
Effect of Forage Plant Mixture and Biostimulants Application on the Yield, Changes of Botanical Composition, and Microbiological Soil Activity

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Weather Conditions
2.2. Experimental Design
2.3. Characteristics of Fertilizers with Biostimulants
2.4. Botanical Composition and Biomass Yield
2.5. Soil Sampling and Analysis
2.6. Statistical Analyses
3. Results
3.1. Weather Condition
3.2. Botanical Composition
3.3. Biomass Production
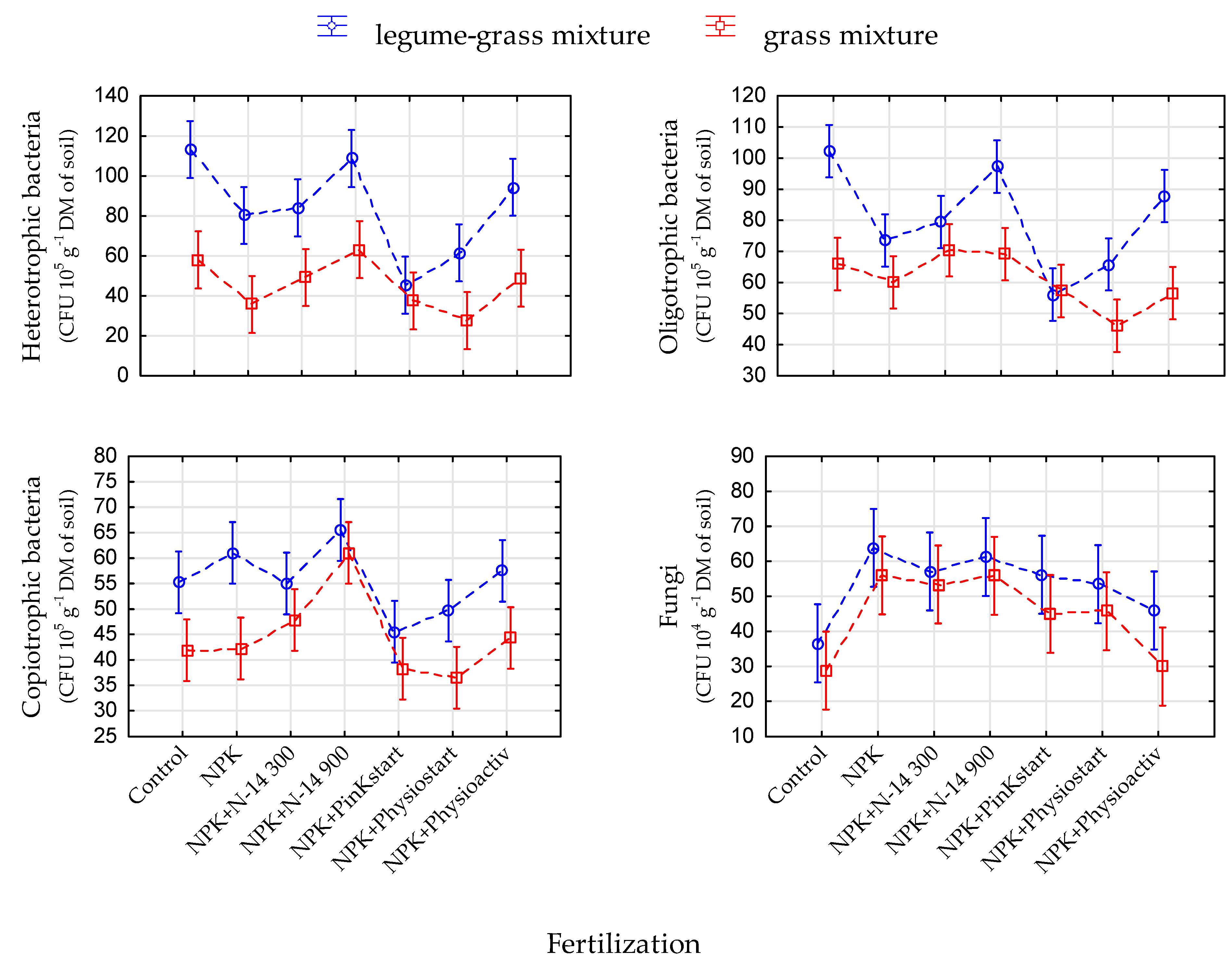
3.4. Soil pH and Counts of Microorganisms
3.5. Soil Enzymes Activity
4. Discussion
5. Conclusions
- Traditional NPK fertilization without biostimulants decreased the counts of most of the groups of soil microorganisms under analysis and the activity of soil enzymes, as compared with the non-fertilised absolute control variant. This effect was changed only in two cases, where the mineral NPK fertilization was accompanied by biostimulants, i.e., the N-14 at a dose of 900 kg·ha−1 and Physioactiv.
- The applied fertilizers had multidimensional and largely indirect influence on the soil microbiome and the activity of soil enzymes. It was mainly caused by the modification of the share of Trifolium repens in the sward and the pH of the soil environment. The effect of additional substances contained in the biostimulants seemed to be significant only at very high doses of these fertilizers—the N-14 applied at a dose of 900 kg·ha−1 was effective but it was ineffective at a dose of 300 kg·ha−1.
- The optimisation of the soil pH with CaCO3 applied at a dose increasing its value from 5.5 to 6.5 may reduce the negative effect of intensive nitrogen fertilization on the competitiveness of Trifolium repens against grasses. In our experiment this effect was observed after the application of the Physioactiv biostimulant. It is necessary to check whether the same effect can be observed after the application of other fertilizers containing calcium in the carbonate form.
- The effect of biostimulants on forage plants and soil microflora is not well understood yet. The investigation of new solutions and combinations of mineral nutrients with new biostimulants in fertilizers, which will affect plants and soil not only by optimizing soil pH are still needed.
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Aydin, I.; Uzun, F. Nitrogen and phosphorus fertilization of rangelands affects yield, forage quality and the botanical composition. Eur. J. Agron. 2005, 23, 8–14. [Google Scholar] [CrossRef]
- Hejcman, M.; Sochorová, L.; Pavlů, V.; Štrobach, J.; Diepolder, M.; Schellberg, J. The steinach grassland experiment: Soil chemical properties, sward height and plant species composition in three cut alluvial meadow after decades-long fertilizer application. Agric. Ecosyst. Environ. 2014, 184, 76–87. [Google Scholar] [CrossRef]
- Wolski, K.; Biernacik, M.; Świerszcz, S.; Talar-Krasa, M.; Leshchenko, O. Effect of the application of a biostimulant and mineral fertilizers on the concentration of mineral elements in the sward of forage mixtures cultivated on light soil. J. Elem. 2019, 24, 385–397. [Google Scholar]
- Dinka, M.O.; Dawit, M. Spatial Variability and Dynamics of Soil pH, Soil Organic Carbon and Matter Content: The Case of the Wonji Shoa Sugar-cane Plantation. J. Water Land Dev. 2019, 42, 59–66. [Google Scholar] [CrossRef] [Green Version]
- Turner, T.R.; James, E.K.; Poole, P.S. The plant microbiome. Genome Biol. 2013, 14, 209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swędrzyńska, D.; Zielewicz, W.; Swędrzyński, A. Comparison of soil bioconditioners and standard fertilization in terms of the impact on yield and vitality of Lolium perenne and soil biological properties. Open Life Sci. 2019, 14, 666–680. [Google Scholar] [CrossRef] [Green Version]
- Burzyńska, I. Monitoring of selected fertilizer nutrients in surface waters and soils of agricultural land in the river valley in central Poland. J. Water Land Dev. 2019, 43, 41–48. [Google Scholar] [CrossRef] [Green Version]
- Dobrzyński, J.; Wierzchowski, P.S.; Stępień, W.; Górska, E.B. The reaction of cellulolytic and potentially cellulolytic spore-forming bacteria to various types of crop management and farmyard manure fertilization in bulk soil. Agronomy 2021, 11, 772. [Google Scholar] [CrossRef]
- Wierzchowski, P.S.; Dobrzyński, J.; Mazur, K.; Kierończyk, M.; Wardal, W.J.; Sakowski, T.; Barszczewski, J. Chemical properties and bacterial community reaction to acidified cattle slurry fertilization in soil from maize cultivation. Agronomy 2021, 11, 601. [Google Scholar] [CrossRef]
- Shahrajabian, M.H.; Chaski, C.; Polyzos, N.; Petropoulos, S.A. Biostimulants application: A low input cropping management tool for sustainable farming of vegetables. Biomolecules 2021, 11, 698. [Google Scholar] [CrossRef] [PubMed]
- Sible, C.N.; Seebauer, J.R.; Below, F.E. Plant biostimulants: A categorical review, their implications for row crop production, and relation to soil health indicators. Agronomy 2021, 11, 1297. [Google Scholar] [CrossRef]
- Mendes, R.; Garbeva, P.; Raaijmakers, J.M. The rhizosphere microbiome: Significance of plant beneficial, plant pathogenic, and human pathogenic microorganisms. FEMS Microbiol. Rev. 2013, 37, 634–663. [Google Scholar] [CrossRef]
- Ahemad, M.; Kibret, M. Mechanisms and applications of plant growth promoting rhizobacteria: Current perspective. JKSUS 2014, 26, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Riggs, E.; Hobbie, S.E. Mechanisms driving the soil organic matter decomposition response to nitrogen enrichment in grassland soils. Soil Biol. Biochem. 2016, 99, 54–65. [Google Scholar] [CrossRef] [Green Version]
- Neufeld, K.R.; Garyston, S.J.; Bittman, S.; Krzie, M.; Hunt, D.E.; Smukler, S.M. Long-term alternative dairy manure management approaches enhance microbial biomass and activity in perennial forage grass. Biol. Fertil. Soils 2017, 53, 613–626. [Google Scholar] [CrossRef]
- Woźniak, M.; Gałązka, A. The rhizosphere microbiome and its beneficial effects on plants—Current knowledge and perspectives. Post. Microbiol. 2019, 58, 59–69. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.Q.; Tang, Y.H.; Jiang, J.; Yang, Y.H. Characterizing the dynamics of soil organic carbon in grasslands on the Qunghai-Tibetan Plateau. Sci. China Ser. D Earth Sci. 2007, 50, 113–120. [Google Scholar] [CrossRef]
- Zhong, W.H.; Cai, Z.C. Long-term effects of inorganic fertilizers on microbial biomass and community functional diversity in a paddy soil derived from quaternary red clay. Appl. Soil Ecol. 2007, 36, 84–91. [Google Scholar] [CrossRef]
- Swędrzyńska, D.; Małecka, I.; Blecharczyk, A.; Swędrzyński, A.; Starzyk, J. The effect of various long-term tillage systems on some chemical and biological properties of soil. Pol. J. Environ. Stud. 2013, 22, 1835–1844. [Google Scholar]
- Telesiński, A.; Krzyśko-Łupicka, T.; Cybulska, K.; Wróbel, J. Response of soil phosphatase activities to contamination with two types of tar oil. Environ. Sci. Pollut. Res. 2018, 25, 28642–28653. [Google Scholar] [CrossRef]
- Zuber, S.M.; Villamil, M.B. Meta-analysis approach to assess effect of tillage on microbial biomass and enzyme activities. Soil Biol. Biochem. 2016, 97, 176–187. [Google Scholar] [CrossRef] [Green Version]
- Salazar, S.; Sanchez, L.; Alvarez, J.; Valverde, A.; Galindo, P.; Igual, J.M.; Peix, A.; Santa-Regina, I. Correlation among soil enzyme activities under different forest system management practices. Ecol. Eng. 2011, 37, 1123–1131. [Google Scholar] [CrossRef]
- García, C.; Hernández, M.T.; Costa, F. Potential use of dehydrogenase activity as an index of microbial activity in degraded soils. Commun. Soil Sci. Plant Anal. 1997, 28, 123–134. [Google Scholar] [CrossRef]
- Kalembasa, S.; Symanowicz, B. Enzymatic activity of soil after applying various waste organic materials, ash, and mineral fertilizers. Pol. J. Environ. Stud. 2012, 21, 1635–1641. [Google Scholar]
- Lemanowicz, J.; Bartkowiak, A.; Breza-Boruta, B. Changes in phosphorus content, phosphatase activity and some physicochemical and microbiological parameters of soil within the range of impact of illegal dumping sites in Bydgoszcz. Environ. Earth Sci. 2016, 75, 510. [Google Scholar] [CrossRef] [Green Version]
- Swędrzyńska, D.; Zielewicz, W.; Swędrzyński, A.; Starzyk, J.; Wolna-Maruwka, A. Influence of the soleflor soil bioconditioner on the microbiological state of soil and the vigour and yield of orchard grass (Dactylis glomerata L.). Rocz. Ochr. Środ. 2015, 17, 1320–1338. [Google Scholar]
- Francioli, D.; Schulz, E.; Lentendu, G.; Wubet, T.; Buscot, F.; Reitz, T. Mineral vs. organic amendments: Microbial community structure, activity and abundance of agriculturally relevant microbes are driven by long-term fertilization strategies. Front. Microbiol. 2016, 7, 1446. [Google Scholar] [CrossRef] [Green Version]
- Gu, Y.; Wang, Y.; Lu, S.; Xiang, Q.; Yu, X.; Zhao, K.; Zou, L.; Chen, Q.; Tu, S.; Zhang, X. Long-term fertilization structures bacterial and archaeal communities along soil depth gradient in a paddy soil. Front. Microbiol. 2017, 8, 1516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rouphael, Y.; Colla, G. Toward a sustainable agriculture through plant biostimulants: From experimental data to practical applications. Agronomy 2020, 10, 1461. [Google Scholar] [CrossRef]
- Du Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Talar-Krasa, M.; Wolski, K.; Biernacik, M. Biostimulants and possibilities of their usage in grassland. Grassl. Sci. 2019, 65, 205–209. [Google Scholar] [CrossRef]
- Regulation (EU) 2019/1009 of the European Parliament and of the Council of 5 June 2019 Laying down Rules on the Making Available on the Market of EU Fertilising Products and Amending Regulations (EC) No 1069/2009 and (EC) No 1107/2009 and Repealing Regulation (EC) No 2003/2003. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A32019R1009 (accessed on 9 August 2021).
- Niewiadomska, A.; Sulewska, H.; Wolna-Maruwka, A.; Ratajczak, K.; Waraczewska, Z.; Budka, A. The influence of Bio-stimulants and foliar fertilizers on yield, plant features, and the level of soil biochemical activity in white lupine (Lupinus albus L.) Cultivation. Agronomy 2020, 10, 150. [Google Scholar] [CrossRef] [Green Version]
- Martyniuk, S.; Księżak, J. Evaluation of pseudo-microbial biopreparations used in crop production. Pol. J. Agron. 2011, 6, 27–33. [Google Scholar]
- Mayer, J.; Scheid, S.; Oberholzer, H.R. How effective are “Effective microorganisms”? Results from an organic farming field experiment. In Proceedings of the 16th JFOAM Organic World Congress, Modena, Italy, 18–20 June 2008; pp. 40–43. Available online: http//orgprints.org/14838 (accessed on 8 August 2021).
- Van Vliet, P.C.J.; Bloem, J.; de Goede, R.G.M. Microbial diversity, nitrogen loss and grass production after addition of Effective Microorganisms (EM) to slurry manure. Appl. Soil Ecol. 2006, 32, 188–198. [Google Scholar] [CrossRef]
- Yakhin, O.I.; Lubyanov, A.A.; Yakhin, I.A.; Brown, P.H. Biostimulants in plant science: A global perspective. Front. Plant. Sci. 2017, 7, 2049. [Google Scholar] [CrossRef] [Green Version]
- Craigie, J.S. Seaweed extract stimuli in plant science and agriculture. J. Appl. Phycol. 2011, 23, 371–393. [Google Scholar] [CrossRef]
- Khan, W.; Rayirath, U.P.; Subramanian, S.; Jithesh, M.N.; Rayorath, P.; Hodges, D.M.; Critchley, A.T.; Craige, J.S.; Norrie, J.; Prithiviraj, B. Seaweed extracts as biostimulants of plant growth and development. J. Plant. Growth Regul. 2009, 28, 386–399. [Google Scholar] [CrossRef]
- Renaut, S.; Masse, J.; Norrie, J.P.; Blal, B.; Hijri, M. A commercial seaweed extract structured microbial communities associated with tomato and pepper roots and significantly increased crop yield. Microb. Biotechnol. 2019, 12, 1346–1358. [Google Scholar] [CrossRef] [Green Version]
- Milton, R.F. Liquid seaweed as a fertilizer. Proc. Int. Seaweed Symp. 1964, 4, 428–431. [Google Scholar]
- González, A.; Castro, J.; Vera, J.; Moenne, A. Seaweed oligosaccharides stimulate plant growth by enhancing carbon and nitrogen assimilation, basal metabolism, and cell division. J. Plant. Growth Regul. 2013, 32, 443–448. [Google Scholar] [CrossRef] [Green Version]
- WRB. World reference base for soil resources. In Word Soil Resources Reports, 106; Food and Agriculture Organization of the United Nations: Rome, Italy, 2015. [Google Scholar]
- Marcinek, J.; Komisarek, J.; Bednarek, R.; Mocek, A.; Skiba, S.; Wiatrowska, K. Polish soil classification. Soil Sci. Annu. 2011, 62, 193. [Google Scholar]
- Zielewicz, W.; Goliński, P.; Wróbel, B. Effect of application of mineral fertilizers containing biological additives on botanical composition, nitrogen status and yielding of grass-legume sward. J. Res. Appl. Agric. Eng. 2018, 63, 146–150. [Google Scholar]
- Available online: https://pl.timacagro.com/ (accessed on 1 May 2021).
- Filipek, J. Zagadnienia wielkości próbek przeznaczonych do analizy botaniczno-wagowej w doświadczeniach łąkarskich (Issues of samples size intended for botanical-weight analysis in meadow experiments). Post. Nauk Rol. 1970, 50, 77–96. (In Polish) [Google Scholar]
- Novak, J. Evaluation of grassland quality. Ecologia 2004, 23, 127–143. [Google Scholar]
- Merck-Polska. 101621 Standard Count Agar for Microbiology. 2004, p. 1. Available online: https://www.merckmillipore.com/PL/pl/product/Standard-count-agar,MDA_CHEM-101621#anchor_TI (accessed on 10 November 2016).
- Hattori, R.; Hattori, T. Sensitivity to salts and organic compounds of soil bacteria isolated on diluted media. J. Gen. Appl. Microbiol. 1980, 26, 1–14. [Google Scholar] [CrossRef]
- Martin, J.P. Use of acid, rose bengal and streptomycin in the plate method for estimating soil fungi. Soil Sci. 1950, 69, 215–232. [Google Scholar] [CrossRef]
- Thalmann, A. The method of determining the dehydrogenase activity in the soil using triphenytetrazolium chloride (TTC) (Zur Methodik der Bestimmung der Dehydrogenase aktivität im Boden mittels triphenytetrazoliumchlorid (TTC)). Landwirtsch Forsch. 1968, 21, 249–258. [Google Scholar]
- Tabatabai, M.A.; Bremner, J.M. Use of p-nitrophenyl phosphate for assay of soil phosphatase activity. Soil Biol. Biochem. 1969, 1, 301–307. [Google Scholar] [CrossRef]
- Hoffman, G.; Teicher, K. Colorimetric method for determining the activity of the soil urease. J. Plant. Nutr. Fertil. Soil Sci. 1961, 95, 55–63. (In German) [Google Scholar]
- Johnson, J.L.; Temple, K.L. Some variables affecting the measurement of “catalase activity” in soil 1. Soil Sci. Soc. Am. 1964, 28, 207–209. [Google Scholar] [CrossRef]
- Stefanic, F.; Ellade, G.; Chirnageanu, J. Researches concerning a biological index of soil fertility. In Proceeding of the 5th Symposium of Soil Biology; Nemes, M.P., Kiss, S., Papacostea, P., Stefanic, C., Rusan, M., Eds.; Romanian National Society of Soil Science: Bucharest, Romania, 1984; pp. 35–45. [Google Scholar]
- Abdi, H.; Lynne, J.W. Principal component analysis. Wiley Interdiscip. Rev. Comput. Stat. 2010, 2, 433–459. [Google Scholar] [CrossRef]
- Jongman, R.H.G.; Bunce, R.G.H.; Metzger, M.J.; Mucher, C.A.; Howard, D.C.; Mateus, V.L. Objectives and applications of a statistical environmental stratification of Europe. Landsc. Ecol. 2006, 21, 409–429. [Google Scholar] [CrossRef]
- Berendsen, R.L.; Pieterse, C.M.J.; Bakker, P.A.H.M. The rhizosphere microbiome and plant health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef]
- Berg, G.; Grube, M.; Schloter, M.; Smalla, K. Unraveling the plant microbiome: Looking back and future perspectives. Front. Microbiol. 2014, 5, 148. [Google Scholar] [CrossRef] [Green Version]
- Bulgarelli, D.; Schlaeppi, K.; Spaepen, S.; Van Themaat, E.V.L.; Schulze-Lefert, P. Structure and functions of the bacterial microbiota of plants. Annu. Rev. Plant Biol. 2013, 64, 807–838. [Google Scholar] [CrossRef] [Green Version]
- Lakshmanan, V.; Selvaraj, G.; Bais, H.P. Functional soil microbiome: Belowground solutions to an aboveground problem. Plant Physiol. 2014, 166, 689–700. [Google Scholar] [CrossRef] [Green Version]
- Tshewang, S.; Rengel, Z.; Siddique, K.H.M.; Solaiman, Z.M. Nitrogen and potassium fertilisation influences growth, rhizosphere carboxylate exudation and mycorrhizal colonisation in temperate perennial pasture grasses. Agronomy 2020, 10, 1878. [Google Scholar] [CrossRef]
- Viljoen, C.; van der Colf, J.; Swanepoel, P.A. Benefits are limited with high nitrogen fertiliser rates in kikuyu-ryegrass pasture systems. Land 2020, 9, 173. [Google Scholar] [CrossRef]
- Dainese, M. Using natural gradients to infer a potential response to climate change: An example on the reproductive performance of Dactylis glomerata L. Biology 2012, 1, 857–868. [Google Scholar] [CrossRef]
- Lee, H.S.; Lee, I.D. Effect of N fertilizer levels on the dry matter yield, quality and botanical composition in eight-species mixtures. Korean J. Anim. Sci. 2000, 42, 727–734. [Google Scholar]
- Snyman, H.A. Short-term response of rangeland botanical composition and productivity to fertilization (N and P) in a semi-arid climate of South Africa. J. Arid Environ. 2002, 50, 167–183. [Google Scholar] [CrossRef]
- Kalmbacher, R.; Martin, F. Shifts in botanical composition of Flatwoods range following fertilization. J. Range Manag. 1996, 49, 530–534. [Google Scholar] [CrossRef] [Green Version]
- Samuel, M.J.; Hart, R.H. Nitrogen fertilization, botanical composition and biomass production on mixed-grass rangeland. J. Range Manag. 1998, 51, 408–416. [Google Scholar] [CrossRef] [Green Version]
- Silsbury, J.H.; Catchpoole, D.W.; Wallace, W. Effects of nitrate and ammonium on nitrogenase (C2H2 reduction) activity of swards of subterranean clover, Trifolium subterraneum L. Funct. Plant Biol. 1986, 13, 257–273. [Google Scholar] [CrossRef]
- Serraj, R.; Vadez, V.; Denison, R.F.; Sinclair, T.R. Involvement of ureides in nitrogen fixation inhibition in soybean. Plant Physiol. 1999, 119, 289–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saito, A.; Tanabata, S.; Tanabata, T.; Tajima, S.; Ueno, M.; Ishikawa, S.; Ohtake, N.; Sueyoshi, K.; Ohyama, T. Effect of nitrate on nodule and root growth of soybean (Glycine max (L.) Merr.). Int. J. Mol. Sci. 2014, 15, 4464–4480. [Google Scholar] [CrossRef] [PubMed]
- Baker, M.J.; Williams, W.M. White Clover; C.A.B International: Wallingford, UK, 1987. [Google Scholar]
- Rysiak, A.; Chabuz, W.; Sawicka-Zugaj, W.; Zdulski, J.; Grzywaczewski, G.; Kulik, M. Comparative impacts of grazing and mowing on the floristics of grasslands in the buffer zone of Polesie National Park, eastern Poland. Glob. Ecol. Conserv. 2020, 27, e01612. [Google Scholar] [CrossRef]
- Fulkerson, W.J.; Michell, P.J. The effect of height and frequency of mowing on the yield and composition of perennial ryegrass—White clover swards in the autumn to spring period. Grass Forage Sci. 1987, 42, 169–174. [Google Scholar] [CrossRef]
- Bolan, N.S.; Hedley, M.J.; White, R.E. Processes of soil acidification during nitrogen cycling with emphasis on legume based pastures. Plant Soil 1991, 134, 53–63. [Google Scholar] [CrossRef]
- Filipek, T.; Skowrońska, M. Current dominant causes and effects of acidification of soils under agricultural use in Poland. Acta Agroph. 2013, 20, 283–294. [Google Scholar]
- Grzebisz, W.; Diatta, J.B.; Szczepaniak, W. Production and ecological conditions of liming arable land soils (Produkcyjne i ekologiczne uwarunkowania wapnowania gleb gruntów ornych). Nawozy Nawoż. 2006, 2, 69–85. (In Polish) [Google Scholar]
- Zielewicz, W.; Swędrzyńska, D.; Swędrzyński, A. The effect of different doses of Soleflor soil improver and mineral fertilizers on botanical composition and yielding of grass-legume sward (Wpływ zróżnicowanych dawek polepszacza glebowego Soleflor i nawozów mineralnych na skład botaniczny i plonowanie runi motylkowato-trawiastej). Łąk. Pol./Grassl. Sci. Pol. 2015, 18, 267–279. (In Polish) [Google Scholar]
- Elgersma, A.; Hassink, J. Effects of white clover (Trifolium repens L.) on plant and soil nitrogen and soil organic matter in mixtures with perennial ryegrass (Lolium perenne L.). Plant Soil 1997, 197, 177–186. [Google Scholar] [CrossRef]
- Helios, W. Effect of white clover (Trifolium repens L.) undersowing cultivation and nitrogen fertilization on weed infestation, biomass yield and its component, content and uptake of macroelements of willow (Salix viminalis L.). Agronomy 2021, 11, 786. [Google Scholar] [CrossRef]
- Wolinska, A.; Frąc, M.; Oszust, K.; Szafranek-Nakonieczna, A.; Zielenkiewicz, U.; Stępniewska, Z. Microbial biodiversity of meadows under different modes of land use: Catabolic and genetic fingerprinting. World J. Microbiol. Biotechnol. 2017, 33, 154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teixeira, R.A.; Soares, T.G.; Fernandes, A.R.; de Souza Braz, A.M. Grasses and legumes as cover crop in no-tillage system in northeastern Pará Brazil. Acta Amaz. 2014, 44, 411–418. [Google Scholar] [CrossRef]
- Borkert, C.M.; Gaudêncio, C.A.; Pereira, J.E. Mineral nutrients in the aboveground biomass of ground cover crops (Nutrientes minerais na biomassa da parte aérea em culturas de cobertura do solo). Pesqui. Agropecu. Bras. 2003, 38, 143–153. [Google Scholar] [CrossRef]
- Swift, M.J.; Heal, O.W.; Anderson, J.M. Decomposition in Terrestrial Ecosystems; Blackwell Scientific Publication: Oxford, UK, 1979; p. 384. [Google Scholar]
- Bidar, G.; Garçon, G.; Pruvot, C.; Dewaele, D.; Cazier, F.; Douay, F.; Shirali, P. Behavior of Trifolium repens and Lolium perenne growing in a heavy metal contaminated field: Plant metal concentration and phytotoxicity. Environ. Pollut. 2007, 147, 546–553. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Hildago, P.; Hirsch, A.M. The nodule microbiome: N2-Fixing rhizobia do not live alone. Phytobiomes 2017, 1, 70–72. [Google Scholar] [CrossRef] [Green Version]
- Oleńska, E.; Małek, W. Genomic polymorphism of Trifolium repens root nodule symbionts from heavy metal-abundant 100-year-old waste heap in southern Poland. Arch. Microbiol. 2019, 201, 1405–1414. [Google Scholar] [CrossRef] [Green Version]
- Oleńska, E.; Małek, W. Sequence analysis of hypothetical lysine exporter genes of Rhizobium leguminosarum bv. trifolii from calamine old waste heaps and their evolutionary history. Curr. Microbiol. 2013, 66, 493–498. [Google Scholar] [CrossRef] [Green Version]
- Oleńska, E.; Małek, W. Genetic differentiation of Trifolium repens microsymbionts deriving from Zn–Pb waste-heap and control area in Poland. J. Basic Microbiol. 2015, 55, 462–470. [Google Scholar] [CrossRef]
- Hugenholtz, P.; Pace, N.R. Identifying microbial diversity in the natural environment: A molecular phylogenetic approach. Trends Biotechnol. 1996, 14, 190–197. [Google Scholar] [CrossRef]
- VanInsberghe, D.; Hartmann, M.; Stewart, G.R.; Mohn, W.W. Isolation of a substantial proportion of forest soil bacterial communities detected via pyrotag sequencing. Appl. Environ. Microbiol. 2013, 79, 2096–2098. [Google Scholar] [CrossRef] [Green Version]
- Niewiadomska, A.; Majchrzak, L.; Borowiak, K.; Wolna-Maruwka, A.; Waraczewska, Z.; Budka, A.; Gaj, R. The influence of tillage and cover cropping on soil microbial parameters and spring wheat physiology. Agronomy 2020, 10, 200. [Google Scholar] [CrossRef] [Green Version]
- Acosta-Martinez, V.; Tabatabai, M.A. Enzyme activities in a limed agricultural soil. Biol. Fertil. Soils 2000, 31, 85–91. [Google Scholar] [CrossRef]
- Błońska, E. Enzyme activity in forest peat soils. Folia For. Polonica Ser. A For. 2010, 52, 20–25. [Google Scholar]
- Fernández-Calviño, D.; Soler-Rovira, P.; Polo, A.; Díaz-Raviña, M.; Arias-Estévez, M.; Plaza, C. Enzyme activities in vineyard soils long-term treated with copper-based fungicides. Soil Biol. Biochem. 2010, 42, 2119–2127. [Google Scholar] [CrossRef]
- Włodarczyk, T.; Stępniewski, W.; Brzezińska, M. Dehydrogenase activity, redox potential, and emissions of carbon dioxide and nitrous oxide from Cambisols under flooding conditions. Biol. Fertil. Soils 2002, 36, 200–206. [Google Scholar] [CrossRef]
- Acosta-Martinez, V.; Tabatabai, M.A. Phosphorus cycle enzymes. In Methods of Soil Enzymology; Dick, R.P., Ed.; Soil Science Society of Ameriaca Inc.: Madison, WI, USA, 2011; pp. 161–182. [Google Scholar]
- Tautges, N.E.; Sullivan, T.S.; Reardon, C.L.; Burke, I.C. Soil microbial diversity and activity linked to crop yield and quality in a dryland organic wheat production system. Appl. Soil Ecol. 2016, 108, 258–268. [Google Scholar] [CrossRef] [Green Version]
- Abramczyk, K.; Gałązka, A. Diversity of soil microorganisms on areas under protection or with significant natural values. Sylwan 2017, 161, 496–503. (In Polish) [Google Scholar]
- Brock, J.L.; Kane, G.J. Variability in establishing white clover in pastures on farms. Proc. N. Z. Grassl. Assoc. 2003, 65, 223–228. [Google Scholar] [CrossRef]
- Caradus, J.R.; Woodfield, D. Overview and vision for white clover white clover: New Zealand’s competitive edge. Spec. Publ.-Agron. Soc. N. Z. 1995, 11, 1–6. [Google Scholar]
- Elgersma, A.; Schlepers, H.; Nassiri, M. Interactions between perennial ryegrass (Lolium perenne L.) and white clover (Trifolium repens L.) under contrasting nitrogen availability: Productivity, seasonal patterns of species composition, N2 fixation, N transfer and N recovery. Plant Soil 2000, 221, 281–299. [Google Scholar] [CrossRef]
- Schlueter, D.; Tracy, B. Sowing Method effects on clover establishment into permanent pasture. Agron. J. 2011, 104, 1217–1222. [Google Scholar] [CrossRef] [Green Version]
- Crush, J.R.; Nichols, S.N.; Ouyang, L. Adventitious root mass distribution in progeny of four perennial ryegrass (Lolium perenne L.) groups selected for root shape. N. Z. J. Agric. Res. 2010, 53, 193–200. [Google Scholar] [CrossRef] [Green Version]
- Fulkerson, W.J.; Donaghy, D.J. Plant-soluble carbohydrate reserves and senescence—Key criteria for developing an effective grazing management system for ryegrass-based pastures: A review. Aust. J. Exp. Agric. 2001, 41, 261–275. [Google Scholar] [CrossRef]
- Böhme, L.; Langer, U.; Böhme, F. Microbial biomass, enzyme activities and microbial community structure in two European long-term field experiments. Agric. Ecosyst. Environ. 2005, 109, 141–152. [Google Scholar] [CrossRef]
- Pietri, J.C.A.; Brookes, P.C. Substrate inputs and pH as factors controlling microbial biomass, activity and community structure in an arable soil. Soil Biol. Biochem. 2009, 41, 1396–1405. [Google Scholar] [CrossRef]
- Rousk, J.; Bååth, E.; Brookes, P.C.; Lauber, C.L.; Lozupone, C.; Caporaso, J.G.; Knight, R.; Fierer, N. Soil bacterial and fungal communities across a pH gradient in an arable soil. ISME J. 2010, 4, 1340–1351. [Google Scholar] [CrossRef]
- Wolińska, A.; Stępniewska, Z. Dehydrogenase activity in the soil environment. In Dehydrogenases; Canuto, R.A., Ed.; InTech: London, UK, 2012; pp. 183–210. [Google Scholar]
- Wolinska, A.; Stępniewska, Z.; Szymańska, E. Dehydrogenase activity of soil microorganisms and the total DNA level in soil of different use. J. Agric. Sci. Technol. B 2013, 3, 613–621. [Google Scholar]
- Praveen-Kumar, B.; Tarafdar, J.C. 2,3,5-Triphenyltetrazolinium chloride (TTC) as a electron acceptor of culturable soil bacteria, fungi, and actinomycetes. Biol. Fertil. Soils 2003, 38, 186–189. [Google Scholar] [CrossRef]
- Swędrzyńska, D.; Grześ, S. Microbiological parameters of soil under sugar beet as a response to the long-term application of different tillage systems. Pol. J. Environ. Stud. 2015, 24, 285–294. [Google Scholar] [CrossRef] [Green Version]
- Brzezińska, M.; Włodarczyk, T. Enzymes of intracellular redox transformations (oxidoreductases). Acta Agrophys. 2005, 3, 11–26. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
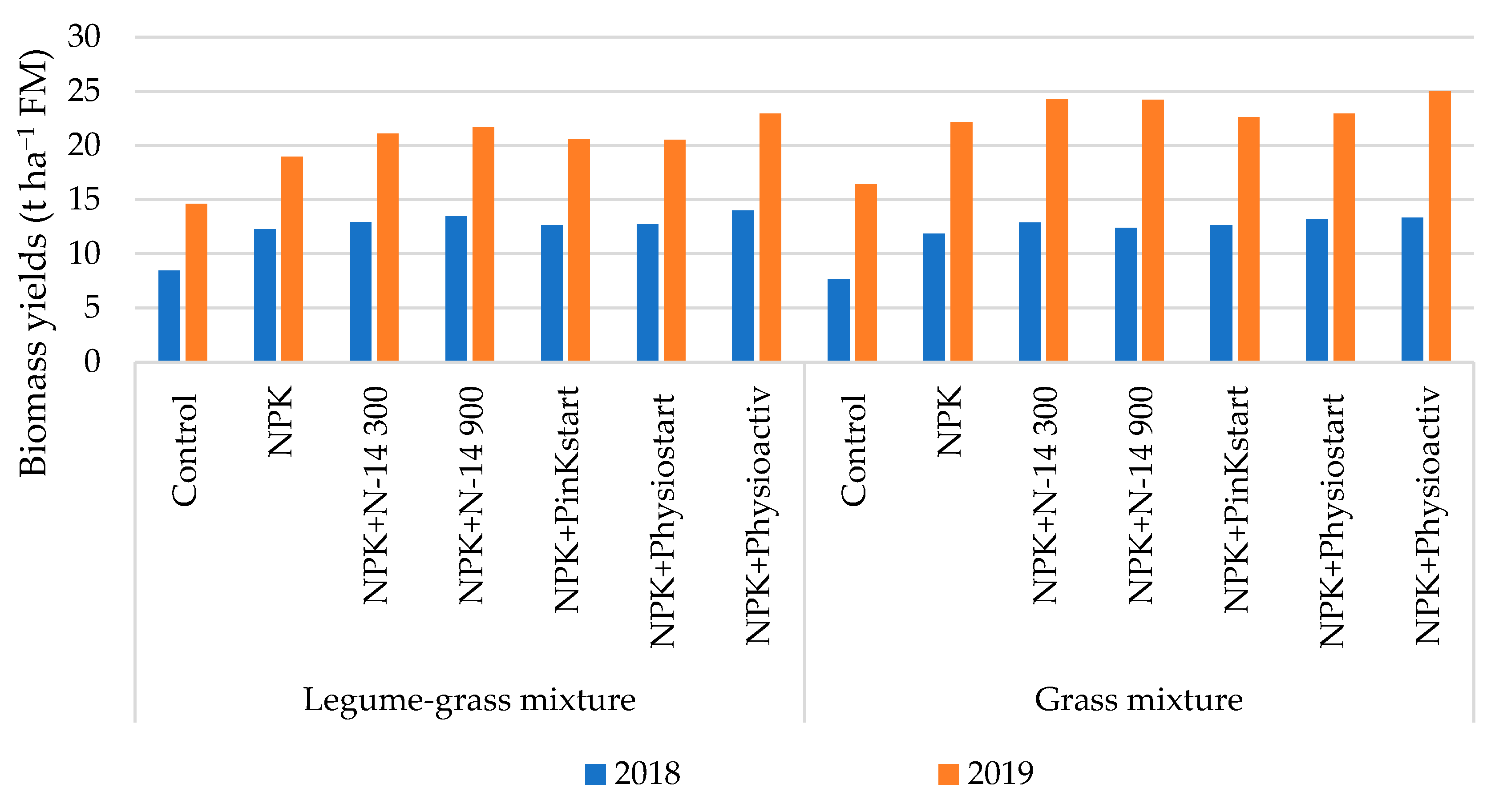{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mixture | Fertilization | Mixture × Fertiliz. | Error | Total | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| df = 1 | df = 6 | df = 6 | df = 154 | df = 167 | |||||||
| SS | F | p | SS | F | p | SS | F | p | SS | SS | |
| Lolium perenne | 342.9 | 15.2 | <0.001 | 4941.6 | 36.4 | <0.001 | 16,998.6 | 125.2 | <0.001 | 3485.5 | 25,768.6 |
| Other grasses | 24,432.6 | 783.7 | <0.001 | 21,329.3 | 114.7 | <0.001 | 6220.3 | 33.3 | <0.001 | 4801.2 | 56,783.4 |
| All grasses | 18,987.9 | 877.8 | <0.001 | 6353.6 | 49.0 | <0.001 | 3428.8 | 26.4 | <0.001 | 3331.2 | 32,098.5 |
| Trifolium repens | 21,488.1 | 1722.9 | <0.001 | 5915.3 | 79.0 | <0.001 | 4165.8 | 55.7 | <0.001 | 1920.7 | 33,489.9 |
| Study Year | Forage Plant Mixture | Fertilization Treatment (FT) | p-Values | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| GM | LGM | Control | NPK | NPK + N-14 300 | NPK + N-14 900 | NPK + PinKstart | NPK + Physiostart | NPK + Physioactiv | GM | FT | GM × FT | |
| 2018 | 12.0 a | 12.4 a | 8.1 a | 12.1 b | 12.9 b | 12.9 b | 12.6 b | 12.9 b | 13.7 b | ns | <0.001 | ns |
| 2019 | 22.5 a | 20.1 b | 15.5 a | 20.6 b | 22.7 bc | 23.0 bc | 21.6 bc | 21.7 bc | 24.0 c | <0.001 | <0.001 | ns |
| Mixture | Fertilization | Mixture × Fertilization | Error | Total | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| df = 1 | df = 6 | df = 6 | df = 154 | df = 167 | |||||||
| SS | F | p | SS | F | p | SS | F | p | SS | SS | |
| pH | 0.090 | 1.97 | ns | 13.30 | 48.42 | <0.001 | 0.123 | 0.45 | ns | 3.205 | 16.718 |
| Heterotrophic bacteria | 30,590.6 | 99.91 | <0.001 | 23,159.1 | 12.61 | <0.001 | 4160.0 | 2.26 | 0.047 | 21,433.8 | 79,346.6 |
| Oligotrophic bacteria | 16,013.4 | 73.43 | <0.001 | 18,819.4 | 14.38 | <0.001 | 6379.4 | 4.87 | 0.001 | 33,585.5 | 74,797.8 |
| Copiotrophic bacteria | 5130.5 | 45.39 | <0.001 | 7134.1 | 10.52 | <0.001 | 881.3 | 1.30 | ns | 17,405.7 | 30,551.6 |
| Fungi | 3049.8 | 7.98 | 0.005 | 15,393.9 | 6.72 | <0.001 | 588.6 | 0.26 | ns | 58,792.8 | 77,825.1 |
| Mixture | Fertilization | Mixture × Fertiliz. | Error | Total | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| df = 1 | df = 6 | df = 6 | df = 154 | df = 167 | |||||||
| SS | F | p | SS | F | p | SS | F | p | SS | SS | |
| Dehydrogenases | 51.93 | 9.58 | 0.003 | 587.01 | 18.06 | <0.001 | 113.29 | 3.49 | 0.005 | 379.23 | 1131.46 |
| Acid phosphatase | 0.0107 | 54.22 | <0.001 | 0.0344 | 2.90 | 0.014 | 0.0068 | 5.71 | 0.003 | 0.1342 | 0.0347 |
| Alkaline phosphatase | 0.0106 | 30.05 | <0.001 | 0.0262 | 11.32 | <0.001 | 0.0057 | 2.68 | 0.021 | 0.0248 | 0.0672 |
| Catalase | 29.074 | 33.20 | <0.001 | 28.362 | 5.40 | <0.001 | 22.409 | 4.26 | 0.001 | 44.55 | 99.990 |
| Urease | 0.950 | 0.003 | ns | 830.26 | 0.45 | ns | 194.37 | 0.01 | ns | 21,436.9 | 22,462.4 |
| Variable | pH | Biomass Yield | Lolium perenne | Other Grasses | All Grasses | Trifolium repens | Hetero. Bacteria | Oligo. Bacteria | Copio. Bacteria | Fungi | Dehydro. | Acid Phosphat. | Alkaline Phosphat. | Urease |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Biomass yield | 0.09 | |||||||||||||
| Lolium perenne | 0.05 | −0.13 | ||||||||||||
| Other grasses | −0.32 ** | 0.18 * | −0.66 ** | |||||||||||
| All grasses | −0.38 ** | 0.12 | 0.02 | 0.74 ** | ||||||||||
| Trifolium repens | 0.36 ** | −0.12 | −0.03 | −0.67 ** | −0.92 | |||||||||
| Heterotrophic bacteria | 0.37 ** | −0.05 | 0.11 | −0.61 ** | −0.72 | 0.69 ** | ||||||||
| Oligotrophic bacteria | 0.23 ** | −0.14 | −0.02 | −0.45 ** | −0.61 | 0.60 ** | 0.66 ** | |||||||
| Copiotrophic bacteria | 0.13 | −0.08 | −0.04 | −0.26 ** | −0.38 | 0.36 ** | 0.56 ** | 0.66 ** | ||||||
| Fungi | −0.29 ** | −0.11 | −0.20 ** | 0.14 | 0.01 | −0.02 | 0.14 | 0.16 * | 0.18 * | |||||
| Dehydrogenases | 0.57 ** | 0.03 | 0.22 ** | −0.51 ** | −0.48 | 0.46 ** | 0.21 ** | 0.24 ** | 0.04 | −0.45 ** | ||||
| Acid phosphat. | −0.14 | −0.23 ** | 0.28 ** | −0.38 ** | −0.25 | 0.24 ** | 0.26 ** | 0.15 | 0.26 ** | 0.19 * | 0.17 * | |||
| Alkaline phosphatase | 0.67 ** | 0.25 ** | 0.28 ** | −0.56 ** | −0.49 | 0.50 ** | 0.41 ** | 0.27 ** | 0.17 * | −0.23 ** | 0.63 ** | 0.12 | ||
| Urease | 0.23 ** | 0.59 ** | 0.06 | −0.15 | −0.14 | 0.15 * | 0.07 | 0.15 * | 0.07 | −0.42 ** | 0.44 ** | −0.27 ** | 0.45 ** | |
| Catalase | 0.52 ** | 0.16 * | −0.01 | −0.38 ** | −0.52 | 0.49 ** | 0.65 ** | 0.39 ** | 0.40 ** | 0.06 | 0.36 ** | 0.26 ** | 0.68 ** | 0.21 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zielewicz, W.; Swędrzyński, A.; Dobrzyński, J.; Swędrzyńska, D.; Kulkova, I.; Wierzchowski, P.S.; Wróbel, B. Effect of Forage Plant Mixture and Biostimulants Application on the Yield, Changes of Botanical Composition, and Microbiological Soil Activity. Agronomy 2021, 11, 1786. https://doi.org/10.3390/agronomy11091786
Zielewicz W, Swędrzyński A, Dobrzyński J, Swędrzyńska D, Kulkova I, Wierzchowski PS, Wróbel B. Effect of Forage Plant Mixture and Biostimulants Application on the Yield, Changes of Botanical Composition, and Microbiological Soil Activity. Agronomy. 2021; 11(9):1786. https://doi.org/10.3390/agronomy11091786
Chicago/Turabian StyleZielewicz, Waldemar, Arkadiusz Swędrzyński, Jakub Dobrzyński, Dorota Swędrzyńska, Iryna Kulkova, Paweł Stanisław Wierzchowski, and Barbara Wróbel. 2021. "Effect of Forage Plant Mixture and Biostimulants Application on the Yield, Changes of Botanical Composition, and Microbiological Soil Activity" Agronomy 11, no. 9: 1786. https://doi.org/10.3390/agronomy11091786