
Microbial Biomass Sulphur—An Important Yet Understudied Pool in Soil

 , , , and
, , , and
Abstract
:1. Introduction
2. Data Acquisition, Handling, and Statistical Analysis
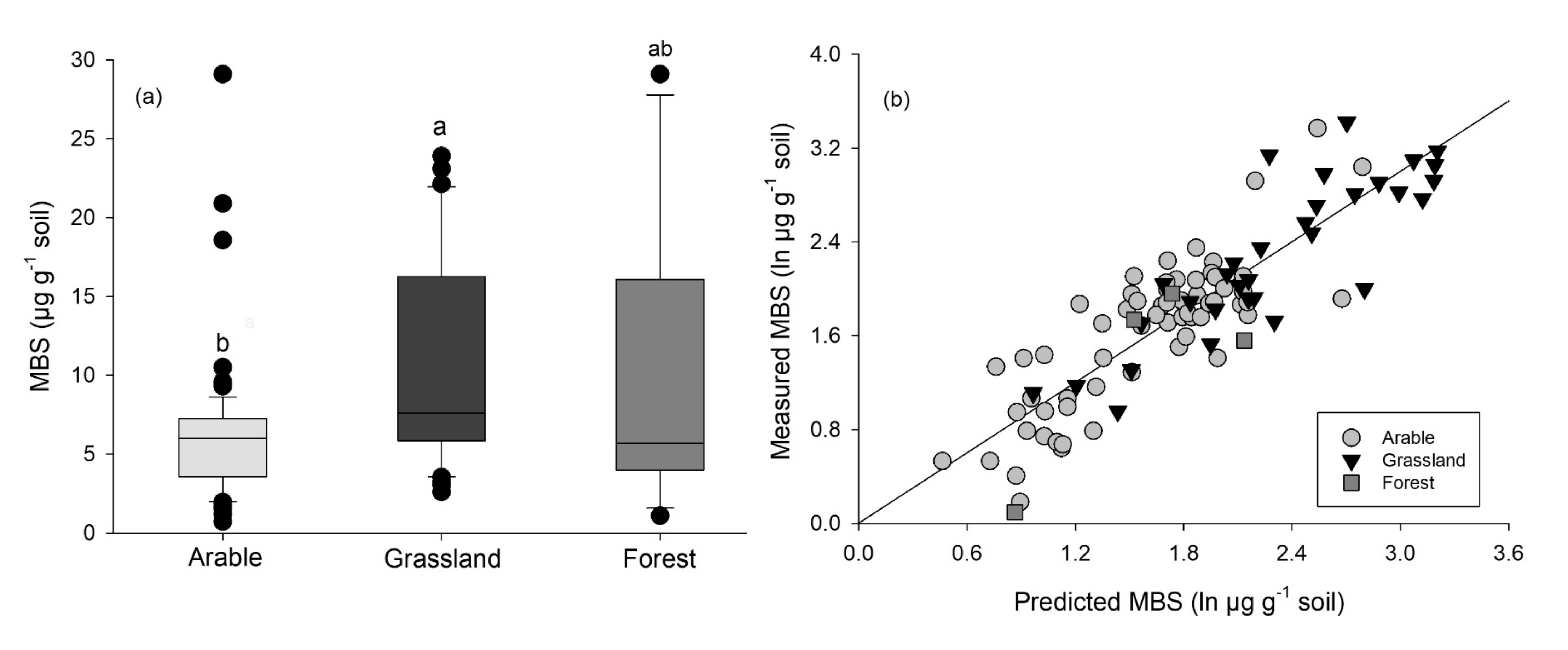
3. Land-Use Effects on MBS
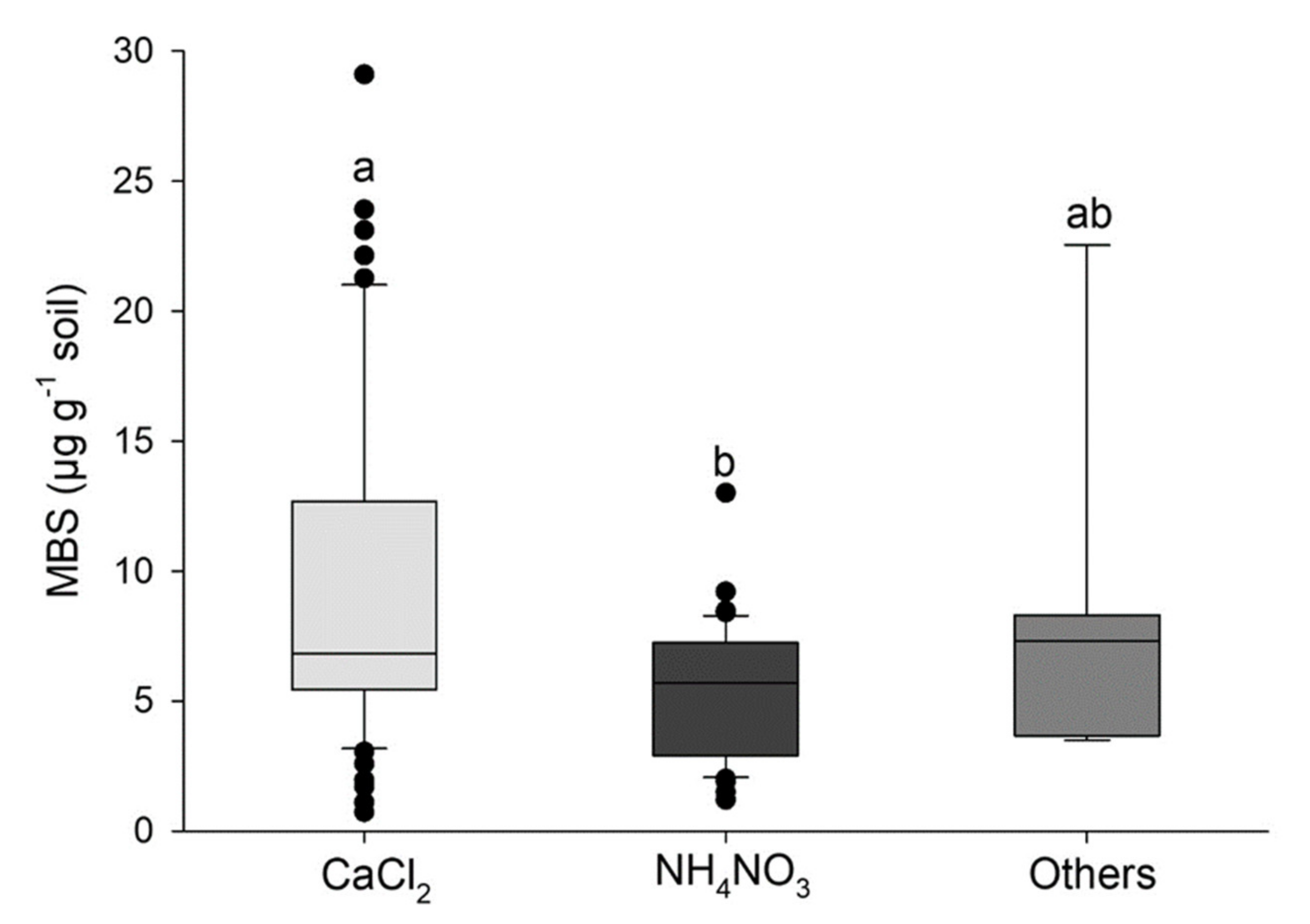
4. Methodological Remarks on MBS Determination
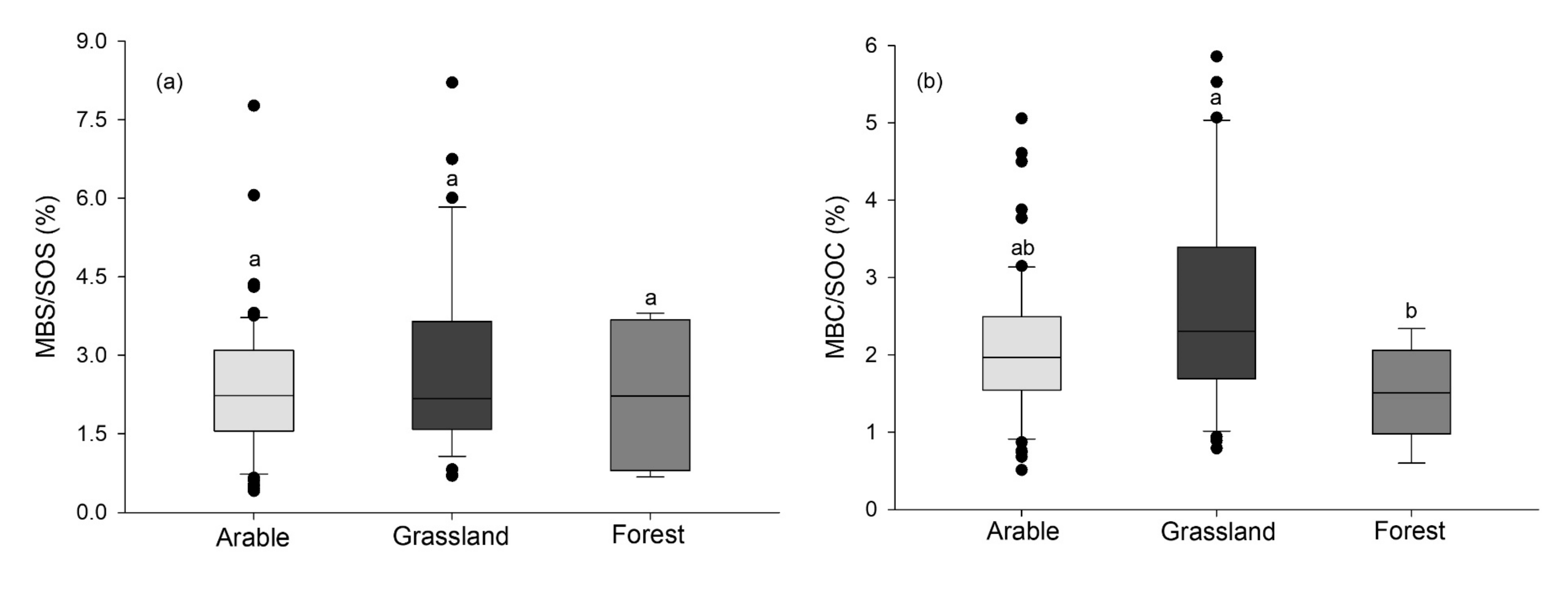
5. MBS Stoichiometry
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Muhammad, S.; Müller, T.; Joergensen, R.G. Relationships between soil biological and other soil properties in saline and alkaline arable soils from the Pakistani Punjab. J. Arid Environ. 2008, 72, 448–457. [Google Scholar] [CrossRef]
- Khan, K.S.; Mack, R.; Castillo, X.; Kaiser, M.; Joergensen, R.G. Microbial biomass, fungal and bacterial residues, and their relationships to the soil organic matter C/N/P/S ratios. Geoderma 2016, 271, 115–123. [Google Scholar] [CrossRef]
- Johnson, D.C.; Dean, D.R.; Smith, A.D.; Johnson, M.K. Structure, function, and formation of biological iron-sulfur clusters. Ann. Rev. Biochem. 2005, 74, 247–281. [Google Scholar] [CrossRef]
- Rimmer, D.L. Free radicals, antioxidants, and soil organic matter recalcitrance. Eur. J. Soil Sci. 2006, 57, 91–94. [Google Scholar] [CrossRef]
- Schlichting, A.; Rimmer, D.L.; Eckhardt, K.-U.; Heumann, S.; Abbott, G.D.; Leinweber, P. Identifying potential antioxidant compounds in NaOH extracts of UK soils and vegetation by untargeted mass spectrometric screening. Soil Biol. Biochem. 2013, 58, 16–26. [Google Scholar] [CrossRef]
- Kirkby, C.A.; Richardson, A.E.; Wade, L.J.; Batten, G.D.; Blanchard, C.; Kirkegaard, J.A. Carbon-nutrient stoichiometry to increase soil carbon sequestration. Soil Biol. Biochem. 2013, 60, 77–86. [Google Scholar] [CrossRef]
- Tipping, E.; Somerville, C.J.; Luster, J. The C:N:P:S stoichiometry of soil organic matter. Biogeochemistry 2016, 130, 117–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapman, S.J. Microbial sulphur in some Scottish soils. Soil Biol. Biochem. 1987, 19, 301–305. [Google Scholar] [CrossRef]
- Heinze, S.; Raupp, J.; Joergensen, R.G. Effects of fertilizer and spatial heterogeneity in soil pH on microbial biomass indices in a long-term field trial of organic agriculture. Plant Soil 2010, 328, 203–215. [Google Scholar] [CrossRef]
- Heinze, S.; Rauber, R.; Joergensen, R.G. Influence of mouldboard plough and rotary harrow tillage on microbial biomass and nutrient stocks in two long-term experiments on loess derived Luvisols. Appl. Soil Ecol. 2010, 46, 405–412. [Google Scholar] [CrossRef]
- Banerjee, M.R.; Chapman, S.J. The significance of microbial biomass sulphur in soil. Biol. Fertil. Soils 1996, 22, 116–125. [Google Scholar] [CrossRef]
- Aliasgharzadeh, N.; Saedi, S.; Zamzami, S. Efficiency of acidophilic Thiobacillus in sulfur oxidation and pH reducing in soil. J. Agric. Sci. 1998, 8, 75–91. [Google Scholar]
- Dedourge, O.; Vong, P.-C.; Lasserre-Joulin, F.; Benizri, E.; Guckert, A. Immobilization of sulphur-35, microbial biomass and arylsulphatase activity in soils from field-grown rape, barley and fallow. Biol. Fertil. Soils 2003, 38, 181–185. [Google Scholar] [CrossRef]
- Kertesz, M.A.; Mirleau, P. The role of soil microbes in plant sulphur nutrition. J. Exp. Bot. 2004, 55, 1939–1945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jannoura, R.; Bruns, C.; Joergensen, R.G. Organic fertilizer effects on pea yield, nutrient uptake, microbial root colonization and soil microbial biomass indices in organic farming systems. Eur. J. Agron. 2013, 49, 32–41. [Google Scholar] [CrossRef]
- Nowlan, C.R.; Martin, R.V.; Philip, S.; Lamsal, L.N.; Krotkov, N.A.; Marais, E.A.; Wang, S.; Zhang, Q. Global dry deposition of nitrogen dioxide and sulfur dioxide inferred from space-based measurements. Glob. Biogeochem. Cycles 2014, 28, 1025–1043. [Google Scholar] [CrossRef] [Green Version]
- Aas, W.; Mortier, A.; van Bowersox, V.; Cherian, R.; Faluvegi, G.; Fagerli, H.; Hand, J.; Klimont, Z.; Galy-Lacaux, C.; Lehmann, C.M.B.; et al. Global and regional trends of atmospheric sulfur. Sci. Rep. 2019, 9, 953. [Google Scholar] [CrossRef]
- Saggar, S.; Bettany, J.R.; Stewart, J. Measurement of microbial sulfur in soil. Soil Biol. Biochem. 1981, 13, 493–498. [Google Scholar] [CrossRef]
- Strick, J.E.; Nakas, J.P. Calibration of a microbial sulfur technique for use in forest soils. Soil Biol. Biochem. 1984, 16, 289–291. [Google Scholar] [CrossRef]
- Ghani, A.; McLaren, R.; Swift, R.G. The incorporation and transformations of 35S in soil: Effects of soil conditioning and glucose or sulphate additions. Soil Biol. Biochem. 1993, 25, 327–335. [Google Scholar] [CrossRef]
- Vong, P.-C.; Dedourge, O.; Lasserre-Joulin, F.; Guckert, A. Immobilized-S, microbial biomass-S and soil arylsulfatase activity in the rhizosphere soil of rape and barley as affected by labile substrate C and N additions. Soil Biol. Biochem. 2003, 35, 1651–1661. [Google Scholar] [CrossRef]
- Chalk, P.M.; Inácio, C.T.; Chen, D. Tracing S dynamics in agro-ecosystems using 34S. Soil Biol. Biochem. 2017, 114, 295–308. [Google Scholar] [CrossRef]
- Wirth, J.S.; Wang, T.; Huang, Q.; White, R.H.; Whitman, W.B. Dimethylsulfoniopropionate sulfur and methyl carbon assimilation in Ruegeria species. mBio 2020, 11, e00329-20. [Google Scholar] [CrossRef] [Green Version]
- Tcherkez, G.; Tea, I. 32S/34S isotope fractionation in plant sulphur metabolism. New Phytol. 2013, 200, 44–53. [Google Scholar] [CrossRef]
- Cleveland, C.C.; Liptzin, D. C:N:P stoichiometry in soil: Is there a “Redfield ratio” for the microbial biomass? Biogeochemistry 2007, 85, 235–252. [Google Scholar] [CrossRef]
- Manzoni, S.; Trofymow, J.A.; Jackson, R.B.; Porporato, A. Stoichiometric controls on carbon, nitrogen, and phosphorus dynamics in decomposing litter. Ecol. Monogr. 2010, 80, 89–106. [Google Scholar] [CrossRef]
- Zechmeister-Boltenstern, S.; Keiblinger, K.M.; Mooshammer, M.; Peñuelas, J.; Richter, A.; Sardans, J.; Wanek, W. The application of ecological stoichiometry to plant–microbial–soil organic matter transformations. Ecol. Monogr. 2015, 85, 133–155. [Google Scholar] [CrossRef] [Green Version]
- Hartman, W.H.; Richardson, C.J. Differential nutrient limitation of soil microbial biomass and metabolic quotients (qCO2): Is there a biological stoichiometry of soil microbes? PLoS ONE 2013, 8, e57127. [Google Scholar] [CrossRef]
- Chen, J.; Seven, J.; Zilla, T.; Dippold, M.A.; Blagodatskaya, E.; Kuzyakov, Y. Microbial C:N:P stoichiometry and turnover depend on nutrients availability in soil: A 14C, 15N and 33P triple labelling study. Soil Biol. Biochem. 2019, 131, 206–216. [Google Scholar] [CrossRef]
- Coonan, E.C.; Kirkby, C.A.; Kirkegaard, J.A.; Amidy, M.R.; Strong, C.L.; Richardson, A.E. Microorganisms and nutrient stoichiometry as mediators of soil organic matter dynamics. Nutr. Cycl. Agroecosyst. 2020, 117, 273–298. [Google Scholar] [CrossRef]
- Khan, K.S.; Joergensen, R.G. Stoichiometry of the soil microbial biomass in response to amendments with varying C/N/P/S ratios. Biol. Fertil. Soils 2019, 55, 265–274. [Google Scholar] [CrossRef]
- Khan, K.S.; Heinze, S.; Joergensen, R.G. Simultaneous measurement of S, macronutrients, and heavy metals in the soil microbial biomass with CHCl3 fumigation and NH4NO3 extraction. Soil Biol. Biochem. 2009, 41, 309–314. [Google Scholar] [CrossRef]
- Ahern, C.R.; Baker, D.E.; Aitken, R.L. Models for relating pH measurements in water and calcium chloride for a wide range of pH, soil types and depths. Plant Soil 1995, 171, 47–52. [Google Scholar] [CrossRef]
- Doerffel, K. Statistik in der Analytischen Chemie, 3rd ed.; Verlag Chemie: Weinheim, Germany, 1984. [Google Scholar]
- Wu, J.; O’Donnell, A.G.; He, Z.L.; Syers, J.K. Fumigation-extraction method for the measurement of soil microbial biomass-S. Soil Biol. Biochem. 1994, 26, 117–125. [Google Scholar] [CrossRef]
- Haynes, R.J. The use of polyethylene mulches to change soil microclimate as revealed by enzyme activity and biomass nitrogen, sulphur and phosphorus. Biol. Fertil. Soils 1987, 5, 235–240. [Google Scholar] [CrossRef]
- Haynes, R.J.; Swift, R.S. Effects of lime and phosphate additions on changes in enzyme activities, microbial biomass and levels of extractable nitrogen, sulphur and phosphorus in an acid soil. Biol. Fertil. Soils 1988, 6, 153–158. [Google Scholar] [CrossRef]
- Sarathchandra, S.U.; Perrott, K.W.; Littler, R.A. Soil microbial biomass: Influence of simulated temperature changes on size, activity and nutrient-content. Soil Biol. Biochem. 1989, 21, 987–993. [Google Scholar] [CrossRef]
- Perrott, K.W.; Sarathchandra, S.U. Seasonal variations in soil S flush and possible contributions from plant roots in the measurement of soil microbial sulfur, phosphorus, potassium and nitrogen. Soil Res. 1990, 28, 747. [Google Scholar] [CrossRef]
- Anderson, T.-H.; Domsch, K.H. Ratios of microbial biomass carbon to total organic carbon in arable soils. Soil Biol. Biochem. 1989, 21, 471–479. [Google Scholar] [CrossRef]
- Anderson, T.-H.; Domsch, K.H. Soil microbial biomass: The eco-physiological approach. Soil Biol. Biochem. 2010, 42, 2039–2043. [Google Scholar] [CrossRef]
- Chowdhury, M.H.; Kouno, K.; Ando, T.; Nagaoka, T. Microbial biomass, S mineralization and S uptake by African millet from soil amended with various composts. Soil Biol. Biochem. 2000, 32, 845–852. [Google Scholar] [CrossRef]
- Merchant, S.S.; Helmann, J.D. Elemental economy: Microbial strategies for optimizing growth in the face of nutrient limitation. Adv. Microb. Physiol. 2012, 60, 91–210. [Google Scholar] [CrossRef] [Green Version]
- Godchaux, W.; Leadbetter, E.R. Sulfonolipids of gliding bacteria. Structure of the N-acylaminosulfonates. J. Biol. Chem. 1984, 259, 2982–2990. [Google Scholar] [CrossRef]
- Benning, C. Biosynthesis and function of the sulfolipid sulfoquinovosyl diacylglycerol. Ann. Rev. Plant Physiol. Plant Mol. Biol. 1998, 49, 53–75. [Google Scholar] [CrossRef] [PubMed]
- Brake, M.; Höper, H.; Joergensen, R.G. Land use-induced changes in activity and biomass of microorganisms in raised bog peats at different depths. Soil Biol. Biochem. 1999, 31, 1489–1497. [Google Scholar] [CrossRef]
- Zederer, D.P.; Talkner, U.; Spohn, M.; Joergensen, R.G. Microbial biomass phosphorus and C/N/P stoichiometry in forest floor and a horizons as affected by tree species. Soil Biol. Biochem. 2017, 111, 166–175. [Google Scholar] [CrossRef]
- Rosinger, C.; Rousk, J.; Sandén, H. Can enzymatic stoichiometry be used to determine growth-limiting nutrients for microorganisms?—A critical assessment in two subtropical soils. Soil Biol. Biochem. 2019, 128, 115–126. [Google Scholar] [CrossRef]
- Brookes, P.C.; Powlson, D.S.; Jenkinson, D.S. Measurement of microbial biomass phosphorus in soil. Soil Biol. Biochem. 1982, 14, 319–329. [Google Scholar] [CrossRef]
- Brookes, P.C.; Landman, A.; Pruden, G.; Jenkinson, D.S. Chloroform fumigation and the release of soil nitrogen: A rapid direct extraction method to measure microbial biomass nitrogen in soil. Soil Biol. Biochem. 1985, 17, 837–842. [Google Scholar] [CrossRef]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Saggar, S.; Bettany, J.R.; Stewart, J. Sulfur transformations in relation to carbon and nitrogen in incubated soils. Soil Biol. Biochem. 1981, 13, 499–511. [Google Scholar] [CrossRef]
- Banerjee, M.R.; Chapman, S.J.; Killham, K. Factors influencing the determination of microbial biomass sulphur in soil. Commun. Soil Sci. Plant Anal. 1993, 24, 939–950. [Google Scholar] [CrossRef]
- Chowdhury, M.A.H.; Kouno, K.; Ando, T. Correlation among microbial biomass s, soil properties, and other biomass nutrients. Soil Sci. Plant Nutr. 1999, 45, 175–186. [Google Scholar] [CrossRef]
- Chapman, S. Partitioning of ryegrass residue sulphur between the soil microbial biomass, other soil sulphur pools and ryegrass (Lolium perenne L.). Biol. Fertil. Soils 1987, 5. [Google Scholar] [CrossRef]
- Perucci, P. Effect of the addition of municipal solid-waste compost on microbial biomass and enzyme activities in soil. Biol. Fertil. Soils 1990, 10, 221–226. [Google Scholar] [CrossRef]
- Castellano, S.D.; Dick, R.P. Modified calibration procedure for the measurement of microbial sulfur in soil. Soil Sci. Soc. Am. J. 1991, 55, 283–285. [Google Scholar] [CrossRef]
- Castellano, S.D.; Dick, R.P. Cropping and sulfur fertilization influence on sulfur transformations in soil. Soil Sci. Soc. Am. J. 1991, 55, 114–121. [Google Scholar] [CrossRef]
- Ghani, A.; McLaren, R.; Swift, R.S. The Incorporation and Remineralisation of 35S in Soil Organic Sulphur Fractions. In Proceedings of the Towards the More Efficient use of Soil and Fertiliser Sulphur, Massey University, Palmerston North, NZ, USA, 17–18 February 1988. [Google Scholar]
- Ghani, A.; McLaren, R.G.; Swift, R.S. Seasonal fluctuations of sulphate and soil microbial biomass-S in the surface of a Wakanui soil. N. Z. J. Agric. Res. 1990, 33, 467–472. [Google Scholar] [CrossRef]
- Yavitt, J.B.; Wieder, R.K.; Wright, S.J. Soil nutrient dynamics in response to irrigation of a Panamanian tropical moist forest. Biogeochemistry 1992, 19, 1–25. [Google Scholar] [CrossRef]
- Prietzel, J.; Weick, C.; Korintenberg, J.; Seybold, G.; Thumerer, T.; Treml, B. Effects of repeated (NH4)2SO4 application on sulfur pools in soil, soil microbial biomass, and ground vegetation of two watersheds in the Black Forest/Germany. Plant Soil 2001, 230, 287–305. [Google Scholar] [CrossRef]
- Murugan, R.; Koch, H.-J.; Joergensen, R.G. Long-term influence of different tillage intensities on soil microbial biomass, residues and community structure at different depths. Biol. Fertil. Soils 2014, 50, 487–498. [Google Scholar] [CrossRef]
- Murugan, R.; Loges, R.; Taube, F.; Sradnick, A.; Joergensen, R.G. Changes in soil microbial biomass and residual indices as ecological indicators of land use change in temperate permanent grassland. Microb. Ecol. 2014, 67, 907–918. [Google Scholar] [CrossRef]
- Heinze, S.; Oltmanns, M.; Joergensen, R.G.; Raupp, J. Changes in microbial biomass indices after 10 years of farmyard manure and vegetal fertilizer application to a sandy soil under organic management. Plant Soil 2011, 343, 221–234. [Google Scholar] [CrossRef]
- Murugan, R.; Loges, R.; Taube, F.; Joergensen, R.G. Specific response of fungal and bacterial residues to one-season tillage and repeated slurry application in a permanent grassland soil. Appl. Soil Ecol. 2013, 72, 31–40. [Google Scholar] [CrossRef]
- Wu, J.; O’Donnell, A.G.; Syers, J.K. Microbial growth and sulphur immobilization following the incorporation of plant residues into soil. Soil Biol. Biochem. 1993, 25, 1567–1573. [Google Scholar] [CrossRef]
- He, Z.L.; Wu, J.; O’Donnell, A.G.; Syers, J.K. Seasonal responses in microbial biomass carbon, phosphorus and sulphur in soils under pasture. Biol. Fertil. Soils 1997, 24, 421–428. [Google Scholar] [CrossRef]
- Johnson, C.M.; Nishita, H. Microestimation of sulfur in plant materials, soils, and irrigation waters. Anal. Chem. 1952, 24, 736–742. [Google Scholar] [CrossRef]
- Ghani, A.; McLarren, R.G.; Swift, R.S. Mobilization of recently-formed soil organic sulphur. Soil Biol. Biochem. 1993, 25, 1739–1744. [Google Scholar] [CrossRef]
- Nguyen, M.L.; Goh, K.M. Accumulation of soil sulphur fractions in grazed pastures receiving long-term superphosphate applications. N. Z. J. Agric. Res. 1990, 33, 111–128. [Google Scholar] [CrossRef]
- Maynard, D.G.; Stewart, J.; Bettany, J.R. Sulfur and nitrogen mineralization in soils compared using two incubation techniques. Soil Biol. Biochem. 1983, 15, 251–256. [Google Scholar] [CrossRef]
- Gupta, V.V.S.R.; Lawrence, J.R.; Germida, J.J. Impact of elemental sulfur fertilization on agricultural soils. I. Effects on microbial biomass and enzyme activities. Can. J. Soil. Sci. 1988, 68, 463–473. [Google Scholar] [CrossRef]
- Gupta, V.V.S.R.; Germida, J.J. Microbial biomass and extractable sulfate sulfur levels in native and cultivated soils as influenced by air-drying and rewetting. Can. J. Soil. Sci. 1989, 69, 889–894. [Google Scholar] [CrossRef]
- Malik, K.M.; Khan, K.S.; Akhtar, M.S.; Ahmed, Z.I. Sulfur distribution and availability in alkaline subtropical soils affected by organic amendments. J. Soil Sci. Plant Nutr. 2020, 20, 2253–2266. [Google Scholar] [CrossRef]
- Chen, C.R.; Condron, L.M.; Davis, M.R.; Sherlock, R.R. Effects of land-use change from grassland to forest on soil sulfur and arylsulfatase activity in New Zealand. Soil Res. 2001, 39, 749–757. [Google Scholar] [CrossRef]
- Yang, Z.; Singh, B.R.; Hansen, S.; Hu, Z.; Riley, H. Aggregate associated sulfur fractions in long-term (>80 years) fertilized soils. Soil Sci. Soc. Am. J. 2007, 71, 163–170. [Google Scholar] [CrossRef]
- Joergensen, R.G. Quantification of the microbial biomass by determining ninhydrin-reactive N. Soil Biol. Biochem. 1996, 28, 301–306. [Google Scholar] [CrossRef]
- Martens, R. Current methods for measuring microbial biomass C in soil: Potentials and limitations. Biol. Fertil. Soils 1995, 19, 87–99. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, W.; Chen, W.; Zhang, N.; Zeng, H. Comparison of seasonal soil microbial process in snow-covered temperate ecosystems of northern China. PLoS ONE 2014, 9, e92985. [Google Scholar] [CrossRef] [Green Version]
- Joergensen, R.G.; Wichern, F. Alive and kicking: Why dormant soil microorganisms matter. Soil Biol. Biochem. 2018, 116, 419–430. [Google Scholar] [CrossRef]
- Randlett, D.L.; Zak, D.R.; MacDonald, N.W. Sulfate adsorption and microbial immobilization in northern hardwood forests along an atmospheric deposition gradient. Can. J. For. Res. 1992, 22, 1843–1850. [Google Scholar] [CrossRef]
- Needelman, B.A.; Wander, M.M.; Shi, G.S. Organic carbon extraction efficiency in chloroform fumigated and non-fumigated soils. Soil Sci. Soc. Am. J. 2001, 65, 1731–1733. [Google Scholar] [CrossRef]
- Wu, J.; Joergensen, R.G.; Pommerening, B.; Chaussod, R.; Brookes, P.C. Measurement of soil microbial biomass C by fumigation-extraction—An automated procedure. Soil Biol. Biochem. 1990, 22, 1167–1169. [Google Scholar] [CrossRef]
- Joergensen, R.G.; Mueller, T. The fumigation-extraction method to estimate soil microbial biomass: Calibration of the kEN value. Soil Biol. Biochem. 1996, 28, 33–37. [Google Scholar] [CrossRef]
- Greenfield, L.G. Release of microbial cell N during chloroform fumigation. Soil Biol. Biochem. 1995, 27, 1235–1236. [Google Scholar] [CrossRef]
- Eberhardt, U.; Apel, G.; Joergensen, R.G. Effects of direct chloroform fumigation on suspended cells of 14C and 32P labelled bacteria and fungi. Soil Biol. Biochem. 1996, 28, 677–679. [Google Scholar] [CrossRef]
- Joergensen, R.G.; Wichern, F. Quantitative assessment of the fungal contribution to microbial tissue in soil. Soil Biol. Biochem. 2008, 40, 2977–2991. [Google Scholar] [CrossRef]
- Jenkinson, D.S.; Powlson, D.S.; Wedderburn, R.W.M. The effects of biocidal treatments on metabolism in soil—III. The relationship between soil biovolume, measured by optical microscopy, and the flush of decomposition caused by fumigation. Soil Biol. Biochem. 1976, 8, 189–202. [Google Scholar] [CrossRef]
- Tate, K.R.; Ross, D.J.; Feltham, C.W. A direct extraction method to estimate soil microbial c: Effects of experimental variables and some different calibration procedures. Soil Biol. Biochem. 1988, 20, 329–335. [Google Scholar] [CrossRef]
- Joergensen, R.G. Organic matter and micro-organisms in tropical soils. In Soil Biology and Agriculture in the Tropics; Dion, P., Ed.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 17–44. [Google Scholar]
- Salamanca, E.F.; Raubuch, M.; Joergensen, R.G. Microbial reaction of secondary tropical forest soils to the addition of leaf litter. Appl. Soil Ecol. 2006, 31, 53–61. [Google Scholar] [CrossRef]
- Jenkinson, D.S. Soil organic matter and its dynamics. In Russell’s Soil Conditions and Plant Growth, 11th ed.; Wild, A., Ed.; Longman: Essex, UK, 1988; pp. 565–607. [Google Scholar]
- Nguyen, M.L.; Goh, K.M. Sulphur cycling and its implications on sulphur fertilizer requirements of grazed grassland ecosystems. Agric. Ecosyst. Environ. 1994, 49, 173–206. [Google Scholar] [CrossRef]
- Guillén-Navarro, K.; Encarnación, S.; Dunn, M.F. Biotin biosynthesis, transport and utilization in rhizobia. FEMS Microbiol. Lett. 2005, 246, 159–165. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, G.; Mendel, R.R. Molybdenum cofactor biosynthesis and molybdenum enzymes. Ann. Rev. Plant Biol. 2006, 57, 623–647. [Google Scholar] [CrossRef]
- Spalding, M.D.; Prigge, S.T. Lipoic acid metabolism in microbial pathogens. Microbiol. Mol. Biol. Rev. 2010, 74, 200–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xavier, J.C.; Patil, K.R.; Rocha, I. Integration of biomass formulations of genome-scale metabolic models with experimental data reveals universally essential cofactors in prokaryotes. Metab. Eng. 2017, 39, 200–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pócsi, I.; Prade, R.A.; Penninckx, M.J. Glutathione, altruistic metabolite in fungi. Adv. Microbial. Physiol. 2004, 49, 1–76. [Google Scholar] [CrossRef]
- Smirnova, G.V.; Oktyabrsky, O.N. Glutathione in bacteria. Biochemistry 2005, 70, 1199–1211. [Google Scholar] [CrossRef]
- Schiraldi, C.; Cimini, D.; de Rosa, M. Production of chondroitin sulfate and chondroitin. Appl. Microbiol. Biotechnol. 2010, 87, 1209–1220. [Google Scholar] [CrossRef] [PubMed]
- Joergensen, R.G. Amino sugars as specific indices for fungal and bacterial residues in soil. Biol. Fertil. Soils 2018, 54, 559–568. [Google Scholar] [CrossRef]
- Fitzgerald, J.W.; Strickland, T.C.; Swank, W.T. Metabolic fate of inorganic sulphate in soil samples from undisturbed and managed forest ecosystems. Soil Biol. Biochem. 1982, 14, 529–536. [Google Scholar] [CrossRef]
- Linder, T. Assimilation of alternative sulfur sources in fungi. World J. Microbiol. Biotechnol. 2018, 34, 51. [Google Scholar] [CrossRef] [Green Version]
- Klionsky, D.J.; Herman, P.K.; Emr, S.D. The fungal vacuole: Composition, function, and biogenesis. Microbiol. Rev. 1990, 54, 266–292. [Google Scholar] [CrossRef] [PubMed]
- Engelking, B.; Flessa, H.; Joergensen, R.G. Shifts in amino sugar and ergosterol contents after addition of sucrose and cellulose to soil. Soil Biol. Biochem. 2007, 39, 2111–2118. [Google Scholar] [CrossRef]
- Baldrian, P.; Valásková, V. Degradation of cellulose by basidiomycetous fungi. FEMS Microbiol. Rev. 2008, 32, 501–521. [Google Scholar] [CrossRef] [Green Version]
- Chowdhury, M.A.H.; Kouno, K.; Ando, T. Critical sulphur concentration and sulphur requirement of microbial biomass in a glucose and cellulose-amended regosol. Biol. Fertil. Soils 2000, 32, 310–317. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Arable Soils | Grassland Soils | Forest Soils | |
|---|---|---|---|---|
| Soil pH-H2O | Median | 6.7 | 5.9 | 3.9 |
| 25% | 6.0 | 5.4 | 3.8 | |
| 75% | 7.5 | 6.6 | 4.7 | |
| n | 68 | 37 | 7 | |
| SOC (mg g−1 soil) | Median | 13.0 | 35.4 | 36.1 |
| 25% | 8.6 | 23.8 | 20.3 | |
| 75% | 22.8 | 45.4 | 53.2 | |
| n | 65 | 41 | 8 | |
| Total N (mg g−1 soil) | Median | 1.24 | 2.41 | 1.55 |
| 25% | 0.81 | 1.60 | 1.22 | |
| 75% | 1.82 | 3.44 | 1.83 | |
| n | 64 | 27 | 4 | |
| SOS (µg g−1 soil) | Median | 224 | 385 | 189 |
| 25% | 172 | 257 | 152 | |
| 75% | 362 | 654 | 360 | |
| n | 72 | 39 | 4 | |
| MBC (µg g−1 soil) | Median | 274 | 755 | 776 |
| 25% | 151 | 442 | 462 | |
| 75% | 402 | 1709 | 1122 | |
| n | 67 | 35 | 7 | |
| MBN (µg g−1 soil) | Median | 35 | 95 | 107 |
| 25% | 20 | 57 | 87 | |
| 75% | 49 | 138 | 259 | |
| n | 47 | 24 | 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heinze, S.; Hemkemeyer, M.; Schwalb, S.A.; Khan, K.S.; Joergensen, R.G.; Wichern, F. Microbial Biomass Sulphur—An Important Yet Understudied Pool in Soil. Agronomy 2021, 11, 1606. https://doi.org/10.3390/agronomy11081606
Heinze S, Hemkemeyer M, Schwalb SA, Khan KS, Joergensen RG, Wichern F. Microbial Biomass Sulphur—An Important Yet Understudied Pool in Soil. Agronomy. 2021; 11(8):1606. https://doi.org/10.3390/agronomy11081606
Chicago/Turabian StyleHeinze, Stefanie, Michael Hemkemeyer, Sanja Annabell Schwalb, Khalid Saifullah Khan, Rainer Georg Joergensen, and Florian Wichern. 2021. "Microbial Biomass Sulphur—An Important Yet Understudied Pool in Soil" Agronomy 11, no. 8: 1606. https://doi.org/10.3390/agronomy11081606