
Evaluation of Advanced Backcrosses of Eggplant with Solanum elaeagnifolium Introgressions under Low N Conditions

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Genotyping
2.3. Cultivation Conditions
2.4. Traits Evaluated
2.5. Data Analysis
2.6. QTL Detection
3. Results
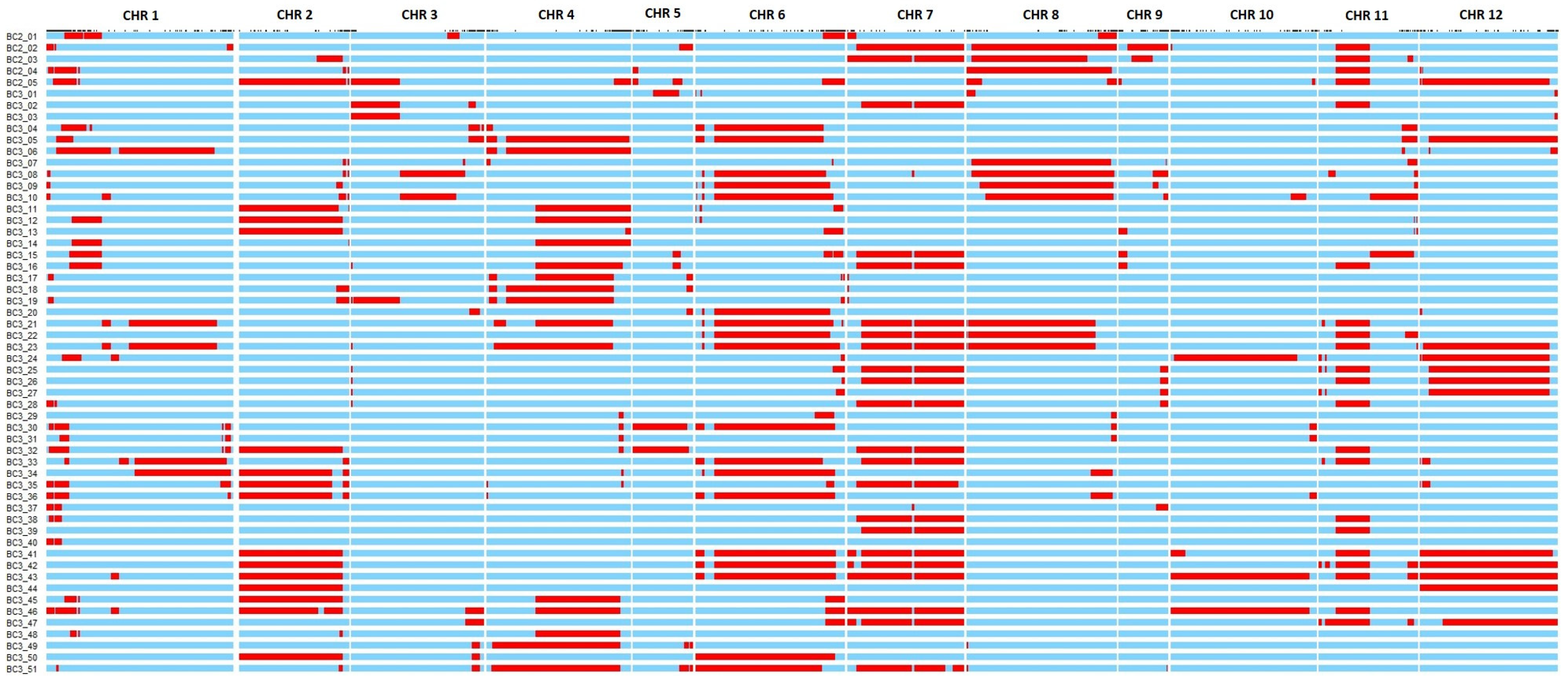
3.1. Genomic Characterization
3.2. Traits Evaluated
3.3. Principal Component Analysis
3.4. Correlations among Traits in the ABs
3.5. QTL Detection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marschner, P.; Rengel, Z. Nutrient availability in soils. In Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Elsevier Ltd.: Amsterdam, The Netherlands, 2011; pp. 315–330. ISBN 9780123849052. [Google Scholar]
- Xu, G.; Fan, X.; Miller, A.J. Plant nitrogen assimilation and use efficiency. Annu. Rev. Plant. Biol. 2012, 63, 153–182. [Google Scholar] [CrossRef] [Green Version]
- Albornoz, F. Crop responses to nitrogen overfertilization: A review. Sci. Hortic. 2016, 205, 79–83. [Google Scholar] [CrossRef]
- Zhang, X.; Davidson, E.A.; Mauzerall, D.L.; Searchinger, T.D.; Dumas, P.; Shen, Y. Managing nitrogen for sustainable development. Nature 2015, 528, 51–59. [Google Scholar] [CrossRef] [Green Version]
- Sharma, L.K.; Bali, S.K. A review of methods to improve nitrogen use efficiency in agriculture. Sustainability 2017, 10, 51. [Google Scholar] [CrossRef] [Green Version]
- Dempewolf, H.; Baute, G.; Anderson, J.; Kilian, B.; Smith, C.; Guarino, L. Past and future use of wild relatives in crop breeding. Crop. Sci. 2017, 57, 1070–1082. [Google Scholar] [CrossRef]
- Gao, Z.; Wang, Y.; Chen, G.; Zhang, A.; Yang, S.; Shang, L.; Wang, D.; Ruan, B.; Liu, C.; Jiang, H.; et al. The indica nitrate reductase gene OsNR2 allele enhances rice yield potential and nitrogen use efficiency. Nat. Commun. 2019, 10, 5207. [Google Scholar] [CrossRef]
- Prohens, J.; Gramazio, P.; Plazas, M.; Dempewolf, H.; Kilian, B.; Díez, M.J.; Fita, A.; Herraiz, F.J.; Rodríguez-Burruezo, A.; Soler, S.; et al. Introgressiomics: A new approach for using crop wild relatives in breeding for adaptation to climate change. Euphytica 2017, 213, 158. [Google Scholar] [CrossRef]
- Knapp, S.; Vorontsova, M.S.; Prohens, J. Wild relatives of the eggplant (Solanum melongena L.: Solanaceae): New understanding of species names in a complex group. PLoS ONE 2013, 8, e57039. [Google Scholar] [CrossRef] [Green Version]
- Chapman, M.A. Eggplant breeding and improvement for future climates. In Genomic Designing of Climate-Smart Vegatable Crops; Springer: Berlin/Heidelberg, Germany, 2020; pp. 257–276. ISBN 9783319933818. [Google Scholar]
- FAO. FAOSTAT Database Collections 2019; FAO: Rome, Italy, 2019. [Google Scholar]
- Plazas, M.; Vilanova, S.; Gramazio, P.; Rodríguez-Burruezo, A.; Fita, A.; Herraiz, F.J.; Ranil, R.; Fonseka, R.; Niran, L.; Fonseka, H.; et al. Interspecific hybridization between eggplant and wild relatives from different genepools. J. Am. Soc. Hortic. Sci. 2016, 141, 34–44. [Google Scholar] [CrossRef] [Green Version]
- Rotino, G.L.; Sala, T.; Toppino, L. Eggplant. In Alien Gene Transfer in Crop Plants; Springer: Berlin/Heidelberg, Germany, 2014; Volume 2, ISBN 978-1-4614-9571-0. [Google Scholar]
- Taher, D.; Solberg, S.Ø.; Prohens, J.; Chou, Y.; Rakha, M.; Wu, T. World Vegetable Center Eggplant Collection: Origin, Composition, Seed Dissemination and Utilization in Breeding. Front. Plant. Sci. 2017, 8, 1484. [Google Scholar] [CrossRef]
- García-Fortea, E.; Gramazio, P.; Vilanova, S.; Fita, A.; Mangino, G.; Villanueva, G.; Arrones, A.; Knapp, S.; Prohens, J.; Plazas, M. First successful backcrossing towards eggplant (Solanum melongena) of a New World species, the silverleaf nightshade (S. elaeagnifolium), and characterization of interspecific hybrids and backcrosses. Sci. Hortic. 2019, 246, 563–573. [Google Scholar] [CrossRef]
- Álvarez-Yépiz, J.C.; Martínez-Yrízar, A.; Búrquez, A.; Lindquist, C. Variation in vegetation structure and soil properties related to land use history of old-growth and secondary tropical dry forests in northwestern Mexico. For. Ecol. Manag. 2008, 256, 355–366. [Google Scholar] [CrossRef]
- Krigas, N.; Tsiafouli, M.A.; Katsoulis, G.; Votsi, N.; Kleunen, M. Van Investigating the Invasion Pattern of the Alien Plant Solanum. Plants 2021, 10, 805. [Google Scholar] [CrossRef]
- Knapp, S.; Sagona, E.; Carbonell, A.K.Z.; Chiarini, F. A revision of the Solanum elaeagnifolium clade (Elaeagnifolium clade; subgenus Leptostemonum, Solanaceae). PhytoKeys 2017, 84, 1–104. [Google Scholar] [CrossRef] [Green Version]
- Kaushik, P.; Gramazio, P.; Vilanova, S.; Raigón, M.D.; Prohens, J.; Plazas, M. Phenolics content, fruit flesh colour and browning in cultivated eggplant, wild relatives and interspecific hybrids and implications for fruit quality breeding. Food Res. Int. 2017, 102, 392–401. [Google Scholar] [CrossRef]
- Gramazio, P.; Prohens, J.; Plazas, M.; Mangino, G.; Herraiz, F.J.; García-Fortea, E.; Vilanova, S. Genomic tools for the enhancement of vegetable crops: A case in eggplant. Notulae Botanicae Horti Agrobotanici Cluj-Napoca 2018, 46, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Barchi, L.; Pietrella, M.; Venturini, L.; Minio, A.; Toppino, L.; Acquadro, A.; Andolfo, G.; Aprea, G.; Avanzato, C.; Bassolino, L.; et al. A chromosome-anchored eggplant genome sequence reveals key events in Solanaceae evolution. Sci. Rep. 2019, 9, 11769. [Google Scholar] [CrossRef] [PubMed]
- Gramazio, P.; Yan, H.; Hasing, T.; Vilanova, S.; Prohens, J.; Bombarely, A. Whole-genome resequencing of seven eggplant (Solanum melongena) and one wild relative (S. incanum) accessions provides new insights and breeding tools for eggplant enhancement. Front. Plant. Sci. 2019, 10, 1220. [Google Scholar] [CrossRef]
- Hirakawa, H.; Shirasawa, K.; Miyatake, K.; Nunome, T.; Negoro, S.; Ohyama, A.; Yamaguchi, H.; Sato, S.; Isobe, S.; Tabata, S.; et al. Draft genome sequence of eggplant (Solanum melongena L.) the representative Solanum species indigenous to the old world. DNA Res. 2014, 21, 649–660. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Qian, J.; Li, W.; Yu, N.; Gan, G.; Jiang, Y.; Li, W.; Liang, X.; Chen, R.; Mo, Y.; et al. A high-quality genome assembly of the eggplant provides insights into the molecular basis of disease resistance and chlorogenic acid synthesis. Mol. Ecol. Resour. 2021, 21, 1274–1286. [Google Scholar] [CrossRef]
- Wei, Q.; Wang, J.; Wang, W.; Hu, T.; Hu, H.; Bao, C. A high-quality chromosome-level genome assembly reveals genetics for important traits in eggplant. Hortic. Res. 2020, 7, 153. [Google Scholar] [CrossRef]
- Barchi, L.; Acquadro, A.; Alonso, D.; Aprea, G.; Bassolino, L.; Demurtas, O.; Ferrante, P.; Gramazio, P.; Mini, P.; Portis, E.; et al. Single Primer Enrichment Technology (SPET) for high-throughput genotyping in tomato and eggplant germplasm. Front. Plant. Sci. 2019, 10, 1005. [Google Scholar] [CrossRef] [Green Version]
- Vilanova, S.; Alonso, D.; Gramazio, P.; Plazas, M.; García-Fortea, E.; Ferrante, P.; Schmidt, M.; Díez, M.J.; Usadel, B.; Giuliano, G.; et al. SILEX: A fast and inexpensive high-quality DNA extraction method suitable for multiple sequencing platforms and recalcitrant plant species. Plant. Methods 2020, 16, 110. [Google Scholar] [CrossRef]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef] [PubMed]
- Van Reeuwijk, L. Procedures for Soil Analysis; International Soil Reference and Information Centre: Wageningen, The Netherlands, 2002; ISBN 90-6672-044-1. [Google Scholar]
- Yañez, J. Análisis de suelos y su interpretación. Hortic. Rev. Ind. Distrib. Y Socioecon. Hortícola Frutas Hortal. Flores Plantas Árboles Ornam. Y Viveros 1989, 49, 75–89. [Google Scholar]
- Moll, R.H.; Kamprath, E.J.; Jackson, W.A. Analysis and interpretation of factors which contribute to efficiency of nitrogen utilization 1. Agron. J. 1982, 74, 562–564. [Google Scholar] [CrossRef]
- Samonte, S.O.P.; Wilson, L.T.; Medley, J.C.; Pinson, S.R.M.; McClung, A.M.; Lales, J.S. Nitrogen utilization efficiency: Relationships with grain yield, grain protein, and yield-related traits in rice. Agron. J. 2006, 98, 168–176. [Google Scholar] [CrossRef] [Green Version]
- Gazulla, M.F.; Rodrigo, M.; Orduña, M.; Gómez, C.M. Determination of carbon, hydrogen, nitrogen and sulfur in geological materials using elemental analysers. Geostand. Geoanal. Res. 2012, 36, 201–217. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of Total Phenolics with Phosphomolybdic-Phosphotungstic Acid Reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Helmja, K.; Vaher, M.; Püssa, T.; Raudsepp, P.; Kaljurand, M. Evaluation of antioxidative capability of the tomato (Solanum lycopersicum) skin constituents by capillary electrophoresis and high-performance liquid chromatography. Electrophoresis 2008, 29, 3980–3988. [Google Scholar] [CrossRef] [PubMed]
- Plazas, M.; Prohens, J.; Cuñat, A.N.; Vilanova, S.; Gramazio, P.; Herraiz, F.J.; Andújar, I. Reducing capacity, chlorogenic acid content and biological activity in a collection of scarlet (Solanum aethiopicum) and Gboma (S. macrocarpon) eggplants. Int. J. Mol. Sci. 2014, 15, 17221–17241. [Google Scholar] [CrossRef] [Green Version]
- Little, T.M.; Hills, F.J. Agricultural Experimentation: Desing and Analysis; Wiley: Hoboken, NJ, USA, 1978; ISBN 0471023523. [Google Scholar]
- Wricke, G.; Weber, E. Quantitative Genetics and Selection in Plant Breeding; De Gruyter: Berlin, Germany, 1986; ISBN 978-3110075618. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing 2020; R Core Team: Vienna, Austria, 2020. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis 2016; Taylor & Francis: Abingdon, UK, 2016. [Google Scholar]
- Hochberg, Y. A sharper bonferroni procedure for multiple tests of significance. Biometrika 1988, 75, 800–802. [Google Scholar] [CrossRef]
- Revelle, W. psych: Procedures for Personality and Phychological Research 2017; Northwestern University: Evanston, IL, USA, 2017. [Google Scholar]
- Wei, T.; Simko, V. R Package “Corrplot”: Visualization of a Correlation Matrix 2017; R Core Team: Vienna, Austria, 2017. [Google Scholar]
- Broman, K.W.; Wu, H.; Sen, S.; Churchill, G.A. R/qtl: QTL mapping in experimental crosses. Bioinformatics 2003, 19, 889–890. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Pozo, N.; Menda, N.; Edwards, J.D.; Saha, P.; Tecle, I.Y.; Strickler, S.R.; Bombarely, A.; Fisher-York, T.; Pujar, A.; Foerster, H.; et al. The Sol Genomics Network (SGN): From genotype to phenotype to breeding. Nucleic Acids Res. 2015, 43, D1036–D1041. [Google Scholar] [CrossRef]
- Van Bueren, E.T.L.; Struik, P.C. Diverse concepts of breeding for nitrogen use efficiency. A review. Agron. Sustain. Dev. 2017, 37, 50. [Google Scholar] [CrossRef] [Green Version]
- Han, M.; Okamoto, M.; Beatty, P.H.; Rothstein, S.J.; Good, A.G. The genetics of nitrogen use efficiency in crop plants. Annu. Rev. Genet. 2015, 49, 269–289. [Google Scholar] [CrossRef]
- Kurai, T.; Wakayama, M.; Abiko, T.; Yanagisawa, S.; Aoki, N.; Ohsugi, R. Introduction of the ZmDof1 gene into rice enhances carbon and nitrogen assimilation under low-nitrogen conditions. Plant. Biotechnol. J. 2011, 9, 826–837. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Xia, X.; Zhang, Z.; Nong, B.; Zeng, Y. QTL mapping by whole genome re-sequencing and analysis of candidate genes for nitrogen use efficiency in rice. Front. Plant. Sci. 2017, 8, 1634. [Google Scholar] [CrossRef] [Green Version]
- Neeraja, C.N.; Voleti, S.R.; Subrahmanyam, D.; Surekha, K.; Rao, P.R. Breeding rice for nitrogen use efficiency. Indian Soc. Genet. Plant Breed. 2019, 79, 208–215. [Google Scholar] [CrossRef]
- Cormier, F.; Foulkers, J.; Hirel, B.; Gouache, D.; Moênne-Loccoz, Y.; Gouis, J. le Review Breeding for increased nitrogen-use efficiency: A review for wheat (T. aestivum L.). Plant Breed. 2016, 278, 255–278. [Google Scholar] [CrossRef] [Green Version]
- Lupini, A.; Preiti, G.; Badagliacca, G.; Abenavoli, M.R.; Sunseri, F.; Monti, M.; Bacchi, M. Nitrogen use efficiency in durum wheat under different nitrogen and water regimes in the mediterranean basin. Front. Plant Sci. 2021, 11, 607226. [Google Scholar] [CrossRef]
- Weber, V.S.; Melchinger, A.E.; Magorokosho, C.; Makumbi, D.; Bänziger, M.; Atlin, G.N. Efficiency of managed-stress screening of elite maize hybrids under drought and low nitrogen for yield under rainfed conditions in southern Africa. Crop. Sci. 2012, 52, 1011–1020. [Google Scholar] [CrossRef] [Green Version]
- Cai, H.; Chu, Q.; Yuan, L. Identification of quantitative trait loci for leaf area and chlorophyll content in maize (Zea mays) under low nitrogen and low phosphorus supply. Mol. Breed. 2012, 30, 251–266. [Google Scholar] [CrossRef]
- Semagn, K.; Beyene, Y.; Babu, R.; Nair, S.; Gowda, M.; Das, B.; Tarekegne, A.; Mugo, S.; Mahuku, G.; Worku, M.; et al. Quantitative trait loci mapping andmolecular breeding for developing stress resilient maize for sub-saharan Africa. Crop. Sci. 2015, 1449–1459. [Google Scholar] [CrossRef]
- Ertiro, B.T.; Labuschagne, M.; Olsen, M.; Das, B.; Prasanna, B.M.; Gowda, M. Genetic dissection of nitrogen use efficiency in tropical maize through genome-wide association and genomic prediction. Front. Plant. Sci. 2020, 11, 474. [Google Scholar] [CrossRef]
- Rajala, A.; Jalli, M.; Jauhiainen, L.; Hannukkala, A. One century of Nordic barley breeding: Nitrogen use efficiency, agronomic traits and genetic diversity. J. Agric. Sci. 2017, 155, 582–598. [Google Scholar] [CrossRef]
- Pujarula, V.; Pusuluri, M.; Bollam, S.; Das, R.R.; Ratnala, R.; Adapala, G.; Thuraga, V.; Rathore, A. Genetic variation for nitrogen use efficiency traits in global diversity panel and parents of mapping populations in pearl millet. Front. Plant Sci. 2021, 12, 625915. [Google Scholar] [CrossRef]
- Stahl, A.; Vollrath, P.; Samans, B.; Frisch, M.; Wittkop, B.; Snowdon, R.J. Effect of breeding on nitrogen use efficiency-associated traits in oilseed rape. J. Exp. Bot. 2019, 70, 1969–1986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiwari, J.K.; Plett, D.; Garnett, T.; Chakrabarti, S.K.; Singh, R.K. Integrated genomics, physiology and breeding approaches for improving nitrogen use efficiency in potato: Translating knowledge from other crops. Funct. Plant Biol. 2018, 45, 587–605. [Google Scholar] [CrossRef] [Green Version]
- Getahun, B.B. Potato breeding for nitrogen-use efficiency: Constraints, achievements, and future prospects. J. Crop Sci. Biotechnol. 2018, 21, 269–281. [Google Scholar] [CrossRef]
- Abenavoli, M.R.; Longo, C.; Lupini, A.; Miller, A.J.; Araniti, F.; Mercati, F.; Princi, M.P.; Sunseri, F. Phenotyping two tomato genotypes with different nitrogen use efficiency. Plant Physiol. Biochem. 2016, 107, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Lupini, A.; Princi, M.P.; Araniti, F.; Miller, A.J.; Sunseri, F.; Abenavoli, M.R. Physiological and molecular responses in tomato under different forms of N nutrition. J. Plant Physiol. 2017, 216, 17–25. [Google Scholar] [CrossRef]
- Liang, L.; Ridoutt, B.G.; Lal, R.; Wang, D.; Wu, W.; Peng, P.; Hang, S.; Wang, L.; Zhao, G. Nitrogen footprint and nitrogen use efficiency of greenhouse tomato production in North China. J. Clean. Prod. 2019, 208, 285–296. [Google Scholar] [CrossRef]
- Mauceri, A.; Bassolino, L.; Lupini, A.; Badeck, F.; Rizza, F.; Schiavi, M.; Toppino, L.; Abenavoli, M.R.; Rotino, G.L.; Sunseri, F. Genetic variation in eggplant for Nitrogen Use Efficiency under contrasting NO3- supply. J. Integr. Plant Biol. 2020, 62, 487–508. [Google Scholar] [CrossRef] [Green Version]
- Mauceri, A.; Rosa Abenavoli, M.; Toppino, L.; Panda, S.; Mercati, F.; Miyassa Aci, M.; Aharoni, A.; Sunseri, F.; Rotino, G.L.; Lupini, A. Transcriptomic insights on molecular regulation of Solanum melongena L. N-Use Efficiency. J. Exp. Bot. 2021, erab121. [Google Scholar] [CrossRef]
- Syfert, M.M.; Castañeda-Álvarez, N.P.; Khoury, C.K.; Särkinen, T.; Sosa, C.C.; Achicanoy, H.A.; Bernau, V.; Prohens, J.; Daunay, M.-C.; Knapp, S. Crop wild relatives of the brinjal eggplant (Solanum melongena): Poorly represented in genebanks and many species at risk of extinction. Am. J. Bot. 2016, 103, 635–651. [Google Scholar] [CrossRef] [Green Version]
- Baixauli, C.; Aguilar, J. Cultivo Sin Suelo de Hortalizas: Aspectos Prácticos y Experiencias; Generalitat Valenciana: Valencia, Spain, 2002; ISBN 8448231457. [Google Scholar]
- Diouf, I.A.; Derivot, L.; Bitton, F.; Pascual, L.; Causse, M. Water deficit and salinity stress reveal many specific QTL for plant growth and fruit quality traits in tomato. Front. Plant Sci. 2018, 9, 279. [Google Scholar] [CrossRef] [Green Version]
- Asins, M.J.; Raga, M.V.; Torrent, D.; Roca, D.; Carbonell, E.A. QTL and candidate gene analyses of rootstock-mediated tomato fruit yield and quality traits under low iron stress. Euphytica 2020, 216, 1–19. [Google Scholar] [CrossRef]
- Särkinen, T.; Bohs, L.; Olmstead, R.G.; Knapp, S. A phylogenetic framework for evolutionary study of the nightshades (Solanaceae): A dated 1000-tip tree. BMC Evol. Biol. 2013, 13, 214. [Google Scholar] [CrossRef] [Green Version]
- Prohens, J.; Plazas, M.; Raigón, M.D.; Simarro, J.M.S.; Stommel, J.R.; Vilanova, S. Characterization of interspecific hybrids and first backcross generations from crosses between two cultivated eggplants (Solanum melongena and S. aethiopicum Kumba group) and implications for eggplant breeding. Euphytica 2012, 186, 517–538. [Google Scholar] [CrossRef]
- Meyer, R.S.; Whitaker, B.D.; Little, D.P.; Wu, S.B.; Kennelly, E.J.; Long, C.L.; Litt, A. Parallel reductions in phenolic constituents resulting from the domestication of eggplant. Phytochemistry 2015, 115, 194–206. [Google Scholar] [CrossRef]
- Stommel, J.R.; Whitaker, B.D. Phenolic Acid Content and composition of eggplant fruit in a germplasm core subset. J. Am. Soc. Hort. Sci. 2003, 128, 704–710. [Google Scholar] [CrossRef]
- Mangino, G.; Plazas, M.; Vilanova, S.; Prohens, J.; Gramazio, P. Performance of a set of eggplant (Solanum melongena) lines with introgressions from its wild relative S. incanum under open field and screenhouse conditions and detection of QTLs. Agronomy 2020, 10, 467. [Google Scholar] [CrossRef] [Green Version]
- Frary, A.; Frary, A.; Daunay, M.-C.; Huvenaars, K.; Mank, R.; Doğanlar, S. QTL hotspots in eggplant (Solanum melongena) detected with a high resolution map and CIM analysis. Euphytica 2014, 197, 211–228. [Google Scholar] [CrossRef] [Green Version]
- Portis, E.; Cericola, F.; Barchi, L.; Toppino, L.; Acciarri, N.; Pulcini, L.; Sala, T.; Lanteri, S.; Rotino, G.L. Association mapping for fruit, plant and leaf morphology traits in eggplant. PLoS ONE 2015, 10, e0135200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plazas, M.; López-Gresa, M.P.; Vilanova, S.; Torres, C.; Hurtado, M.; Gramazio, P.; Andújar, I.; Herráiz, F.J.; Bellés, J.M.; Prohens, J. Diversity and relationships in key traits for functional and apparent quality in a collection of eggplant: Fruit phenolics content, antioxidant activity, polyphenol oxidase activity, and browning. J. Agric. Food Chem. 2013, 61, 8871–8879. [Google Scholar] [CrossRef] [PubMed]
- Hanson, P.M.; Yang, R.Y.; Tsou, S.C.S.; Ledesma, D.; Engle, L.; Lee, T.C. Diversity in eggplant (Solanum melongena) for superoxide scavenging activity, total phenolics, and ascorbic acid. J. Food Compos. Anal. 2006, 19, 594–600. [Google Scholar] [CrossRef]
- Chapman, M.A. (Ed.) The Eggplant Genome; Springer: Berlin/Heidelberg, Germany, 2019; ISBN 9783319992075. [Google Scholar]
- Doganlar, S.; Frary, A.; Daunay, M.-C.; Lester, R.N.; Tanksley, S.D. Conservation of gene function in the Solanaceae as revealed by comparative mapping of domestication traits in eggplant. Genetics 2002, 161, 1713–1726. [Google Scholar] [CrossRef]
- Gramazio, P.; Prohens, J.; Plazas, M.; Andjar, I.; Herraiz, F.J.; Castillo, E.; Knapp, S.; Meyer, R.S.; Vilanova, S. Location of chlorogenic acid biosynthesis pathway and polyphenol oxidase genes in a new interspecific anchored linkage map of eggplant. BMC Plant Biol. 2014, 14, 350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Portis, E.; Barchi, L.; Toppino, L.; Lanteri, S.; Acciarri, N.; Felicioni, N.; Fusari, F.; Barbierato, V.; Cericola, F.; Valè, G.; et al. QTL Mapping in Eggplant Reveals Clusters of Yield-Related Loci and Orthology with the Tomato Genome. PLoS ONE 2014, 9, e89499. [Google Scholar] [CrossRef] [Green Version]
- Miyatake, K.; Saito, T.; Nunome, T.; Yamaguchi, H.; Negoro, S.; Ohyama, A.; Wu, J.; Katayose, Y.; Fukuoka, H. Fine mapping of a major locus representing the lack of prickles in eggplant revealed the availability of a 0.5-kb insertion/deletion for marker-assisted selection. Breed. Sci. 2020, 70, 438–448. [Google Scholar] [CrossRef]
- Zhang, L.; Sun, H.; Xu, T.; Shi, T.; Li, Z.; Hou, W. Comparative transcriptome analysis reveals key genes and pathways involved in prickle development in eggplant. Genes 2021, 12, 341. [Google Scholar] [CrossRef] [PubMed]
- Mangino, G.; Vilanova, S.; Plazas, M.; Prohens, J.; Gramazio, P. Fruit shape morphometric analysis and QTL detection in a set of eggplant introgression lines. Sci. Hortic. 2021, 282, 110006. [Google Scholar] [CrossRef]
- Wei, Q.; Wang, W.; Hu, T.; Hu, H.; Wang, J.; Bao, C. Construction of a SNP-based genetic map using SLAF-Seq and QTL analysis of morphological traits in eggplant. Front. Genet. 2020, 11, 178. [Google Scholar] [CrossRef] [Green Version]
- Toppino, L.; Barchi, L.; Lo Scalzo, R.; Palazzolo, E.; Francese, G.; Fibiani, M.; D’Alessandro, A.; Papa, V.; Laudicina, V.A.; Sabatino, L.; et al. Mapping quantitative trait loci affecting biochemical and morphological fruit properties in eggplant (Solanum melongena L.). Front. Plant Sci. 2016, 7, 256. [Google Scholar] [CrossRef] [Green Version]
- Mouradov, A.; Spangenberg, G. Flavonoids: A metabolic network mediating plants adaptation to their real estate. Front. Plant Sci. 2014, 5, 620. [Google Scholar] [CrossRef]
- Günther, C.S.; Dare, A.P.; McGhie, T.K.; Deng, C.; Lafferty, D.J.; Plunkett, B.J.; Grierson, E.R.P.; Turner, J.L.; Jaakola, L.; Albert, N.W.; et al. Spatiotemporal modulation of flavonoid metabolism in blueberries. Front. Plant Sci. 2020, 11, 545. [Google Scholar] [CrossRef]
- Van der Rest, B.; Danoun, S.; Boudet, A.M.; Rochange, S.F. Down-regulation of cinnamoyl-CoA reductase in tomato (Solanum lycopersicum L.) induces dramatic changes in soluble phenolic pools. J. Exp. Bot. 2006, 57, 1399–1411. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Abbreviation | Units |
|---|---|---|
| Plant traits | ||
| SPAD | SPAD | - |
| Plant height | P-Height | cm |
| Aerial biomass | P-Biomass | kg FW a |
| Stem diameter | P-Diam | mm |
| Prickles in stem | P-StPrick | 0 (absence); 1 (presence) |
| Prickles in leaf | P-LeafPrick | 0 (absence); 1 (presence) |
| Yield | Yield | g plant−1 |
| Nitrogen Use Efficiency | NUE | - |
| Fruit traits | ||
| Fruit pedicel length | F-PedLength | mm |
| Fruit calyx length | F-CaLength | mm |
| Fruit length | F-Length | mm |
| Fruit width | F-Width | mm |
| Prickles in calyx | F-CalPrick | 0 (absence); 1 (presence) |
| Total number of fruits per plant | F-Number | - |
| Fruit mean weight | F-Weight | g |
| Composition traits | ||
| Nitrogen content in leaf | N-Leaf | g kg−1 DM b |
| Carbon content in leaf | C-Leaf | g kg−1 DM |
| Nitrogen content in fruit | N-Fruit | g kg−1 DM |
| Carbon content in fruit | C-Fruit | g kg−1 DM |
| Total phenolics content | TPC | g kg−1 FW |
| Chlorogenic acid content | CGA | g kg−1 FW |
| Total phenolic acid peaks area | TP-Area | units |
| Chlorogenic acid peak area | CGA-Area | % |
| Phenolic acids pattern | TP-Pattern | 0 (S. melongena pattern); 1 (S. elaeagnifolium pattern) |
| Trait | S. elaeagnifolium (n = 5) | S. melongena (n = 4) | BC2 (n = 5) | BC3 (n = 51) | H2 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | Range | CV (%) | Mean | Range | CV (%) | Mean | Range | CV (%) | Mean | Range | CV (%) | ||
| Plant traits | |||||||||||||
| SPAD | 63.7 b | 55.8–71.0 | 9.1 | 46.6 a | 44.0–47.9 | 3.9 | 44.2 a | 41.1–48.1 | 6.1 | 44.7 a | 37.0–56.9 | 9.0 | 0.79 |
| P-Height (cm) | 58.8 a | 50.0–65.0 | 9.3 | 92.3 b | 79.0–107.0 | 12.8 | 102.2 b | 90.0–130.0 | 16.3 | 85.4 b | 47.0–136.0 | 24.5 | 0.68 |
| P-Biomass (kg FW) | 0.27 a | 0.16–0.40 | 35.4 | 1.32 b | 0.85–2.03 | 38.2 | 1.59 b | 0.51–3.80 | 80.7 | 1.42 b | 0.18–4.96 | 76.0 | 0.78 |
| P-Diam (mm) | 10.5 a | 9.1–11.9 | 10.9 | 27.4 b | 19.3–35.4 | 25.4 | 26.3 b | 17.9–36.3 | 28.8 | 24.6 b | 11.0–46.2 | 30.9 | 0.15 |
| P-StPrick | 1.0 b | 1.0–1.0 | 0.0 | 0.0 a | 0–0 | 0.0 | 0.4 a | 0–1 | 136.9 | 0.2 a | 0–1 | 234.1 | 1.00 |
| P-LeafPrick | 0.0 | 0–0 | 0.0 | 0.0 | 0–0 | 0.0 | 0.2 | 0–1 | 223.6 | 0.2 | 0–1 | 234.1 | 1.00 |
| Yield (g) | 52.9 a | 12.0–114.0 | 83.6 | 2891 b | 1925–4020 | 32.5 | 1258 b | 469.0–2941.0 | 79.4 | 2059 b | 124.0–8109.0 | 87.1 | 0.71 |
| NUE | 10.3 a | 1.9–22.3 | 85.1 | 325.8 b | 205.6–483.2 | 37.1 | 200.5 b | 67.8–533.1 | 94.4 | 272.7 b | 37.4–1019.3 | 82.3 | 0.70 |
| Fruit traits | |||||||||||||
| F-PedLength (mm) | 21.3 a | 20.5–22.5 | 4.1 | 58.2 c | 52.6–61.5 | 6.8 | 28.4 ab | 20.1–35.7 | 22.8 | 38.4 b | 17.2–63.7 | 30.8 | 0.89 |
| F-CaLength (mm) | 12.2 a | 11.4–13.2 | 6.4 | 52.5 c | 49.2–56.7 | 5.9 | 29.8 b | 24.3–34.9 | 15.4 | 36.1 b | 20.2–52.6 | 23.8 | 0.87 |
| F-Length (mm) | 10.3 a | 9.1–11.3 | 9.2 | 94.8 c | 86.4–101.1 | 6.6 | 54.0 b | 38.1–71.9 | 23.0 | 66.1 b | 24.5–113.2 | 29.9 | 0.90 |
| F-Width (mm) | 10.1 a | 8.9–11.4 | 10.7 | 45.8 c | 41.8–53.9 | 12.0 | 29.9 b | 22.6–37.7 | 23.0 | 36.0 b | 18.9–50.5 | 19.6 | 0.42 |
| F-CalPrick | 1.0 b | 1.0–1.0 | 0.0 | 0.0 a | 0.0–0.0 | 0.0 | 0.4 a | 0–1 | 136.9 | 0.2 a | 0–1 | 218.2 | 1.00 |
| F-Number | 82.8 | 23.0–165.0 | 81.1 | 48.5 | 38.0–57.0 | 20.5 | 57.2 | 39.0–87.0 | 36.2 | 56.8 | 18.0–132.0 | 46.1 | 0.85 |
| F-Weight (g) | 0.61 a | 0.46–0.82 | 23.1 | 60.98 c | 33.77–77.79 | 31.6 | 20.03 b | 11.44–33.8 | 46.5 | 33.35 b | 5.29–65.88 | 47.5 | 0.57 |
| Composition traits | |||||||||||||
| N-Leaf (g/kg DM) | 45.0 a | 38.9–48.8 | 8.4 | 52.9 b | 49.4–54.7 | 4.5 | 53.1 b | 50.0–54.7 | 3.5 | 52.5 b | 45.0–58.2 | 5.3 | 0.24 |
| C-Leaf (g/kg DM) | 439.8 a | 435.0–446.0 | 1.0 | 443.3 a | 439.0–448.0 | 0.8 | 446.0 a | 437.0–453.0 | 1.6 | 455.5 b | 423.0–469.0 | 1.7 | 0.78 |
| N-Fruit (g/kg DM) | 26.4 | 24.2–30.3 | 9.2 | 24.6 | 22.9–25.5 | 4.9 | 29.3 | 24.6–33.9 | 13.2 | 25.7 | 19.2–40.9 | 15.5 | 0.91 |
| C-Fruit (g/kg DM) | 465.8 b | 461–472 | 1.0 | 428.3 a | 425–430 | 0.5 | 456.8 b | 438–477 | 3.1 | 435.7 a | 384–479 | 3.7 | 0.98 |
| TPC (g/kg FW) | 6.12 c | 4.1–7.44 | 20.9 | 1.99 a | 1.7–2.34 | 16.7 | 4.09 b | 3.62–5.0 | 13.4 | 2.76 a | 1.48–5.75 | 30.8 | 0.86 |
| CGA (g/kg FW) | 2.53 | 1.79–3.41 | 22.8 | 1.78 | 1.61–2.09 | 12.1 | 2.53 | 1.44–3.74 | 32.5 | 2.27 | 1.02–5.02 | 32.8 | 0.92 |
| TP-Area (units) | 32,931 b | 31,290–34,898 | 3.9 | 17,603 a | 15,639–20,419 | 12.1 | 22,874 a | 19,472–28,816 | 17.2 | 20,014 a | 11,517–32,806 | 24.2 | 0.80 |
| CGA-Area (%) | 31.4 a | 27.5–34.8 | 8.3 | 80.4 b | 76.7–81.5 | 1.0 | 67.8 b | 49.1–85.6 | 21.7 | 78.2 b | 49.1–88.4 | 11.7 | 0.99 |
| TP-Pattern | 1.0 c | 1.0–1.0 | 0.0 | 0.0 a | 0.0–0.0 | 0.0 | 0.6 bc | 0–1 | 91.3 | 0.2 ab | 0–1 | 192.6 | 1.00 |
| Trait | QTL | Chr. | Position (Mb.) | Heterozygous Allelic Effect (Units) | LOD Score |
|---|---|---|---|---|---|
| Plant traits | |||||
| Stem diameter (P-Diam) | pd4 | 4 | 21.09–68.11 | −8.9 (mm) | 5.82 |
| Prickles in stem (P-StPrick) | ps6 | 6 | 105.06–105.56 | 0.91 | 11.10 |
| Prickles in leaf (P-LeafPrick) | pl6 | 6 | 105.06–105.56 | 0.82 | 33.08 |
| Fruit traits | |||||
| Fruit width (F-Width) | fw7 | 7 | 0–0.52 | −9.5 (mm) | 6.12 |
| Prickles in fruit calyx (F-CalPrick) | pc6 | 6 | 105.06–105.56 | 0.89 | 447.96 |
| Composition traits | |||||
| Chlorogenic acid content (CGA) | cg5 | 5 | 3.94–4.49 | 2.26 (g kg−1 DM) | 7.15 |
| Total phenolic acid peaks area (TP-Area) | ph6 | 6 | 99.76–100.78 | 5105 | 5.20 |
| Chlorogenic acid peak area (CGA-Area) | ca1 | 1 | 1.05–1.42 | −20.1 (%) | 19.13 |
| Phenolic acids pattern (TP-Pattern) | cp1 | 1 | 1.05–1.42 | 0.88 | 416.14 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Villanueva, G.; Rosa-Martínez, E.; Şahin, A.; García-Fortea, E.; Plazas, M.; Prohens, J.; Vilanova, S. Evaluation of Advanced Backcrosses of Eggplant with Solanum elaeagnifolium Introgressions under Low N Conditions. Agronomy 2021, 11, 1770. https://doi.org/10.3390/agronomy11091770
Villanueva G, Rosa-Martínez E, Şahin A, García-Fortea E, Plazas M, Prohens J, Vilanova S. Evaluation of Advanced Backcrosses of Eggplant with Solanum elaeagnifolium Introgressions under Low N Conditions. Agronomy. 2021; 11(9):1770. https://doi.org/10.3390/agronomy11091770
Chicago/Turabian StyleVillanueva, Gloria, Elena Rosa-Martínez, Ahmet Şahin, Edgar García-Fortea, Mariola Plazas, Jaime Prohens, and Santiago Vilanova. 2021. "Evaluation of Advanced Backcrosses of Eggplant with Solanum elaeagnifolium Introgressions under Low N Conditions" Agronomy 11, no. 9: 1770. https://doi.org/10.3390/agronomy11091770