrossRef] [Green Version]
- Stafford, C.A.; Walker, G.P.; Ullman, D.E. Infection with a plant virus modifies vector feeding behavior. Proc. Natl. Acad. Sci. USA 2011, 108, 9350–9355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Froissart, R.; Duomayrou, J.; Vuillaume, F.; Alizon, S.; Michalakis, Y. The virulence-transmission trade-off in vector-borne plant viruses: A review of (non) exisiting studies. Philos. Trans. R. Soc. BBiol. Sci. 2010, 365, 1907–1918. [Google Scholar] [CrossRef] [Green Version]
- Pusag, J.C.A.; Jahan, S.H.; Lee, K.-S.; Lee, S.; Lee, K.-Y. Upregulation of temperature susceptibility in Bemisia tabaci upon acquisition of Tomato yellow leaf curl virus (TYLCV). J. Insect Physiol. 2012, 58, 1343–1348. [Google Scholar] [CrossRef] [PubMed]
- Harrison, B.D. Plant virus ecology: Ingredients, interactions and environmental influences. Ann. Appl. Biol. 1981, 99, 195–209. [Google Scholar] [CrossRef]
- Anhalt, M.D.; Almeida, R. Effect of Temperature, Vector Life Stage, and Plant Access Period on Transmission of Banana bunchy top virus to Banana. Phytopathology 2008, 98, 743–748. [Google Scholar] [CrossRef] [Green Version]
- Fereres, A.; Peñaflor, M.F.G.; Favaro, C.F.; Azevedo, K.E.; Landi, C.H.; Maluta, N.K.; Lopes, J.R. Tomato infection by whitefly-transmitted circulative and non-circulative viruses induces contrasting changes in plant volatiles and vector behav-iour. Viruses 2016, 8, 225. [Google Scholar] [CrossRef] [Green Version]
- Duman, J.G. Antifreeze and Ice Nucleator Proteins in Terrestrial Arthropods. Annu. Rev. Physiol. 2001, 63, 327–357. [Google Scholar] [CrossRef] [PubMed]
- Storey, K.B. Organic Solutes in Freezing Tolerance. Comp. Biochem. Physiol. Part A Physiol. 1997, 117, 319–326. [Google Scholar] [CrossRef]
- Wang, H.-S.; Kang, L. Effect of cooling rates on the cold hardiness and cryoprotectant profiles of locust eggs. Cryobiology 2005, 51, 220–229. [Google Scholar] [CrossRef]
- Zhao, L.; Jones, W.A. Expression of heat shock protein genes in insect stress responses. Invertebr. Surviv. J. 2012, 9, 93–101. [Google Scholar]
- Feder, M.E.; Hofmann, G.E. Heat-shock proteins, molecular chaperones, and the stress response: Evolutionary and Ecological Physiology. Annu. Rev. Physiol. 1999, 61, 243–282. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.K.; Kim, R.; Kim, S.-H. Crystal structure of a small heat-shock protein. Nat. Cell Biol. 1998, 394, 595–599. [Google Scholar] [CrossRef]
- Sørensen, J.G.; Kristensen, T.N.; Loeschcke, V. The evolutionary and ecological role of heat shock proteins. Ecol. Lett. 2003, 6, 1025–1037. [Google Scholar] [CrossRef]
- Santoro, M.G.; Amici, C.; Rossi, A. Role of Heat Shock Proteins in Viral Infection. In Prokaryotic and Eukaryotic Heat Shock Proteins in Infectious Disease. Heat Shock Proteins; Pockley, A., Calderwood, S., Santoro, M., Eds.; Springer: Dordrecht, The Netherlands, 2009; Volume 4, pp. 51–84. [Google Scholar]
- Hooks, C.R.R.; Fukuda, S.; Perez, E.A.; Manandhar, R.; Wang, K.-H.; Wright, M.G.; Almeida, R.P.P. Aphid Transmission of Banana Bunchy Top Virus to Bananas After Treatment with a Bananacide. J. Econ. Èntomol. 2009, 102, 493–499. [Google Scholar] [CrossRef]
- Harding, R.M.; Burns, T.M.; Dale, J.L. Virus-like particles associated with banana bunchy top disease contain small single stranded DNA. J. Gen. Virol. 1991, 72, 225–230. [Google Scholar] [CrossRef]
- Burns, T.M.; Harding, R.M.; Dale, J.L. Evidence that banana bunchy top virus has a multiple component genome. Arch Virol. 1994, 137, 371–380. [Google Scholar] [CrossRef] [PubMed]
- Wanitchakorn, R.; Hafner, G.J.; Harding, R.M.; Dale, J.L. Functional analysis of proteins encoded by banana bunchy top virus DNA-4 to-6. Microbiol. 2000, 81, 299–306. [Google Scholar] [CrossRef]
- Lowe, S.; Browne, M.; Boudjelas, S.; De Poorter, M. 100 of the World’s Worst Invasive Alien Species: A Selection from the Global Invasive Species Database; The Invasive Species Specialist Group (ISSG): Auckland, New Zealand, 2000; Volume 12, p. 12. [Google Scholar]
- Jebakumar, R.M.; Balasubramanian, V.; Selvarajan, R. Virus titre determines the efficiency of Pentalonia nigronervosa (Aphididae: Hemiptera) to transmit banana bunchy top virus. Virus Dis. 2018, 29, 499–505. [Google Scholar] [CrossRef]
- Rubinstein, G.; Czosnek, H. Long-term association of tomato yellow leaf curl virus with its whitefly vector Bemisia tabaci: Effect on the insect transmission capacity, longevity and fecundity. J. Gen. Virol. 1997, 78, 2683–2689. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.N.; Naqvi, A.R.; Jan, A.T.; Mohd, Q.; Haq, R. Genetic diversity and possible evidence of recombination among Banana Bunchy Top Virus (BBTV) isolates. Int. Res. J. Microbiol. 2010, 1, 1–12. [Google Scholar]
- Watanabe, S.; Bressan, A. Tropism, compartmentalization and retention of banana bunchy top virus (Nanoviridae) in the aphid vector Pentalonia nigronervosa. J. Gen. Virol. 2013, 94, 209–219. [Google Scholar] [CrossRef]
- Shirima, R.R.; Maeda, D.G.; Kanju, E.; Ceasar, G.; Tibazarwa, F.I.; Legg, J.P. Absolute quantification of cassava brown streak virus mRNA by real-time qPCR. J. Virol. Methods 2017, 245, 5–13. [Google Scholar] [CrossRef]
- R. Core Team. R: A Language and Environment for Statistical Computing. 2013. Available online: https://cran.r-project.org/bin/windows/base/old/3.4.3/ (accessed on 30 November 2017).
- De Mendiburu Delgado, F. Una Herramienta de Análisis Estadístico Para la Investigación Agrícola; Repositorioslatino Americanos, National University of Engineering (Peru): 2009. Available online: https://repositorioslatinoamericanos.uchile.cl/handle/2250/2356485 (accessed on 4 August 2021).
- Wickham, H. The tidyverse. R Package Ver. 2017, 1, 836. [Google Scholar]
- Li, M.; Liu, J.; Liu, S.-S. Tomato yellow leaf curl virus infection of tomato does not affect the performance of the Q and ZHJ2 biotypes of the viral vector Bemisia tabaci. Insect Sci. 2010, 18, 40–49. [Google Scholar] [CrossRef]
- Czosnek, H.; Ghanim, M.; Morin, S.; Rubinstein, G.; Fridman, V.; Zeidan, M. Whiteflies: Vectors, and victims (?), of geminiviruses. Adv. Virus Res. 2001, 57, 291–322. [Google Scholar] [PubMed]
- Rajan, P. Biology of Pentalonia nigronervosa f. caladii van der Goot, vector of ‘katte’ disease of cardamom. J. Plant. Crops. 1981, 9, 34–41. [Google Scholar]
- Nancy, J. Movement of Banana Bunchy Top Virus and Its Relationship with Its Aphid Vector (Pentalonia nigronervosa). Master’s Thesis, Bharathidasan University, Tiruchirapalli, India, 2003. [Google Scholar]
- Kang, L.; Chen, B.; Wei, J.; Liu, T.-X. Roles of Thermal Adaptation and Chemical Ecology in Liriomyza Distribution and Control. Annu. Rev. Èntomol. 2009, 54, 127–145. [Google Scholar] [CrossRef]
- Hoffmann, A.A.; Sorensen, J.G.; Loeschcke, V. Adaptation of Drosophila to temperature extremes: Bringing together quanti-tative and molecular approaches. J. Therm. Biol. 2003, 28, 175–216. [Google Scholar] [CrossRef]
- Gu, J.; Huang, L.X.; Shen, Y.; Huang, L.H.; Feng, Q.L. Hsp70 and small Hsps are the major heat shock protein members in-volved in midgut metamorphosis in the common cutworm, Spodoptera litura. Insect Mol. Biol. 2012, 5, 535–543. [Google Scholar] [CrossRef]
- Hayward, S.A.L.; Pavlidesb, S.C.; Tammariellob, S.P.; Rineharta, J.P.; Denlinger, D.L. Temporal expression patterns of dia-pause-associated genes in flesh fly pupae from the onset of diapause through post-diapause quiescence. J. Insect. Physiol. 2005, 51, 631–640. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhao, Q.; Duan, X.; Song, C.; Chen, M. Transcription of four Rhopalosiphum padi (L.) heat shock protein genes and their responses to heat stress and insecticide exposure. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2017, 205, 48–57. [Google Scholar] [CrossRef]
- Lü, Z.-C.; Wan, F.-H. Differential gene expression in whitefly (Bemisia tabaci) B-biotype females and males under heat-shock condition. Comp. Biochem. Physiol. Part D Genom. Proteom. 2008, 3, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Porras, M.F.; Navas, C.A.; Marden, J.H.; Mescher, M.C.; De Moraes, C.M.; Pincebourde, S.; Sandoval-Mojica, A.; Raygoza-Garay, J.A.; Holguin, G.A.; Rajotte, E.G.; et al. Enhanced heat tolerance of viral-infected aphids leads to niche expansion and reduced interspecific competition. Nat. Commun. 2020, 11, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Xu, N.; Zhong, T.; Feng, W.; Zhou, G. Tolerance and responsive gene expression of Sogatella furcifera under extreme temperature stresses are altered by its vectored plant virus. Sci. Rep. 2016, 6, 31521. [Google Scholar] [CrossRef] [Green Version]
- Van Munster, M. Impact of abiotic stresses on plant virus transmission by aphids. Viruses 2020, 12, 216. [Google Scholar] [CrossRef] [Green Version]
- Wu, R.Y.; Su, H.J. Transmission of banana bunchy top virus by aphids to banana plantlets from tissue culture. Bot. Bull. Acad. Sin. 1990, 31, 7–10. [Google Scholar]
- Ghosh, A.; Chakrabarti, S.; Mandal, B.; Kumar, N.K. Aphids as vectors of the plant viruses in India. In A Century of Plant Virology in India, 1st ed.; Mandal, B., Govind Pratap, R., Virendram, K.B., Rakesh, K.J., Eds.; Springer: Singapore, 2017; pp. 515–536. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Serial No. | Gene | Primer Sequence | Product Size (~) |
|---|---|---|---|
| 1 | DNA-S (q-PCR) | F-5′GCTTCTTGTGTGCAAGGTGA′3 R-5′CCTCAACACGGTTGTCTTCC′3 | 226 |
| 2 | Hsp40 | F-5′GGGAAATGTCCGTCAAAGAA′3 R-5′GAGTTTCATGGTGTCGCCTA′3 | 206 |
| 3 | Hsp70 | F-5′TCTTTCCTCATCCACCCAAG′3 R-5′ACAATGTCGTTGATGGCAGA′3 | 186 |
| 4 | Hsp90 | F-5′TCCCTGTTGCATTGTCACAT′3 R-5′TTGTCGTTGCAATCAGCTTC′3 | 186 |
| Development Stage | Development Periods (Days) | Difference of Mean (Days) (NVr–Vr) | ||
| Nymph | Viruliferous (Vr) ± SE | Non-Viruliferous (NVr) ± SE | ||
| Instar 1 | 2.333 ± 0.333 a | 3 ± 0 b | 0.6667 | |
| Instar 2 | 2.666 ± 0.333 a | 3.6667 ± 0.333 b | 1.001 | |
| Instar 3 | 2.33 ± 0.333 a | 3.3333 ± 0.333 b | 1 | |
| Instar 4 | 2.667 ± 0.333 a | 3 ± 0.577 a | 0.3333 | |
| Adult | Apterous Aphid | 25.667 ± 0.667 a | 35.333 ± 0.333 b | 9.666 |
| Total | 35.66667 a | 48.33333 b | 12.663 | |
| Sl. No. | Temperature (°C) | Virus Titer (Viral Copies ± SE) | CT (Threshold Cycle) (Mean ± SE) | BBTD Incidence (%) | Time Taken for BBTD Symptom Expression (Days) (Mean ± SE) |
|---|---|---|---|---|---|
| 1 | 15 | 12.34 ± 0.27 | 28.32 ± 0.566 | 50 | 189.21 ± 9.19 |
| 2 | 25 | 116,203.91 ± 1244.40 | 15.21 ± 0.566 | 100 | 35.32 ± 8.73 |
| 3 | 38 | 371.20 ± 6.97 | 23.44 + 0.536 | 70 | 97.1 ± 7.55 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chakraborty, S.; Barman, M.; Samanta, S.; Roy, M.; Tarafdar, J. Effect of Banana Bunchy Top Virus on the Heat Shock Protein Genes of Pentalonia nigronervosa during Temperature Susceptibility and Its Effect on Virus Transmission. Agronomy 2021, 11, 1866. https://doi.org/10.3390/agronomy11091866
Chakraborty S, Barman M, Samanta S, Roy M, Tarafdar J. Effect of Banana Bunchy Top Virus on the Heat Shock Protein Genes of Pentalonia nigronervosa during Temperature Susceptibility and Its Effect on Virus Transmission. Agronomy. 2021; 11(9):1866. https://doi.org/10.3390/agronomy11091866
Chicago/Turabian StyleChakraborty, Swati, Mritunjoy Barman, Snigdha Samanta, Moupiya Roy, and Jayanta Tarafdar. 2021. "Effect of Banana Bunchy Top Virus on the Heat Shock Protein Genes of Pentalonia nigronervosa during Temperature Susceptibility and Its Effect on Virus Transmission" Agronomy 11, no. 9: 1866. https://doi.org/10.3390/agronomy11091866