
Breeding for Prolificacy, Total Carotenoids and Resistance to Downy Mildew in Small-Ear Waxy Corn by Modified Mass Selection

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
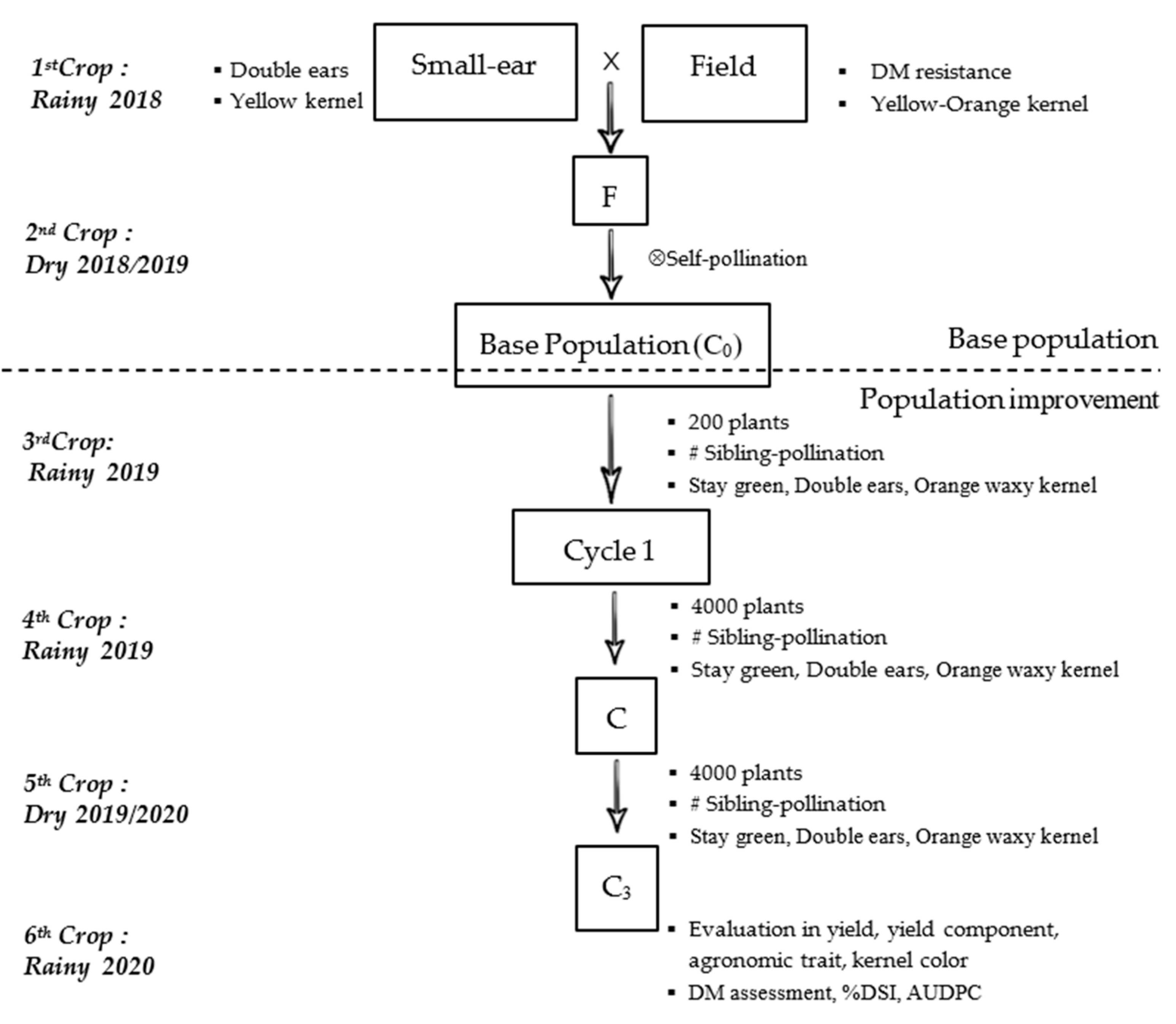
2.1. Plant Materials
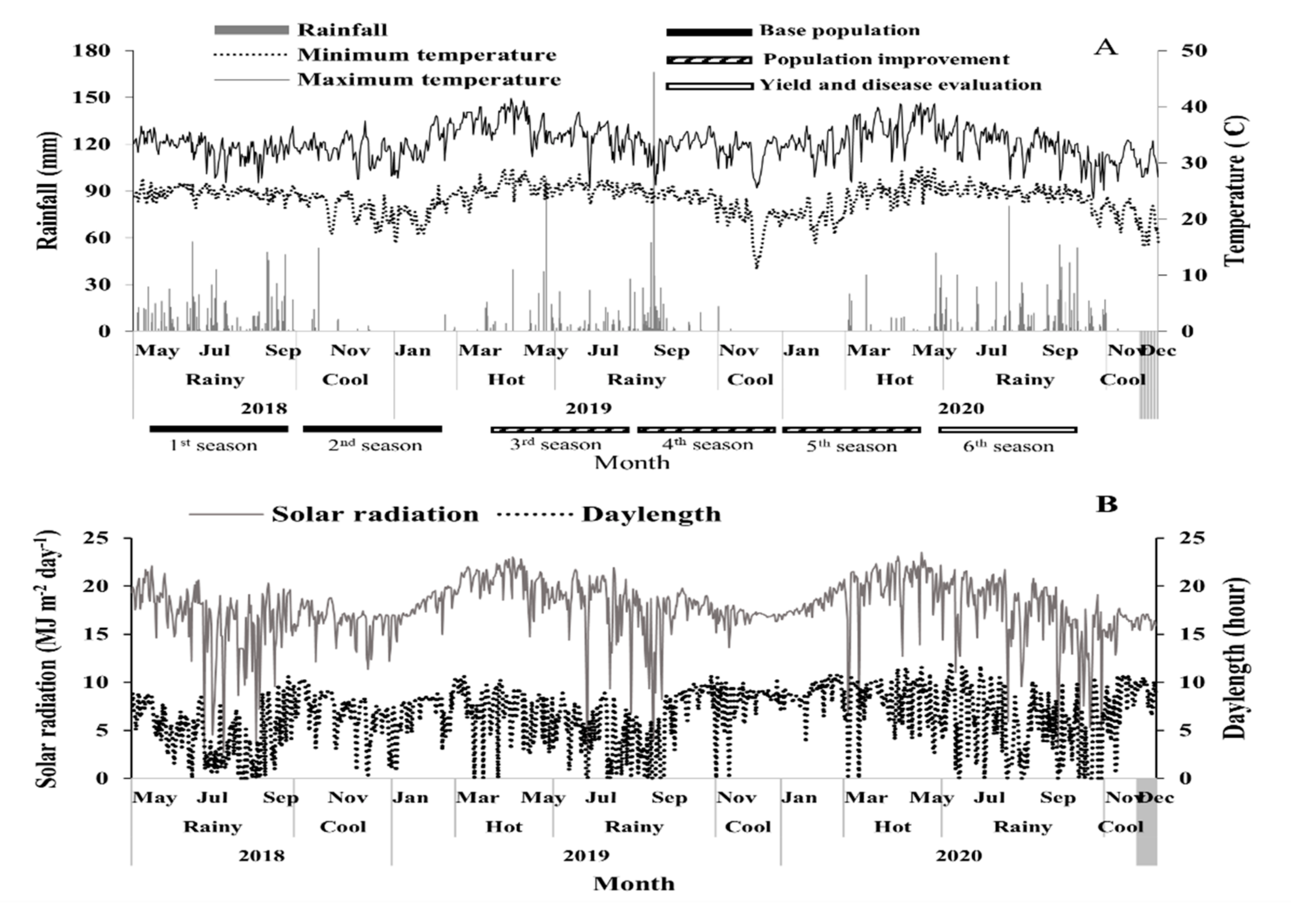
2.2. Base Population and Population Improvement
2.3. Yield Trial and Agronomic Evaluation
2.4. Experimental Design and Pathogen Inoculation
2.5. Disease Assessment
2.6. Statistical Analysis
3. Results and Discussion
3.1. Yield and Yield Components
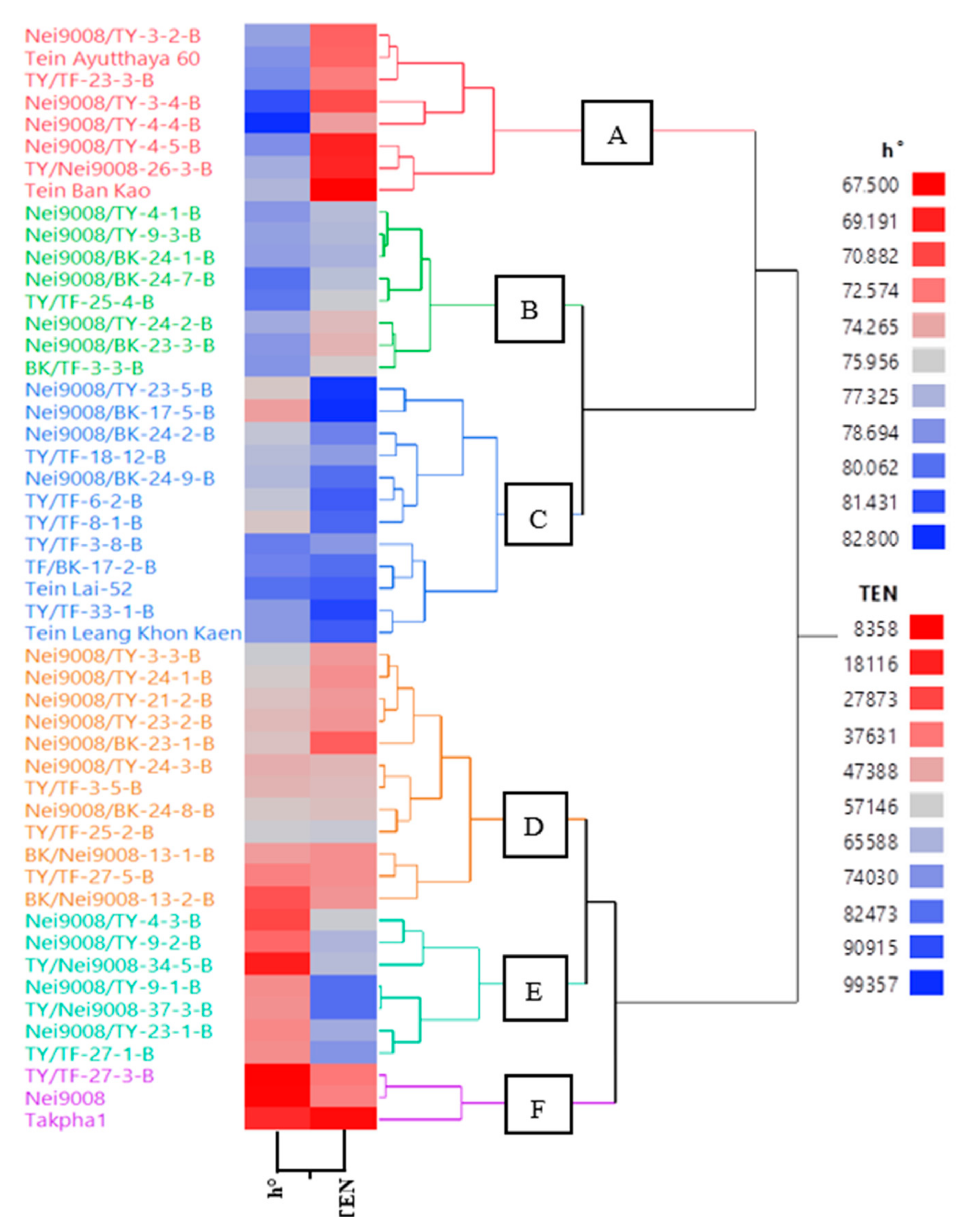
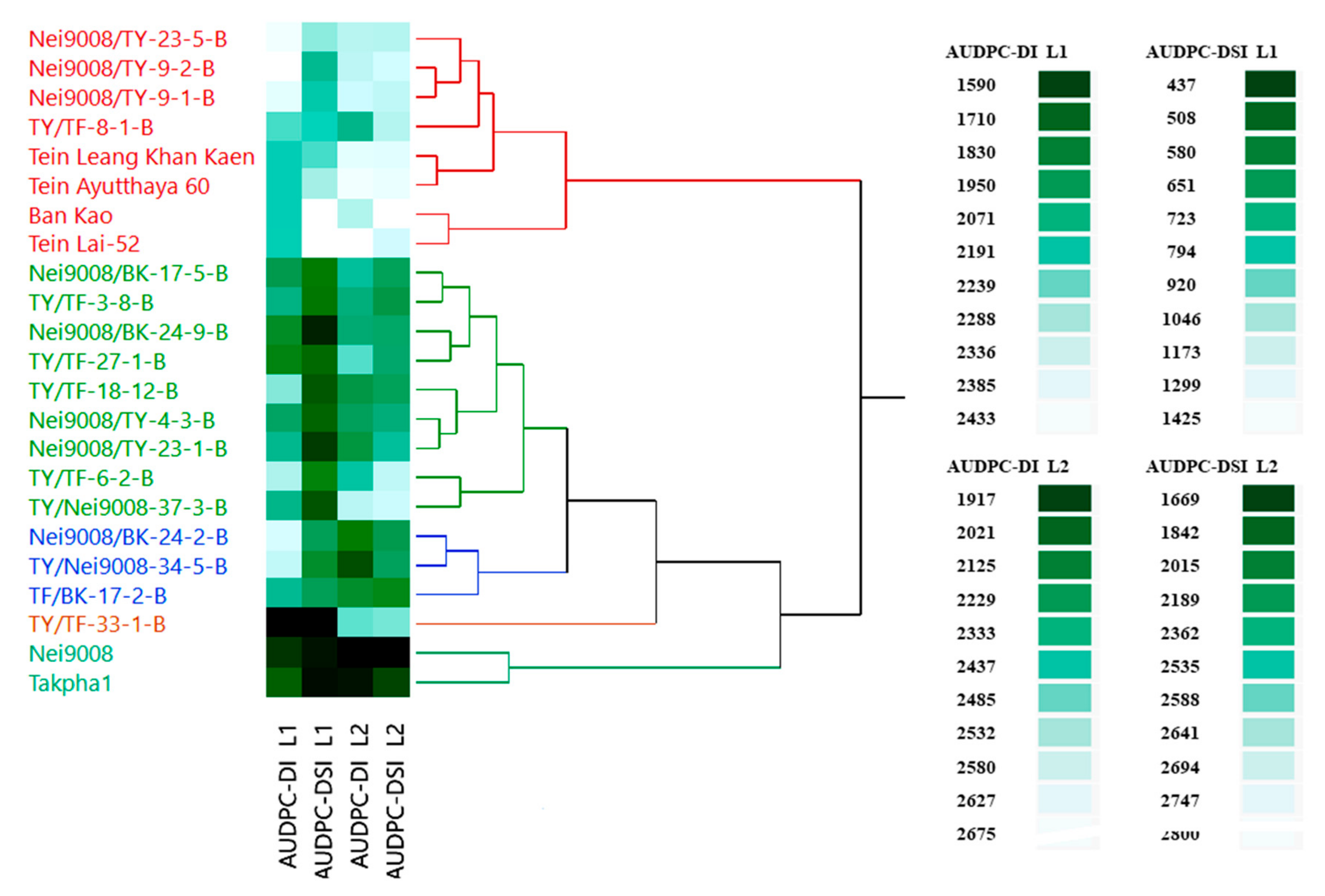
3.2. Cluster Analysis
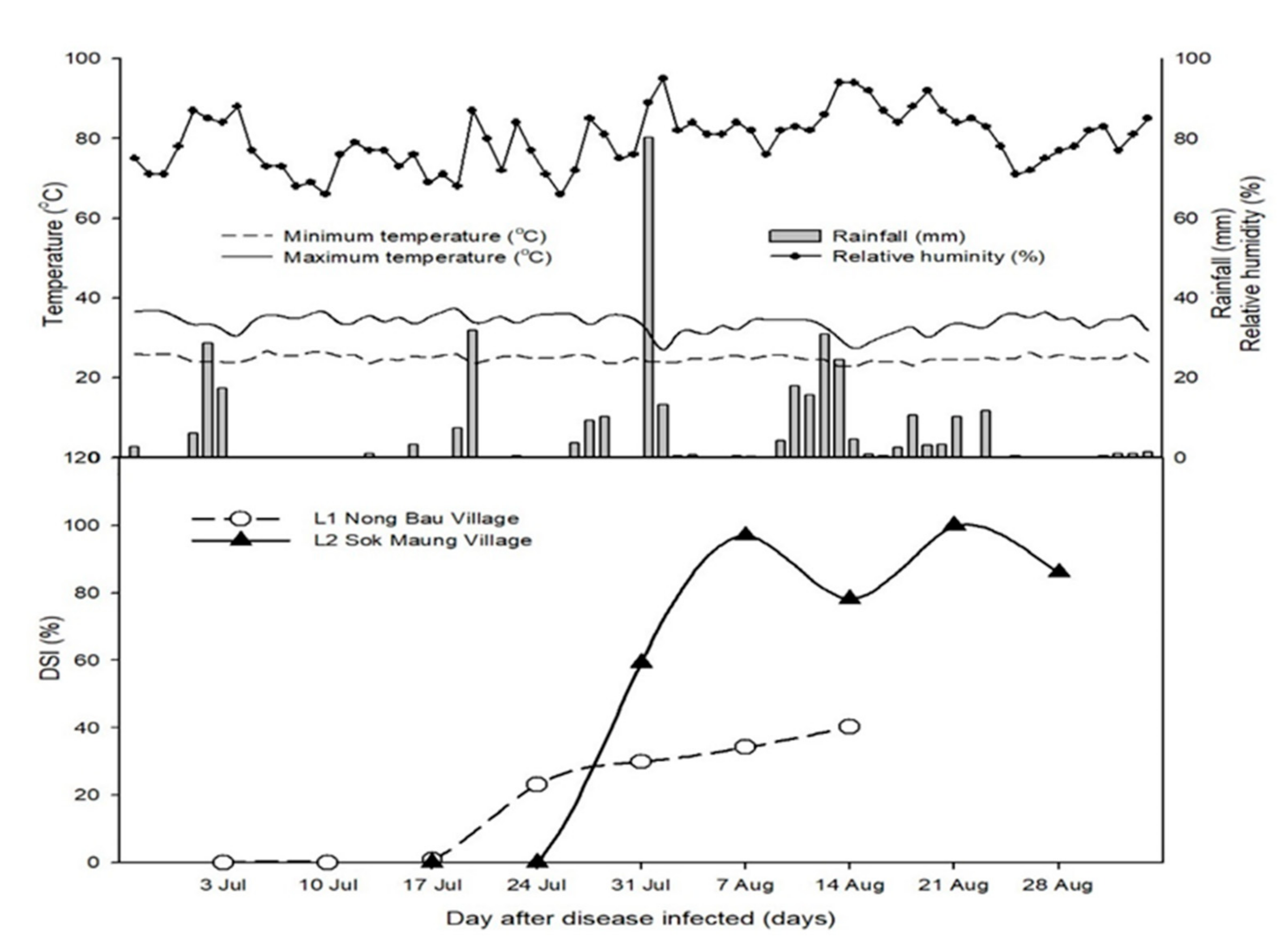
3.3. Disease Parameters
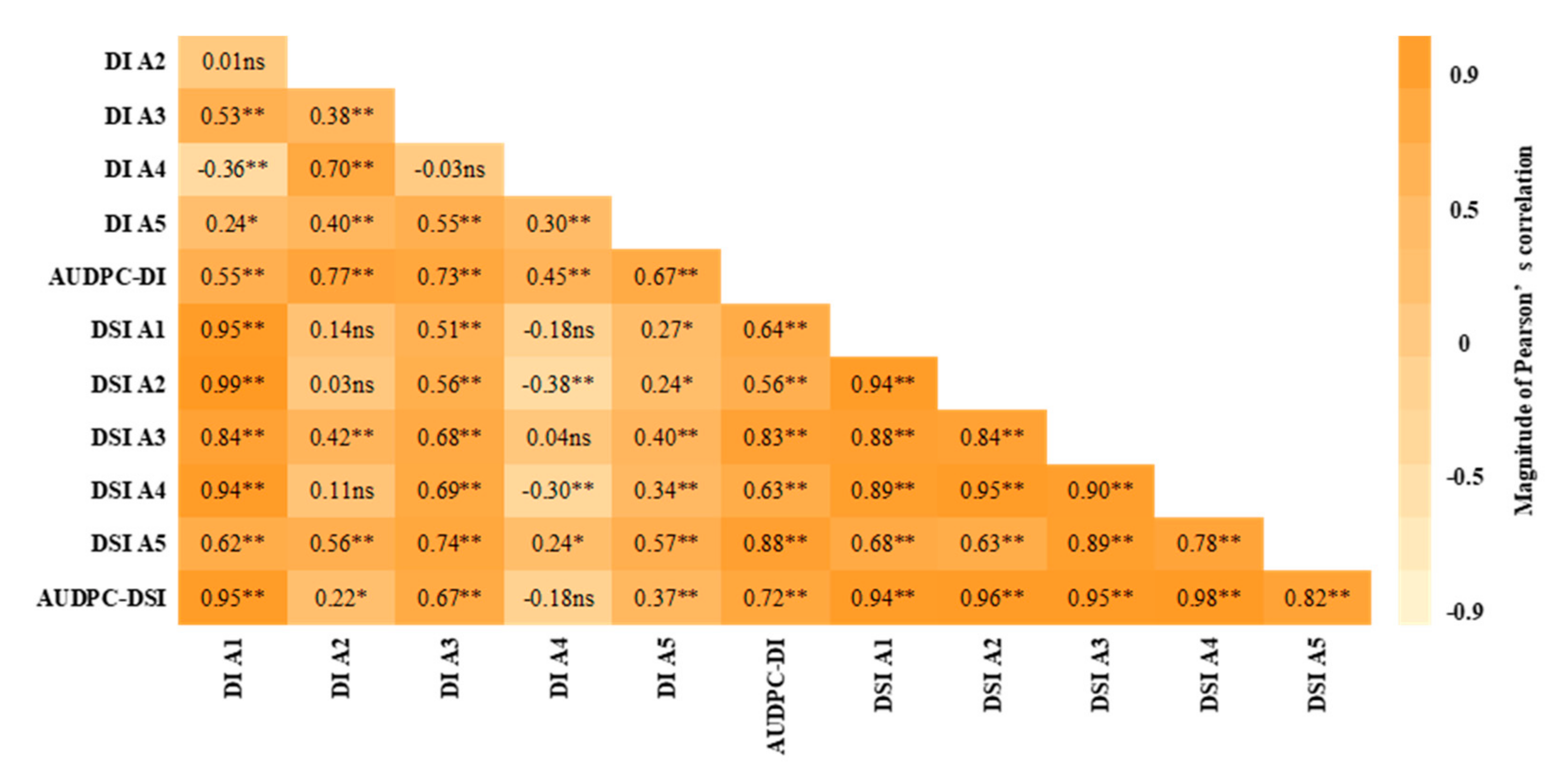
3.4. Correlation between Different Plant Ages for Disease Parameters
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xiaoyang, W.; Dan, C.; Yuqing, L.; Weihua, L.; Xinming, Y.; Xiuquan, L.; Juan, D.; Lihui, L. Molecular characteristics of two new waxy mutations in China waxy maize. Mol. Breed. 2017, 37, 27. [Google Scholar] [CrossRef]
- Fergason, V. High amylose and waxy corns. In Specialty Corns, 2nd ed.; Hallauer, A.R., Ed.; CRC Press: Boca Raton, FL, USA; London, UK; New York, NY, USA; Washington, DC, USA, 2001; pp. 71–92. [Google Scholar]
- Kesornkeaw, P.; Lertrat, K.; Suriharn, B. Response to four cycles of mass selection for prolificacy at low and high population densities in small ear waxy corn. Asian J. Plant Sci. 2009, 8, 425–432. [Google Scholar] [CrossRef]
- Chandler, K.; Lipka, A.E.; Owens, B.F.; Li, H.; Buckler, E.S.; Rocheford, T.; Gore, M.A. Genetic analysis of visually scored orange kernel color in maize. Crop Sci. 2013, 53, 189–200. [Google Scholar] [CrossRef] [Green Version]
- Exconde, O.R.; Raymundo, A.D. Yield loss caused by Philippine corn downy mildew. Philip. Agric. 1974, 58, 115–120. [Google Scholar]
- George, M.L.; Prasanna, C.B.M.; Rathore, R.S.; Setty, T.A.S.; Kasim, F.; Azrai, M.; Vasal, S.; Balla, O.; Hautea, D.; Canama, A.; et al. Identification of QTLs conferring resistance to downy mildews of maize in Asia. Theor. Appl. Genet. 2003, 107, 544–551. [Google Scholar] [CrossRef] [PubMed]
- Rashid, Z.; Zaidi, P.H.; Vinayan, M.T.; Sharma, S.S.; Srirama Setty, T.A. Downy mildew resistance in maize (Zea mays L.) across Peronosclerospora species in lowland tropical Asia. Crop Prot. 2013, 43, 183–191. [Google Scholar] [CrossRef]
- Janruang, P.; Unartngam, J. Morphological and molecular based identification of corn downy mildew distributed in Thailand. IJAT 2018, 14, 845–860. [Google Scholar]
- Kim, H.C.; Kim, K.H.; Song, K.; Kim, J.Y.; Lee, B.M. Identification and Validation of Candidate Genes Conferring Resistance to Downy Mildew in Maize (Zea mays L.). Genes 2020, 11, 191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lohithaswa, H.C.; Jyothi, K.; Kumar, K.S.; Hittalmani, S. Identification and introgression of QTLs implicated in resistance to sorghum downy mildew (Peronosclerospora sorghi (Weston and Uppal) C. G. Shaw) in maize through marker-assisted selection. J. Genet. 2015, 94, 741–748. [Google Scholar] [CrossRef]
- Premalatha, N.; Sundaram, K.M.; Arumugachamy, S. Screening and source of resistance to downy mildew (Peronosclerospora sorghi) in Maize (Zea Mays L.). Electron. J. Plant Breed. 2012, 3, 788–793. [Google Scholar]
- Sukto, S.; Lomthaisong, K.; Sanitchon, J.; Chankaew, S.; Scott, M.P.; Lubberstedt, T.; Lertrat, K.; Suriharn, B. Variability in Prolificacy, Total Carotenoids, Lutein, and Zeaxanthin of Yellow Small-Ear Waxy Corn Germplasm. Int. J. Agron. 2020, 2020, 8818768. [Google Scholar] [CrossRef]
- Kopsell, D.A.; Kopsell, D.E. Accumulation and bioavailability of dietary carotenoids in vegetable crops. Trends Plant Sci. 2006, 11, 499–507. [Google Scholar] [CrossRef]
- Nuss, E.T.; Arscott, S.A.; Bresnahan, K.; Pixley, K.V.; Rocheford, T.; Hotz, C.; Siamusantu, W.; Chileshe, J.; Tanumihardjo, S.A. Comparative intake of white- versus orange-colored maize by Zambian children in the context of promotion of biofortified maize. Food Nutr. Bull. 2012, 33, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Govender, L.; Pillay, K.; Siwela, M.; Modi, A.T.; Mabhaudhi, T. Improving the dietary vitamin A content of rural communities in South Africa by replacing non-biofortified white maize and sweet potato with biofortified maize and sweet potato in traditional dishes. Nutrients 2019, 11, 1198. [Google Scholar] [CrossRef] [Green Version]
- Khamkoh, W.; Ketthaisong, D.; Lomthaisong, K.; Lertrat, K.; Suriharn, B. Recurrent selection method for improvement of lutein and zeaxanthin in orange waxy corn populations. Aust. J. Crop Sci. 2019, 13, 566–573. [Google Scholar] [CrossRef]
- Harris, R.E.; Moll, R.H.; Stuber, C.W. Control and inheritance of prolificacy in maize. Crop Sci. 1976, 16, 843–850. [Google Scholar] [CrossRef]
- Hallauer, A.R. Heritability of prolificacy in maize. J. Hered. 1974, 65, 163–168. [Google Scholar] [CrossRef]
- Laible, C.A.; Dirks, V.A. Genetic variance and selective value of ear number in corn (Zea mays L.). Crop Sci. 1968, 8, 540–543. [Google Scholar] [CrossRef]
- Sorrells, M.E.; Lonnquist, J.H.; Harris, R.E. Inheritance of prolificacy in maize. Crop Sci. 1979, 19, 301–306. [Google Scholar] [CrossRef]
- Ali, S.E.; Saleh, G.B. Response of two cycles of phenotypic mass selection and heritability on two tropical sweet corn (Zea mays L. saccharata) populations. Asian J. Plant Sci. 2003, 2, 65–70. [Google Scholar] [CrossRef] [Green Version]
- Aday, B.A. The Philippine Program in Breeding for Resistance to Downy Mildew of Maize. 1980, pp. 207–219. Available online: https://www.jircas.go.jp/sites/default/files/publication/tars/tars8-_207-219.pdf (accessed on 31 May 2021).
- Khehra, A.S.; Dhillon, B.S.; Saxena, V.K.; Malhi, N.S. Selection for brown stripe downy mildew resistance in maize. Euphytica 1981, 30, 393–396. [Google Scholar] [CrossRef]
- Chander, S.; Guo, Y.Q.; Yang, X.H.; Zhang, J.; Lu, X.Q.; Yan, J.B.; Li, J.S. Using molecular markers to identify two major loci controlling carotenoid contents in maize grain. Theor. Appl. Genet. 2008, 116, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Halilu, A.D.; Adu, S.G.; Aba, D.A.; Usman, I.S. Genetics of carotenoids for provitamin a biofortification in tropical-adapted maize. Crop J. 2016, 4, 313–322. [Google Scholar] [CrossRef] [Green Version]
- Maita, R.; Coors, J.G. Twenty cycles of biparental mass selection for pollinated in the open-pollinated maize population golden glow. Crop Sci. 1996, 36, 1527–1532. [Google Scholar] [CrossRef]
- Hallauer, A.R.; Carena, M.J.; Miranda, F.J.B. Quantitative Genetics in Maize Breeding; Springer: New York, NY, USA, 2010; pp. 577–653. [Google Scholar]
- Zhang, S.H.; Shi, D.S.; Xu, J.S.; Yang, Y.F.; Kang, J.W.; Wang, L.M. Effects of mass selection on adaptive improvement of exotic quality protein maize populations: I. Direct response to selection for early silking. Zhongguo Nong Ye Ke Xue 1995, 21, 271–280. (In Chinese) [Google Scholar]
- Falke, K.C.; Flachenecker, C.; Melchinger, A.E.; Piepho, H.P.; Maurer, H.P.; Frisch, M. Temporal changes in allele frequencies in two European F(2) flint maize populations under modified recurrent full-sib selection. Theor. Appl. Genet. 2007, 114, 765–776. [Google Scholar] [CrossRef] [PubMed]
- Yong, H.; Zhang, F.; Tang, J.; Yang, Z.; Zhao, X.; Li, M.; Zhang, D.; Hao, Z.; Weng, J.; Li, X. Breeding potential of inbred lines derived from five maizes (Zea mays L.) populations. Euphytica 2019, 215, 1–2. [Google Scholar] [CrossRef]
- Senamontry, K.; Lertrat, K.; Suriharn, B. Response to five cycles of modified mass selection for ear length in waxy corn. SABRAO J. Breed. Genet. 2013, 45, 332–340. [Google Scholar]
- Hussanun, S.; Suriharn, B.; Lertrat, K. Yield and early maturity response to four cycles of modified mass selection in purple waxy corn. Turk. J. Field Crop. 2014, 19, 84–89. [Google Scholar] [CrossRef] [Green Version]
- Yong, H.; Tang, J.; Zhao, X.; Zhang, F.; Yang, Z.; Li, Y.; Li, M.; Zhang, D.; Hao, Z.; Weng, J.; et al. Effect of five modified mass selection cycles on combining ability in two Chinese maize populations. Euphytica 2020, 216, 57. [Google Scholar] [CrossRef]
- Grudloyma, P.; Noradechanon, S.; Papintu, K. Maize Research and Development for Downy Mildew Resistance. Volume 11. Unpublished. Available online: https://www.lib.ku.ac.th/KU/CR000320040005.pdf (accessed on 31 May 2021).
- Gao, H.; Gadlage, M.J.; Lafitte, H.R.; Lenderts, B.; Yang, M.; Schroder, M.; Farrell, J.; Snopek, K.; Peterson, D.; Feigenbutz, L.; et al. Superior field performance of waxy corn engineered using CRISPR-Cas9. Nat. Biotechnol. 2020, 38, 579–581. [Google Scholar] [CrossRef]
- Mcguire, R.G. Reporting of objective color measurements. HortScience 1992, 27, 1254–1255. [Google Scholar] [CrossRef] [Green Version]
- Phumichai, C.; Chunwongse, J.; Jampatong, S.; Grudloyma, P.; Pulam, T.; Doungchan, W.; Wongkaew, A.; Kongsiri, N. Detection and integration of gene mapping of downy mildew resistance in maize inbred lines though linkage and association. Euphytica 2012, 187, 369–379. [Google Scholar] [CrossRef]
- Cardwell, J.M.; Kling, J.G.; Bock, C. Methods for screening maize against downy mildew Peronosclerospora sorghi. Plant Breed. 1997, 116, 221–226. [Google Scholar] [CrossRef]
- Raupach, G.S.; Liu, L.; Murphy, J.F.; Tuzun, S.; Kloepper, J.W. Induced Systemic Resistance in Cucumber and Tomato Against Cucumber Mosaic Cucumovirus Using Plant Growth-Promoting Rhizobacteria (PGPR). Plant Dis. 1996, 80, 891–894. [Google Scholar] [CrossRef]
- Anfok, G.H. Benzo-(1,2,3)-thiadiazole-7-carbothioic acid S-methyl ester induces systemic resistance in tomato (Lycopersicon esculentum. Mill cv. Vollendung) to cucumber mosaic virus. Crop Prot. 2000, 19, 401–405. [Google Scholar] [CrossRef]
- Craig, J.; Frederiksen, R.A. Pathotypes of Peronosclerospora sorghi. Plant Dis. 1980, 64, 778–779. [Google Scholar] [CrossRef]
- Nagabhushan, T.; Lohithaswa, H.C.; Sreeramasetty, T.A.; Puttaramanaik, H.S.; Hittalmani, S. Identification of stable source of resistance to sorghum downy mildew in maize (Zea mays L.). JANRM 2014, 1, 176–178. [Google Scholar]
- Das, M.K.; Rajaram, S.; Mundt, C.C.; Kronstad, W.E. Inheritance of slow-rusting resistance to leaf rust in wheat. Crop Sci. 1992, 32, 1452–1456. [Google Scholar] [CrossRef]
- STAR. Biometrics and Breeding Informatics; Version 2.0.1.; PBGB Division, International Rice Research Institute: Los Baños, Laguna, Philippine, 2014. [Google Scholar]
- Statistix 10 Analytical Software. 2105 Miller Landing Road, Tallahassee, Florida, USA. 2013. Available online: http://statistix.com (accessed on 11 March 2021).
- Sofi, P.; Rather, A.G. Genetic analysis of yield trails in local and CIMMYT inbred line crosses using Line × tester analysis in maize (Zea mays L.). Asian J. Plant Sci. 2006, 5, 1039–1042. [Google Scholar]
- Narro, L.; Pandey, S.; Crossa, J.; Leon, C.D.; Salazar, F. Using line × tester interaction for the formation of yellow maize synthetics tolerance to acid soils. Crop Sci. 2003, 43, 1717–1728. [Google Scholar] [CrossRef]
- Mohamed, G.; Lertrat, K.; Suriharn, B. Yield and yield components of purple waxy corn grown under different locations in Thailand. Khon Kaen Agric. J. 2016, 44, 155–166. [Google Scholar]
- Aprilianti, D.K.; Syukur, M.; Suwarno, W.B. Evaluation of yield components of new sweet corn hybrids in Bogor, Indonesia. J. Trop. Crop Sci. 2016, 3, 13–18. [Google Scholar] [CrossRef]
- Bozalan, N.K.; Karadeniz, F. Carotenoid profile, total phenolic content, and antioxidant activity of carrots. Int. J. Food Prop. 2011, 14, 1060–1068. [Google Scholar] [CrossRef]
- Kumi, F.; Badji, A.; Mwila, N.; Odong, T.; Ochwossemakula, M.; Tusiime, G.; Gibson, P.; Biruma, M.; Prom, L.K.; Cuevas, H.E.; et al. New sources of sorghum resistant genotypes to downy mildew disease in Uganda. Biodiversitas 2019, 20, 3391–3397. [Google Scholar] [CrossRef]
- Islam, W. Plant disease epidemiology: Diseasetriangle and forecasting mechanisms in highlights. Hosts Viruses 2018, 5, 7–11. [Google Scholar]
- Kamala, V.; Singh, S.D.; Bramel, P.J.; Manohar, R.D. Sources of resistance to sorghum downy mildew in wild and weedy sorghums. Crop Sci. 2002, 42, 1357–1360. [Google Scholar] [CrossRef]
- Prom, L.K.; Perumal, R.; Montes-Garcia, N.; Isakeit, T.; Odvody, G.N. Evaluation of Gambian and Malian sorghum germplasm against downy mildew pathogen, Peronosclerospora sorghi, in Mexico and the USA. J. Gen. Plant Pathol. 2014, 81, 24–31. [Google Scholar] [CrossRef]
- Bock, C.H.; Jeger, M.J.; Mughogho, L.K.; Cardwell, K.F.; Mtisi, E.; Kaula, G.; Mukansabimana, D. Variability of Peronosclerospora sorghi isolates from different geographic location and hosts in Africa. Mycol. Res. 2000, 104, 61–68. [Google Scholar] [CrossRef]
- Shetty, H.S.; Safeeulla, K.M. Effect of some environmental factors on the asexual phase of Peronosclerospora sorghi. Proc. Indian Acad. Sci. (Plant Sci.) 1981, 90, 45–51. [Google Scholar] [CrossRef]
- Ginting, C.; Prasetyo, J.; Dirmawati, S.R.; Timotiwu, I.P.B.; Widyastuti, T.M.; Chafisa, D.I.R.; Asyifa, A.; Setyowati, E.; Pasaribu, A.H.Z. Identification of maize downey mildew pathogen in Lampung and the effects of varieties and metalaxyl on disease incidence. Annu. Res. Rev. Biol. 2020, 5, 23–35. [Google Scholar] [CrossRef]
- Ajala, S.O.; Kling, J.G.; Kim, S.K.; Obajimi, A.O. Improvement of maize population for resistance to downy mildew. Plant Breed. 2003, 122, 328–333. [Google Scholar] [CrossRef]
- Borges, O.L. Diallel analysis of maize resistance to sorghum downy mildew. Crop Sci. 1987, 27, 178–180. [Google Scholar] [CrossRef]
- Ruswandi, D.; Raymundo, A.D.; Lantican, R.M.; Hautea, D.M.; Salazar, A.M.; Carpena, A.L. Components of quantitative resistance to Philippine downy mildew in maize. Zuriat 2002, 13, 20–26. [Google Scholar] [CrossRef]
- Tadesse, W.; Reents, H.J.; Hsam, S.L.K.; Zeller, F.J. Relationship of seedling and adult plant resistance and evaluation of wheat germplasm against tan spot (Pyrenophora tritici-repentis). Genet. Resour. Crop Evol. 2011, 58, 339–346. [Google Scholar] [CrossRef]
- Chongo, G.; Gossen, B.G. Effect of plant age on resistance to Ascochyta rabiei in chickpea. Can. J. Plant Pathol. 2001, 23, 358–363. [Google Scholar] [CrossRef]
- Pfeiffer, W.H.; McClafferty, B. Harvest plus: Breeding crops for better nutrition. Crop Sci. 2007, 47, S-88–S-105. [Google Scholar] [CrossRef]
- Bouis, H.E.; Welch, R.M. Bioforification-A sustainable agricultural strategy for reducing micronutrient malnutrition in the Global South. Crop Sci. 2010, 50, S20–S32. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Commercial Name | Code Name | Pedigree | Cultivar Type | Corn Type | Color Kernel | DMR | Source |
|---|---|---|---|---|---|---|---|
| Takpha1 | Nei452008 or TF | Pio.3003-3-2-B-3-1-4-BBB | Inbred line | Field corn | Orange | R | NSFCRC |
| Nakhon Sawan1 | Nei9008 | SW(MMS) C2F2 × Pop.28 | Inbred line | Field corn | Intense orange | R | NSFCRC |
| Tein Leang Khon Kaen | TY | Improved from Tein Leang Nhong Bae | OPV | Small-ear waxy corn | yellow | S | PBRCSA |
| Tein Ban Kao | BK | - | OPV | Small-ear waxy corn | yellow | S | RUTS |
| Source of Variation | Replication | Genotype | Error | CV (%) | |
|---|---|---|---|---|---|
| Df | 1 | 49 | 49 | ||
| Unhusked yield | 1st | 11.6 ** | 8.3 ** | 0.2 | 8.5 |
| total | 43.4 ** | 21.0 ** | 0.3 | 8.1 | |
| Husked yield | 1st | 6.8 ** | 2.1 ** | 0.1 | 11.3 |
| total | 17.2 ** | 5.3 ** | 0.2 | 11.7 | |
| Ear number 1/ | 1st | 258.9 ** | 31.4 ** | 13.9 | 31.7 |
| 2nd | 180.3 ** | 22.5 ** | 7.1 | 44.7 | |
| 3rd | 0.3 * | 1.1 ** | 0.1 | 66.3 | |
| total | 905.8 ** | 100.6 ** | 29.4 | 30.0 | |
| Husked ear diameter | 1st | 0.3 * | 0.2 ** | 0.1 | 8.3 |
| Husked ear length | 1st | 5.6 ns | 3.3 * | 2.0 | 11.9 |
| Days to anthesis | 185 ** | 53 ** | 21.1 | 7.9 | |
| Plant height | 9409 ** | 923 ** | 211.0 | 10.1 | |
| Ear height | 4134 ** | 497 ** | 97.0 | 13.3 |
| Varieties | Unhusked Yield (Tons ha−1) | Husked Yield (Tons ha−1) | Ear Diameter (cm) | Ear Length (cm) | Days to Anthesis | Plant Height (cm) | Ear Height (cm) | ||
|---|---|---|---|---|---|---|---|---|---|
| 1st | Total | 1st | Total | ||||||
| Group C | |||||||||
| Nei9008/TY-23-5-B | 5.8 e–h | 10.8 c–e | 2.9 e–h | 5.3 c–e | 2.8 c–n | 13.0 a–j | 54 h–m | 182 ab | 93 bc |
| Nei9008/BK-17-5-B | 4.7 j–n | 8.9 g–i | 1.9 n–u | 3.5 i–n | 2.6 h–n | 11.6 d–k | 56 g–m | 130 h–o | 68 f–q |
| Nei9008/BK-24-2-B | 4.7 k–n | 6.0 p–s | 2.6 g–j | 3.2 k–q | 2.7 g–n | 11.8 c–k | 51 lm | 154 b–j | 73 d–p |
| TY/TF-18-12-B | 4.1 m–q | 6.8 n–p | 2.2 j–q | 3.4 i–n | 2.9 b–k | 13.5 a–h | 61 a–k | 147 c–l | 83 b–i |
| Nei9008/BK-24-9-B | 12.1 a | 17.2 a | 3.4 cd | 6.1 bc | 3.0 b–i | 14.1 a–f | 57 e–m | 157 b–h | 74 c–p |
| TY/TF-6-2-B | 5.5 f–i | 10.8 de | 1.7 q–u | 3.9 g–l | 2.5 i–o | 11.3 e–k | 56 f–m | 157 b–h | 91 b–d |
| TY/TF-8-1-B | 7.7 c | 10.9 c–e | 3.6 c | 5.1 d–f | 3.2 a–f | 14.0 a–f | 55 g–m | 201 a | 125 a |
| TY/TF-3-8-B | 5.8 e–h | 8.1 h–l | 2.8 f–i | 4.3 f–h | 2.7 f–n | 13.4 a–h | 59 b–l | 159 b–g | 90 b–e |
| TF/BK-17-2-B | 3.8 o–r | 7.0 l–p | 2.0 m–s | 3.3 j–p | 2.4 m–o | 11.1 g–k | 57 e–m | 167 b–e | 79 b–m |
| TY/TF-33-1-B | 10.4 b | 14.5 b | 5.9 a | 8.0 a | 3.1 a–g | 14.9 ab | 54 h–m | 175 a–c | 95 b |
| Group E | |||||||||
| Nei9008/TY-4-3-B | 6.3 ef | 10.6 ef | 3.3 c–e | 5.3 c–e | 2.7 h–n | 13.4 a–h | 54 h–m | 144 d–m | 61 k–s |
| Nei9008/TY-9-2-B | 7.1 cd | 9.2 gh | 4.3 b | 5.5 b–d | 3.3 a–d | 14.4 a–d | 54 h–m | 169 b–e | 80 b–k |
| TY/Nei9008-34-5-B | 4.6 k–n | 8.0 i–m | 2.0 m–s | 3.3 j–p | 2.8 f–n | 12.5 b–k | 56 g–m | 169 b–e | 94 bc |
| Nei9008/TY-9-1-B | 6.6 de | 11.8 cd | 3.0 d–g | 5.9 bc | 3.0 a–h | 13.9 a–g | 57 e–m | 159 b–h | 87 b–f |
| TY/Nei9008-37-3-B | 6.3 e–g | 9.7 fg | 3.3 c–f | 5.1 d–f | 3.0 a–h | 13.1 a–j | 54 h–m | 171 b–d | 93 bc |
| Nei9008/TY-23-1-B | 4.3 l–o | 7.4 k–o | 2.4 h–n | 4.1 g–j | 2.9 c–k | 13.5 a–h | 61 a–k | 157 b–h | 85 b–g |
| TY/TF-27-1-B | 4.7 k–n | 8.6 g–j | 2.4 h–m | 4.6 e–g | 2.9 c–k | 13.2 a–i | 54 h–m | 163 b–f | 88 b–e |
| Nei9008 | 4.3 l–o | 7.1 l–o | 2.6 g–k | 4.0 g–k | 2.4 l–o | 14.1 a–e | 67 a–d | 111 n–p | 51 q–t |
| Takpha1 | 2.8 s–u | 3.1 y–A | 0.3 x | 0.4 y | 3.3 a–e | 12.5 a–k | 67 a–c | 109 op | 56 n–t |
| Tein Leang Khan Kaen | 4.0 m–q | 6.6 n–q | 2.5 g–l | 4.1 g–j | 2.8 e–n | 11.4 e–k | 48 m | 142 d–m | 75 c–n |
| Tein Ban Kao | 1.1 x | 1.1 C | 0.4 x | 0.4 y | 2.9 b–k | 13.9 a–g | 62 a–i | 96 p | 38 t |
| Tein Lai-52 | 4.1 m–q | 6.7 n–p | 2.6 g–k | 4.1 g–j | 2.9 c–m | 11.9 c–k | 48 m | 153 b–j | 78 b–m |
| Tein Ayutthaya 60 | 2.9 s–u | 3.9 w–y | 1.5 s–v | 2.0 u–w | 2.9 b–k | 11.8 c–k | 53 i–m | 126 i–o | 63 j–s |
| CV (%) | 8.5 | 8.1 | 11.3 | 11.7 | 8.3 | 11.1 | 7.9 | 10.1 | 13.3 |
| SOV | df | 14 Days | 21 Days | 28 Days | 35 Days | 42 Days | AUDPC |
|---|---|---|---|---|---|---|---|
| DI (%) of the day after infected | |||||||
| Location (L) | 1 | 212,496 ** | 65 ns | 2816 ** | 4788 ** | 411ns | 7,275,656 ** |
| Genotype (G) | 22 | 21 ns | 622 ** | 112 ** | 407 ** | 549 ** | 55,439 ** |
| L × G | 22 | 19 ns | 191 ** | 104 ** | 238 ** | 289 ** | 45,328 ** |
| Pooled error | 44 | 23 | 88 | 49 | 33 | 13 | 11,709 |
| CV (%) | 9.3 | 11.4 | 7.4 | 6.4 | 3.8 | 4.3 | |
| DSI (%) of the day after infected | |||||||
| Location (L) | 1 | 78,080 ** | 127,495 ** | 48,607 ** | 92,771 ** | 38,529 ** | 45,180,000 ** |
| Genotype (G) | 22 | 152 ** | 134 ns | 434 ** | 175 ** | 1191 ** | 362,561 ** |
| L × G | 22 | 120 ** | 138 ns | 128 ns | 168 ** | 333 ** | 105,808 ** |
| Pooled error | 44 | 50 | 120 | 83 | 40 | 71 | 29,444 |
| CV (%) | 23.2 | 18.1 | 16.7 | 9.2 | 13.3 | 11.5 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sukto, S.; Lomthaisong, K.; Sanitchon, J.; Chankaew, S.; Falab, S.; Lübberstedt, T.; Lertrat, K.; Suriharn, K. Breeding for Prolificacy, Total Carotenoids and Resistance to Downy Mildew in Small-Ear Waxy Corn by Modified Mass Selection. Agronomy 2021, 11, 1793. https://doi.org/10.3390/agronomy11091793
Sukto S, Lomthaisong K, Sanitchon J, Chankaew S, Falab S, Lübberstedt T, Lertrat K, Suriharn K. Breeding for Prolificacy, Total Carotenoids and Resistance to Downy Mildew in Small-Ear Waxy Corn by Modified Mass Selection. Agronomy. 2021; 11(9):1793. https://doi.org/10.3390/agronomy11091793
Chicago/Turabian StyleSukto, Supaporn, Khomsorn Lomthaisong, Jirawat Sanitchon, Sompong Chankaew, Shanerin Falab, Thomas Lübberstedt, Kamol Lertrat, and Khundej Suriharn. 2021. "Breeding for Prolificacy, Total Carotenoids and Resistance to Downy Mildew in Small-Ear Waxy Corn by Modified Mass Selection" Agronomy 11, no. 9: 1793. https://doi.org/10.3390/agronomy11091793