
From Traditional Food to Functional Food? Evaluation of Malvaceae Species as Novel Food Crops


Abstract
:1. Introduction
2. Materials and Methods
2.1. Focal Species, Seed Collection
2.2. Research Setup
2.3. Plant Development
2.4. Phytochemical Nutritive Analysis of Leaves
2.4.1. Methanolic Extract Preparation
2.4.2. Total Phenolic Content (TPC)
2.4.3. Ferric-Reducing Antioxidant Power (FRAP)
2.4.4. Ascorbic Acid Determination
2.4.5. Total Protein
2.4.6. Total Dietary Fibers
2.4.7. Reducing Sugars Determination
2.4.8. Elemental Analysis and Non-Nutritive Compounds
2.5. Data Analysis
3. Results
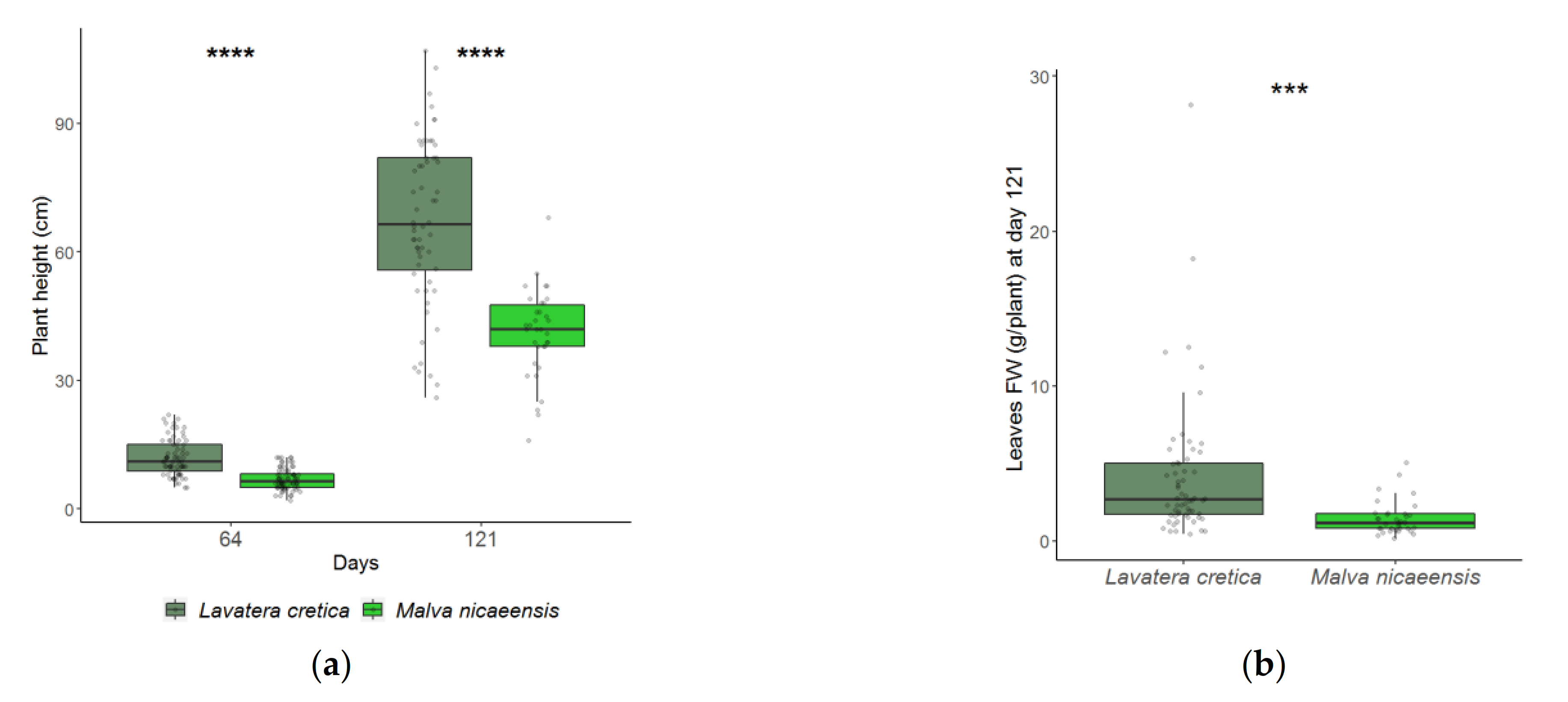
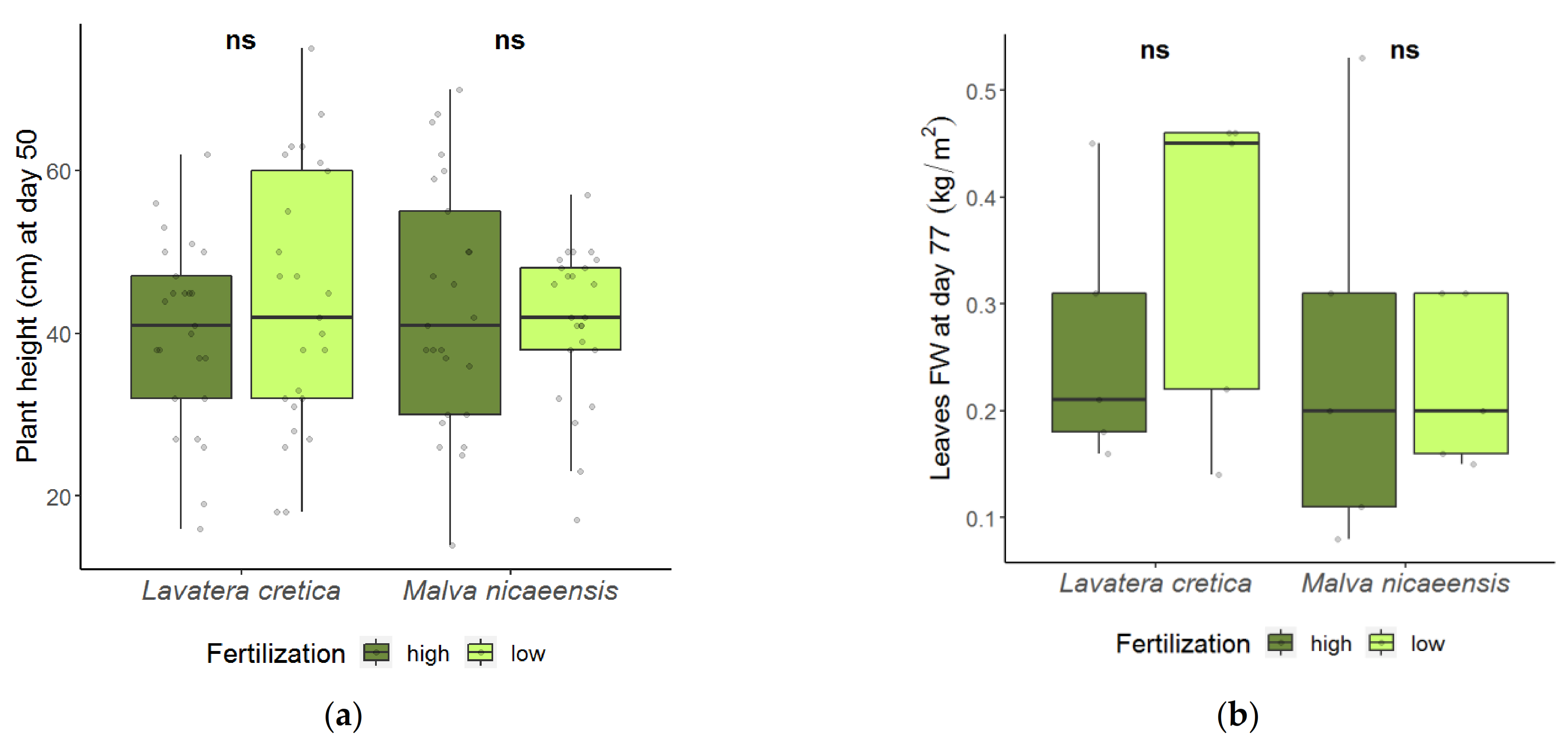
3.1. Plant Development
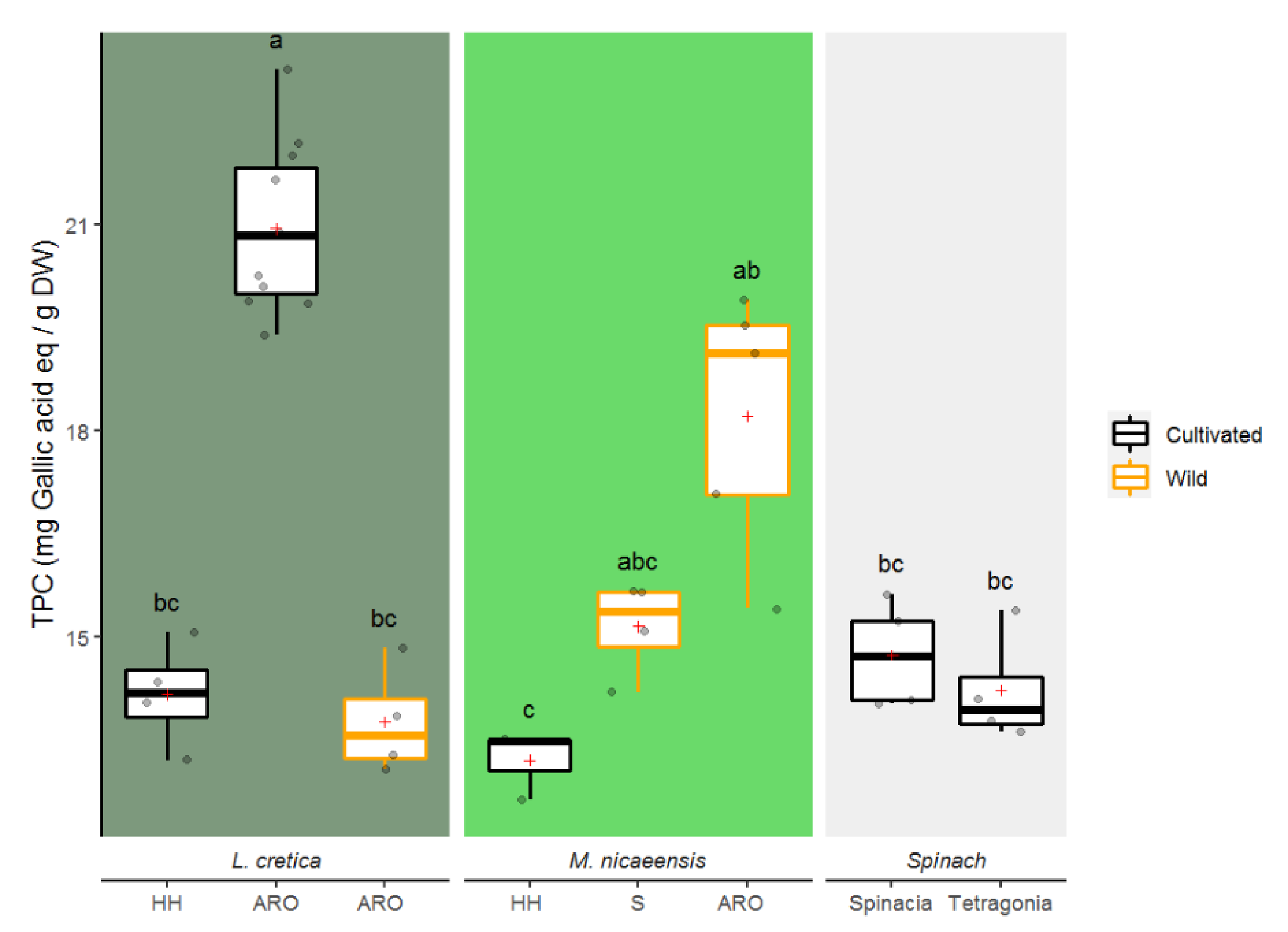
3.1.1. Total Phenolic Content
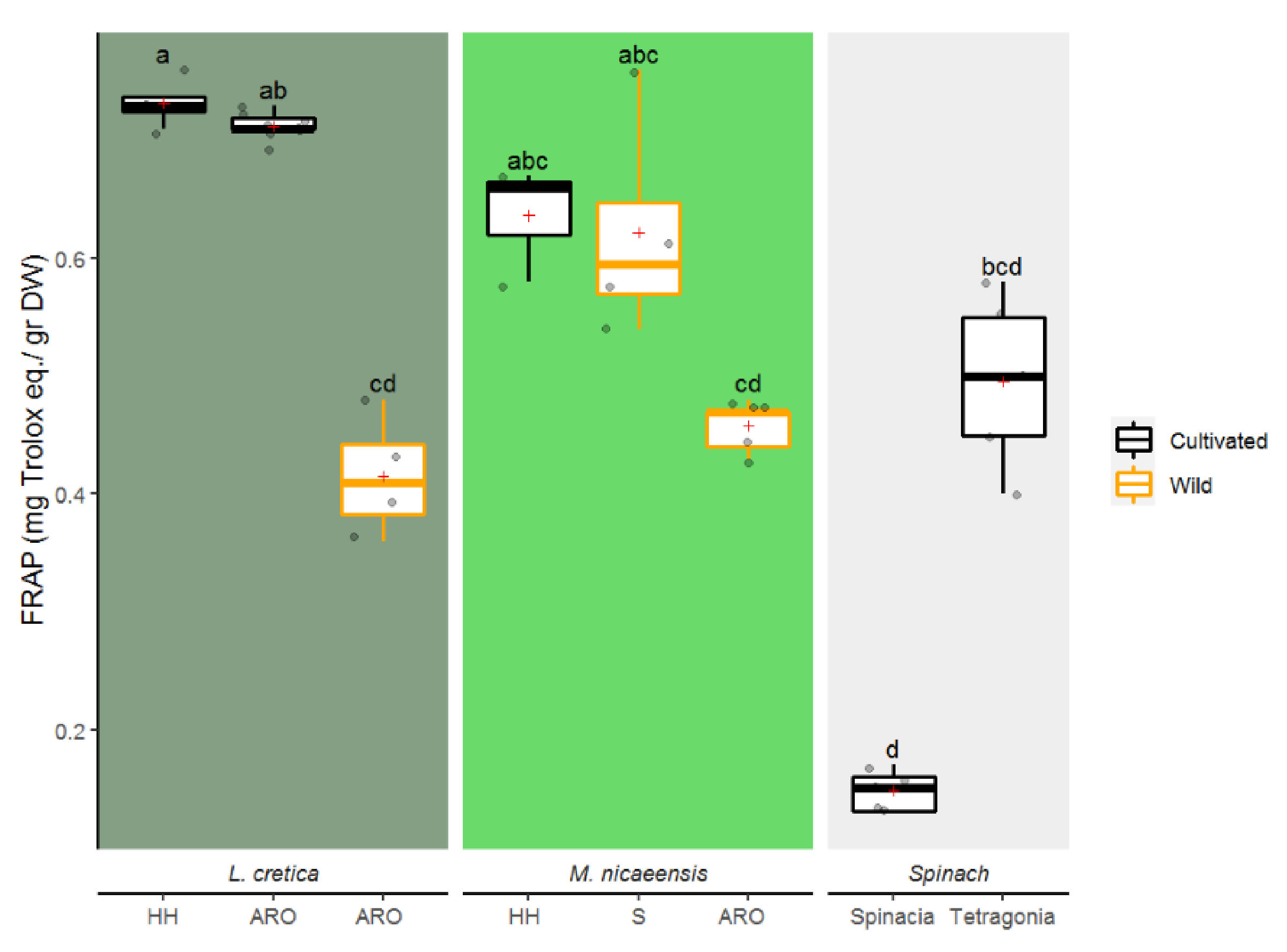
3.1.2. Antioxidative Activity
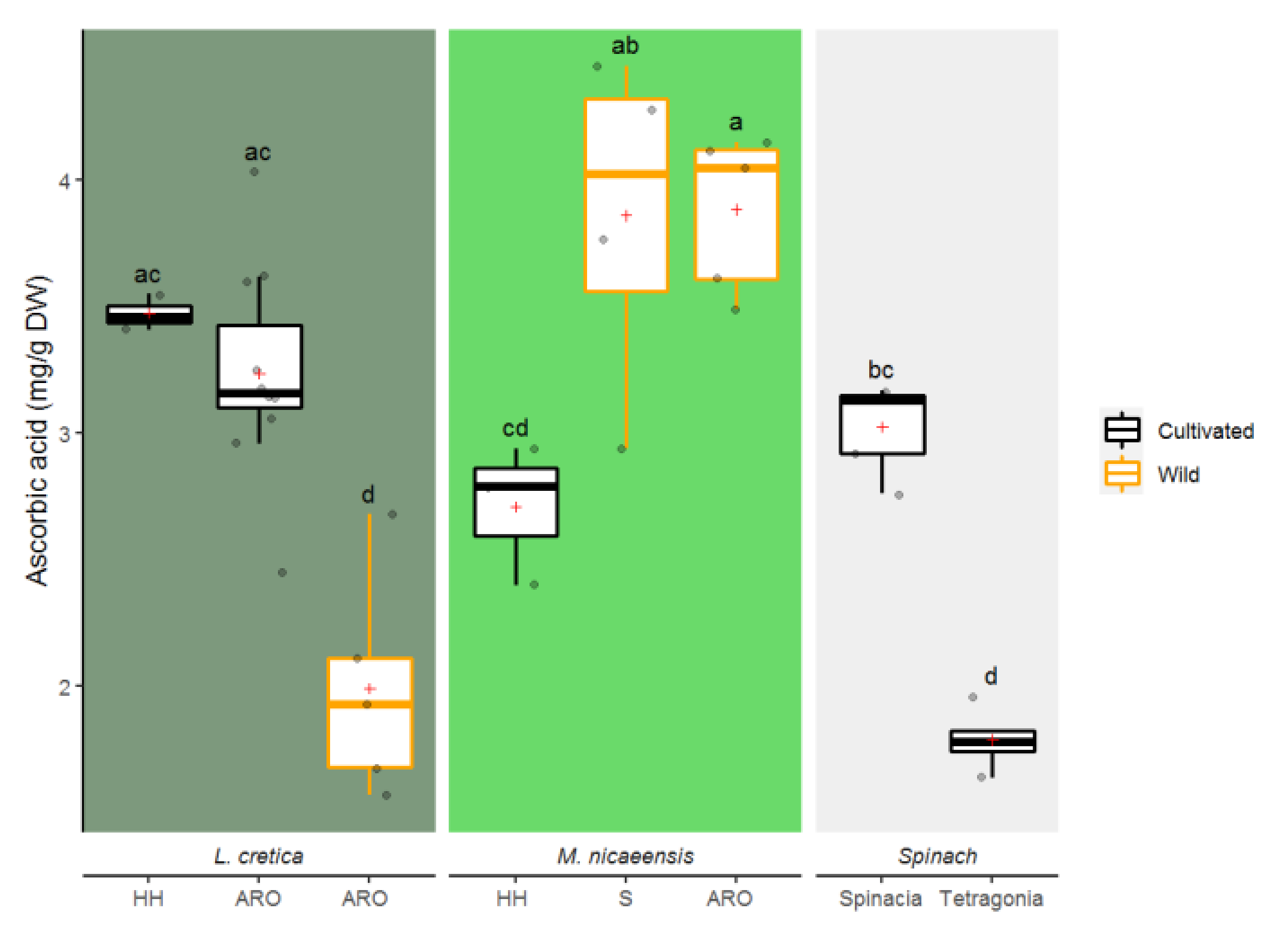
3.1.3. Ascorbic Acid
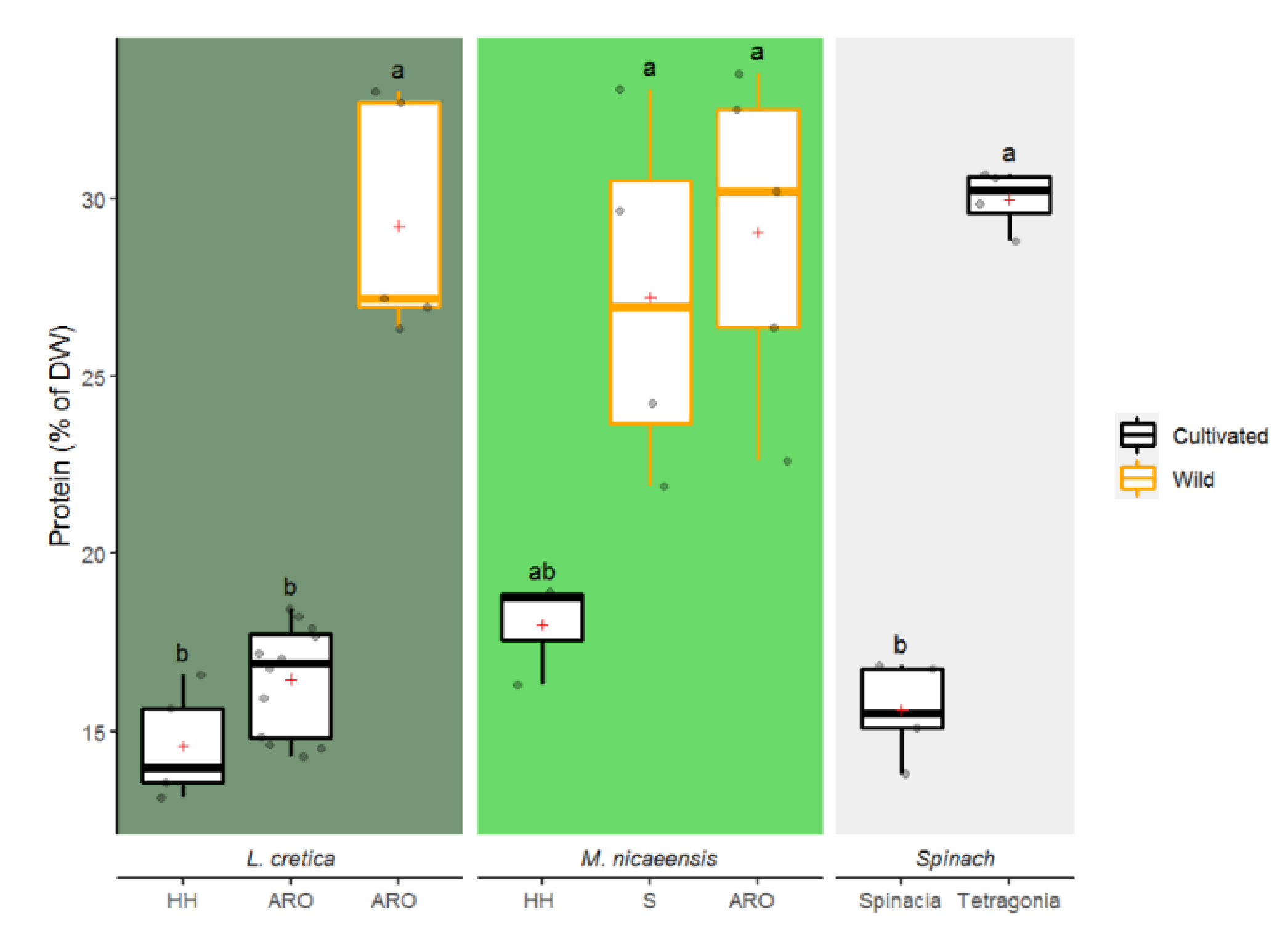
3.1.4. Total Protein
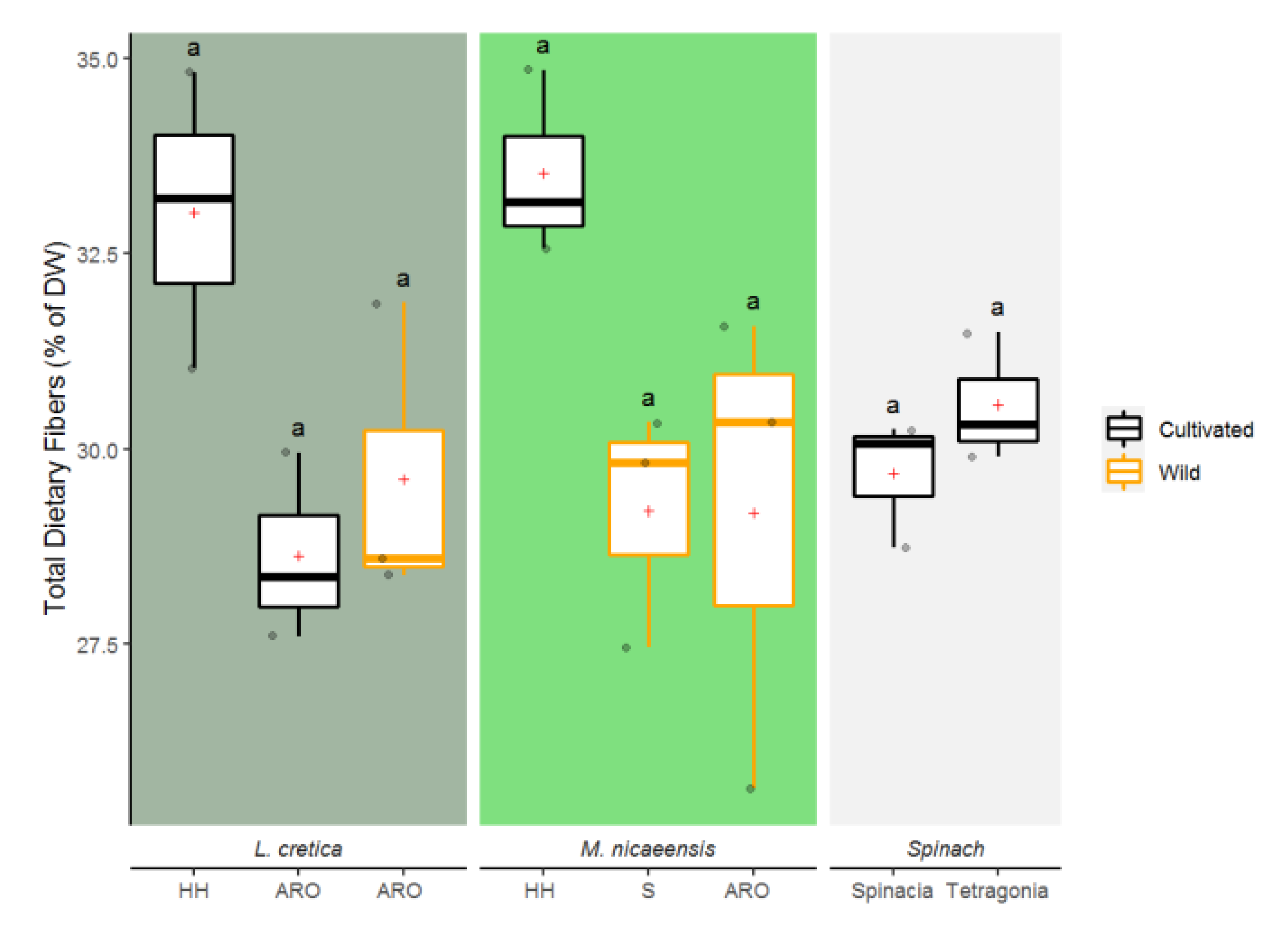
3.1.5. Total Dietary Fibers
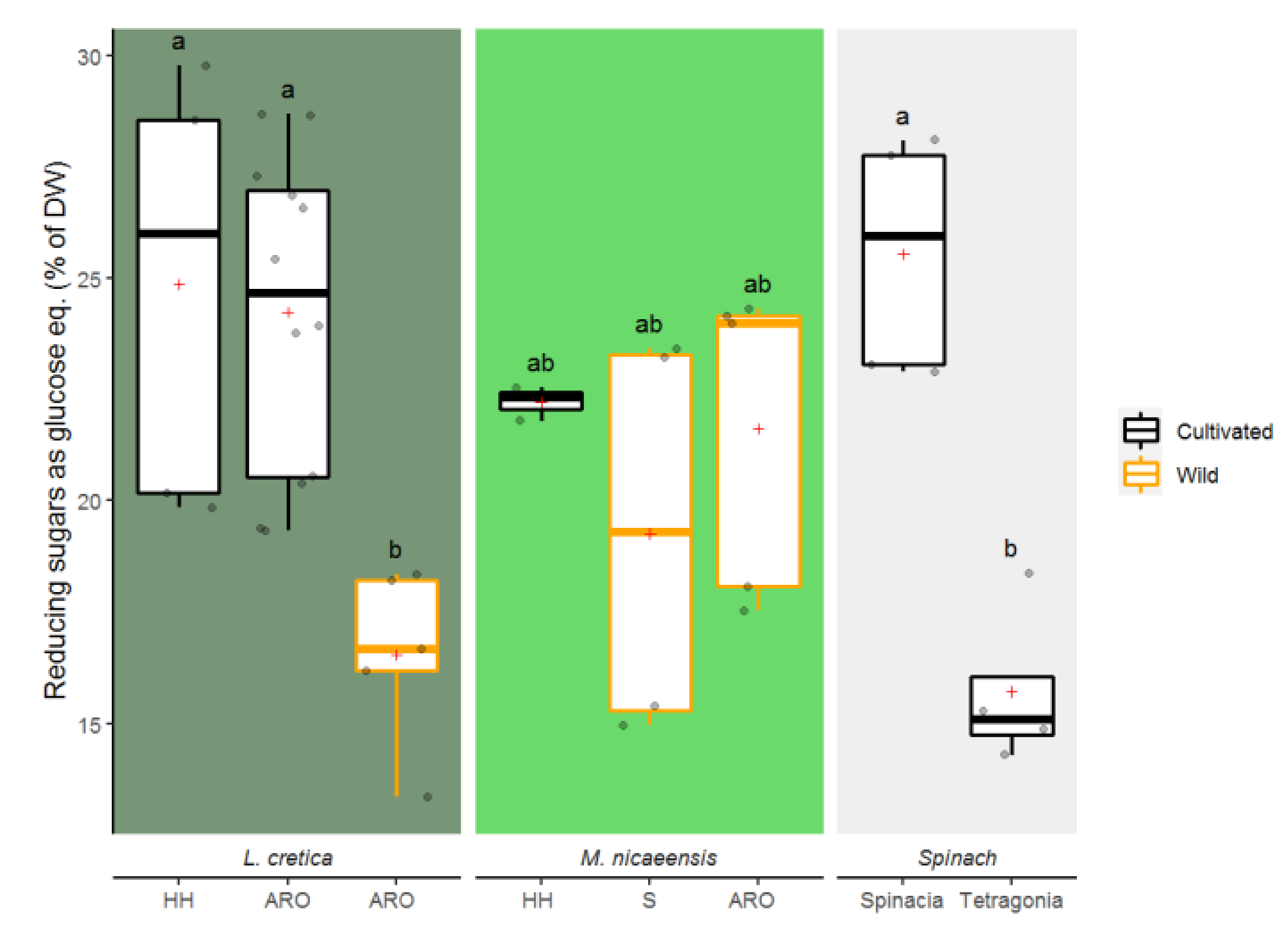
3.1.6. Reducing Sugars
3.1.7. Elemental Analysis and Non-Nutritive Compounds

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | L. cretica (n = 22) | M. nicaeensis (n = 12) | S. oleracea | T. tetragonioides | |||||
|---|---|---|---|---|---|---|---|---|---|
| Source | ARO Cultivated (n = 12) | H. Noi Cultivated (n = 5) | ARO F. Margins (n = 5) | H. Noi Cultivated (n = 3) | ARO F. Margins (n = 5) | Sandala F. Margins (n = 4) | Sandala Cultivated (n = 5) | Sandala Cultivated (n = 5) | |
| Element | Statistical test | mean ± sd/compact letter display of statistical differences | |||||||
| Al µg/g DW | Kruskal Walis & Duun | 78.45 ± 14.44 ab | 148.9 ± 129.6 ab | 147.9 ± 148.24 ab | 59.71 ± 4.59 b | 47.73 ± 18.60 b | 90.3 ± 13.50 ab | 145.18 ± 41.87 a | 80.03 ± 14.40 ab |
| Se µg/g DW | 1.21 ± 1.97 abc | 0 ± 0 c | 0 ± 0 c | 3.31 ± 1.65 a | 2.98 ± 1.38 abc | 2.48 ± 2.86 abc | 2.98 ± 2.16 ab | 0 ± 0 c | |
| Fe µg/g DW | 100.4 ± 18.83 cd | 102.5 ± 21.99 bcd | 84.48 ± 3.42 d | 89.57 ± 5.26 cd | 83.44 ± 2.45 d | 146.6 ± 8.03 ab | 170.03 ± 24.77 a | 120.54 ± 13.46 abc | |
| B µg/g DW | 13.39 ± 5.13 c | 17.91 ± 5.41 abc | 22.86 ± 2.74 a | 14.91 ± 4.33 c | 22.84 ± 13.7 c | 13.66 ± 2.06 c | 23.86 ± 3.44 ab | 20.70 ± 5.26 a | |
| Cu µg/g DW | 9.34 ± 1.43 ab | 4.97 ± 0.005 b | 15.91 ± 8.92 a | 6.37 ± 1.67 ab | 8.94 ± 1.47 ab | 7.46 ± 2.88 ab | 6.96 ± 2.72 ab | 8.88 ± 1.42 ab | |
| Mn µg/g DW | 48.34 ± 17.49 bc | 37.8 ± 5.18 bc | 49.67 ± 9.95 abc | 29.85 ± 0.10 c | 29.80 ± 12.14 c | 27.33 ± 2.94 c | 64.62 ± 4.68 ab | 207.17 ± 25.68 a | |
| Ba µg/g DW | 13.81 ± 2.85 a | 12.94 ± 4.15 ab | 9.94 ± 2.63 abc | 14.92 ± 4.34 ab | 3.97 ± 1.48 c | 9.93 ± 0.03 abc | 4.97 ± 0.004 bc | 3.94 ± 1.44 c | |
| K mg/g DW | 16.64 ± 2.07 c | 18.47 ± 1.11 bc | 24.95 ± 1.34 ab | 19.79 ± 1.59 abc | 22.37 ± 0.44 ab | 24.95 ± 1.49 ab | 37.17 ± 0.93 a | 20.02 ± 3.92 bc | |
| Mg mg/g DW | 2.46 ± 0.23 abc | 2.52 ± 0.12 cd | 2.92 ± 0.09 d | 2.54 ± 0.15 cd | 2.65 ± 0.22 cd | 2.96 ± 0.29 abcd | 4.94 ± 0.02 a | 4.35 ± 0.024 ab | |
| Ca mg/g DW | 18.45 ± 1.90 ab | 19.11 ± 1.39 ab | 21.881.73a | 19.74 ± 0.71 ab | 20.38 ± 0.35 a | 18.15 ± 0.32 abc | 11.34 ± 0.21 c | 13.23 ± 0.57 bc | |
| Na mg/g DW | ANOVA & Tukey | 1.96 ± 0.73 e | 7.87 ± 1.39 cd | 5.10 ± 1.71 d | 12.51 ± 1.38 bc | 1.87 ± 0.68 e | 6.80 ± 0.51 cd | 24.28 ± 7.12 b | 67.82 ± 1.40 a |
| P mg/g DW | 5.06 ± 0.41 b | 4.15 ± 0.39 c | 6.67 ± 0.09 a | 4.83 ± 0.49 bc | 6.22 ± 0.10 a | 5.21 ± 0.54 b | 6.04 ± 0.18 a | 4.30 ± 0.38 c | |
| Cl mg/g DW | 7.1 ± 1.42 de | 7.10 ± 1.42 bc | 6.12 ± 1.98 c | 9.9 ± 2.03 ac | 2.36 ± 0.77 e | 5.37 ± 0.68 cd | 16.5 ± 1.28 a | 11.86 ± 4.12 ab | |
4. Discussion
| Species | L. cretica (n = 22) | M. nicaeensis (n = 12) | S. oleracea | T. tetragonioides | |||||
|---|---|---|---|---|---|---|---|---|---|
| Source | ARO Cultivated (n = 12) | H. Noi Cultivated (n = 5) | ARO F. Margins (n = 5) | H. Noi Cultivated (n = 3) | ARO F. Margins (n = 5) | Sandala F. Margins (n = 4) | Sandala Cultivated (n = 5) | Sandala Cultivated (n = 5) | |
| Molecule | Statistical test | mean ± sd compact letter display of statistical differences | |||||||
| NO3 mg/g DW | Kruskal Walis & Duun | 0 ± 0 b | 0 ± 0 b | 8.5 ± 3.74 a | 1.2 ± 0.52 ab | 3.62 ± 2.18 a | 5.88 ± 3.57 a | 1.96 ± 1.35 ab | 23.2 ± 13.7 a |
| OA mg/g DW | 0 ± 0 b | 0 ± 0 b | 0 ± 0 b | 0 ± 0 b | 0 ± 0 b | 0 ± 0 b | 0.45 ±0.11 a | 0.56 ± 0.11 a | |
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tilman, D.; Cassman, K.G.; Matson, P.A.; Naylor, R.; Polasky, S. Agricultural sustainability and intensive production practices. Nature 2002, 418, 671–677. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.L.; Swallow, B.M. Optimizing expenditures for agricultural land conservation: Spatially-explicit estimation of benefits, budgets, costs and targets. Land Use Policy 2016, 59, 272–283. [Google Scholar] [CrossRef]
- FAO; IFAD; UNICEF; WFP; WHO. The State of Food Security and Nutrition in the World 2020. Transforming Food Systems for Affordable Healthy Diets; FAO: Rome, Italy, 2020; p. 320. [Google Scholar]
- Altieri, M.A. The ecological role of biodiversity in agroecosystems. Agric. Ecosyst. Environ. 1999, 74, 19–31. [Google Scholar] [CrossRef] [Green Version]
- Adams, M.W.; Ellingboe, A.H.; Rossman, E.C. Biological uniformity and disease epidemics. BioScience 1971, 21, 1067–1070. [Google Scholar] [CrossRef]
- Shukla, P.R.; Skea, J.; Calvo Buendia, E.; Masson-Delmotte, V.; Pörtner, H.-O.; Roberts, D.C.; Zhai, P.; Slade, R.; Connors, S.; van Diemen, R.; et al. IPCC, 2019: Climate Change and Land: An IPCC Special Report on Climate Change, Desertification, Land Degradation, Sustainable Land Management, Food Security, and Greenhouse Gas Fluxes in Terrestrial Ecosystems; IPCC: Geneva, Switzerland, 2019. [Google Scholar]
- Peel, M.C.; Finlayson, B.L.; McMahon, T.A. Updated world map of the Koppen-Geiger climate classification. Hydrol. Earth Syst. Sci. 2007, 11, 1633–1644. [Google Scholar] [CrossRef] [Green Version]
- Rewald, B.; Eppel, A.; Shelef, O.; Hill, A.; Degu, A.; Friedjung, A.; Rachmilevitch, S. Hot desert environments. In Life at Extremes: Environments, Organisms and Strategies for Survival; Bell, E., Ed.; CABI: Wallingford, UK, 2012; pp. 196–218. [Google Scholar]
- Shelef, O.; Guy, O.; Solowey, E.; Kam, M.; Degen, A.A.; Rachmilevitch, S. Domestication of plants for sustainable agriculture in drylands: Experience from the Negev Desert. Arid Land Res. Manag. 2016, 30, 209–228. [Google Scholar] [CrossRef]
- Shelef, O.; Fernández-Bayo, J.D.; Sher, Y.; Ancona, V.; Slinn, H.; Achmon, Y. Elucidating Local Food Production to Identify the Principles and Challenges of Sustainable Agriculture. In Sustainable Food Systems from Agriculture to Industry, 1st ed.; Galanakis, C.M., Ed.; Elsevier Inc.: London, UK, 2018; pp. 47–81. [Google Scholar] [CrossRef]
- Shelef, O.; Weisberg, P.J.; Provenza, F.D. The Value of Native Plants and Local Production in an Era of Global Agriculture. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Davies, W.J.; Zhang, J.; Yang, J.; Dodd, I.C. Novel crop science to improve yield and resource use efficiency in water-limited agriculture. J. Agric. Sci. 2011, 149, 123–131. [Google Scholar] [CrossRef]
- Gupta, A.K. Origin of agriculture and domestication of plants and animals linked to early Holocene climate amelioration. Curr. Sci. 2004, 87, 54–59. [Google Scholar]
- Mayer-Chissick, U.; Lev, E. Wild edible plants in Israel tradition versus cultivation. In Medicinal and Aromatic Plants of the Middle-East; Yaniv, Z., Dudai, N., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 9–26. [Google Scholar]
- Ben-Simchon, E.; Sapir, E.; Vaknin, Y.; Shelef, O. Malvaceae spp. leaves as a novel crop for food. Int. J. Agric. For. Life Sci. 2019, 3, 279–286. [Google Scholar]
- Feinbrun-Dothan, N.A.; Danin, A. Analytical Flora of Eretz-Israel; Plitmann, U., Ed.; CANA Publishing House Ltd.: Jerusalem, Israel, 1998; p. 1008. [Google Scholar]
- Shehata, H.S.; Galal, T.M. Factors affecting the distribution and associated species of Malva parviflora in the Nile Delta, Egypt. Weed Biol. Manag. 2015, 15, 42–52. [Google Scholar] [CrossRef]
- Barros, L.; Carvalho, A.M.; Ferreira, I.C.F.R. Leaves, flowers, immature fruits and leafy flowered stems of Malva sylvestris: A comparative study of the nutraceutical potential and composition. Food Chem. Toxicol. 2010, 48, 1466–1472. [Google Scholar] [CrossRef]
- Guarrera, P.M. Food medicine and minor nourishment in the folk traditions of Central Italy (Marche, Abruzzo and Latium). Fitoterapia 2003, 74, 515–544. [Google Scholar] [CrossRef]
- Cooper, M.R.; Johnson, A.W. Poisonous Plants in Britain and Their Effects on Animals and Man; HM Stationery Office: London, UK, 1984. [Google Scholar]
- Koller, D.; Levitan, I. Diurnal Phototropism in Leaves of Lavatera-Cretica L under Conditions of Simulated Solar-Tracking. J. Exp. Botany 1989, 40, 1059–1064. [Google Scholar] [CrossRef]
- Ray, M.F. New combinations in Malva (Malvaceae: Malveae). Novon 1998, 8, 288–295. [Google Scholar] [CrossRef]
- Moreau, D.; Milard, G.; Munier-Jolain, N. A plant nitrophily index based on plant leaf area response to soil nitrogen availability. Agron. Sustain. Dev. 2013, 33, 809–815. [Google Scholar] [CrossRef]
- Santamaria, P. Nitrate in vegetables: Toxicity, content, intake and EC regulation. J. Sci. Food Agr. 2006, 86, 10–17. [Google Scholar] [CrossRef]
- Constantin, J.; Mary, B.; Laurent, F.; Aubrion, G.; Fontaine, A.; Kerveillant, P.; Beaudoin, N. Effects of catch crops, no till and reduced nitrogen fertilization on nitrogen leaching and balance in three long-term experiments. Agric. Ecosyst. Environ. 2010, 135, 268–278. [Google Scholar] [CrossRef]
- Meisinger, J.J.; Delgado, J.A. Principles for managing nitrogen leaching. J. Soil Water Conserv. 2002, 57, 485–498. [Google Scholar]
- Vinograd, A.; Zaady, E.; Kigel, J. Abandoned corrals: Colonization and vegetation recovery of ephemeral habitats in silvo-pastoral systems. J. Plant Ecol. 2020, 13, 722–731. [Google Scholar] [CrossRef]
- Chauhan, B.S.; Gill, G.; Preston, C. Factors affecting seed germination of little mallow (Malva parviflora) in southern Australia. Weed Sci. 2006, 54, 1045–1050. [Google Scholar] [CrossRef]
- Faulkner, G.P.; Pourshahidi, L.K.; Wallace, J.M.W.; Kerr, M.A.; McCrorie, T.A.; Livingstone, M.B.E. Serving size guidance for consumers: Is it effective? Proc. Nutr. Soc. 2012, 71, 610–621. [Google Scholar] [CrossRef] [Green Version]
- Singleton, V.L. Citation Classic-Colorimetry of Total Phenolics with Phosphomolybdic-Phosphotungstic Acid Reagents. Cc/Agr. Biol. Environ. 1985, 48, 18. [Google Scholar]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nobrega, J.A.; Lopes, G.S. Flow-injection spectrophotometric determination of ascorbic acid in pharmaceutical products with the Prussian Blue reaction. Talanta 1996, 43, 971–976. [Google Scholar] [CrossRef]
- Matei, N.; Dobrinas, S.; Radu, G.L. Spectrophotometric determination of ascorbic acid in grapes with the Prussian Blue reaction. An. Univ. Ovidius Constanta-Ser. Chim. 2012, 2, 23. [Google Scholar] [CrossRef]
- Kjeldahl, J.G.C.T. New method for the determination of nitrogen in organic bodies. J. Anal. Chem. 1883, 22, 366–382. [Google Scholar]
- Mariotti, F.; Tome, D.; Mirand, P.P. Converting nitrogen into Protein-Beyond 6.25 and Jones’ factors. Crit. Rev. Food Sci. Nutr. 2008, 48, 177–184. [Google Scholar] [CrossRef]
- Lindsay, H. A colorimetric estimation of reducing sugars in potatoes with 3, 5-dinitrosalicylic acid. Potato Res. 1973, 16, 176–179. [Google Scholar] [CrossRef]
- Chua, M.; Chan, K.; Hocking, T.J.; Williams, P.A.; Perry, C.J.; Baldwin, T.C. Methodologies for the extraction and analysis of konjac glucomannan from corms of Amorphophallus konjac K. Koch. Carbohyd. Polym. 2012, 87, 2202–2210. [Google Scholar] [CrossRef]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Bates, D.; Machler, M.; Bolker, B.M.; Walker, S.C. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biometrical. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [Green Version]
- Kassambara, A. ggpubr: ‘ggplot2’based Publication Ready Plots; R Package Version 0.4.0; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Ogle, D.H.; Wheeler, P.; Dinno, A. FSA: Fisheries Stock Analysis; R Package Version 0.8.32; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Zouari, N.; Fakhfakh, N.; Zouari, S.; Sellami, M.; Abid, M.; Ayadi, M.A.; Zaidi, S.; Neffati, M. Volatile and lipid analyses by gas chromatography/mass spectrometry and nutraceutical potential of edible wild Malva aegyptiaca L.(Malvaceae). Int. J. Food Sci. Nutr. 2011, 62, 600–608. [Google Scholar] [CrossRef]
- Khan, H.; Jan, S.A.; Javed, M.; Shaheen, R.; Khan, Z.; Ahmad, A.; Zaman Safi, S.; Imran, M. Nutritional composition, antioxidant and antimicrobial activities of selected wild edible plants. J. Food Biochem. 2016, 40, 61–70. [Google Scholar] [CrossRef]
- Mehtap, Ö.Z.E.R.; Aksoy, M. Mineral composition and nutritional properties of Malva neglecta and Malvella sherardiana consumed as vegetable in Central Black Sea Region of Turkey. Turk. J. Food Agric. Sci. 2019, 1, 18–23. [Google Scholar]
- Conforti, F.; Sosa, S.; Marrelli, M.; Menichini, F.; Statti, G.A.; Uzunov, D.; Tubaro, A.; Menichini, F.; Della Loggia, R. In vivo anti-inflammatory and in vitro antioxidant activities of Mediterranean dietary plants. J. Ethnopharmacol. 2008, 116, 144–151. [Google Scholar] [CrossRef]
- Brennan, C.S. Dietary fibre, glycaemic response, and diabetes. Mol. Nutr. Food Res. 2005, 49, 560–570. [Google Scholar] [CrossRef]
- Fardet, A.; Leenhardt, F.; Lioger, D.; Scalbert, A.; Rémésy, C. Parameters controlling the glycaemic response to breads. Nutr. Res. Rev. 2006, 19, 18–25. [Google Scholar] [CrossRef]
- Heaney, R.P.; Weaver, C.M. Oxalate-Effect on Calcium Absorbability. Am. J. Clin. Nutr. 1989, 50, 830–832. [Google Scholar] [CrossRef]
- Kelsay, J.L.; Prather, E.S. Mineral balances of human subjects consuming spinach in a low-fiber diet and in a diet containing fruits and vegetables. Am. J. Clin. Nutr. 1983, 38, 12–19. [Google Scholar] [CrossRef]
- Noonan, S.C.; Savage, G.P. Oxalate content of foods and its effect on humans. Asia Pac. J. Clin. Nutr. 1999, 8, 64–74. [Google Scholar] [PubMed]
- Bonsmann, S.S.; Walczyk, T.; Renggli, S.; Hurrell, R.F. Oxalic acid does not influence nonhaem iron absorption in humans: A comparison of kale and spinach meals. Eur. J. Clin. Nutr. 2008, 62, 336–341. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Basheer, L.; Ben-Simchon, E.; Cohen, A.; Shelef, O. From Traditional Food to Functional Food? Evaluation of Malvaceae Species as Novel Food Crops. Agronomy 2021, 11, 1294. https://doi.org/10.3390/agronomy11071294
Basheer L, Ben-Simchon E, Cohen A, Shelef O. From Traditional Food to Functional Food? Evaluation of Malvaceae Species as Novel Food Crops. Agronomy. 2021; 11(7):1294. https://doi.org/10.3390/agronomy11071294
Chicago/Turabian StyleBasheer, Loai, Eyal Ben-Simchon, Alisa Cohen, and Oren Shelef. 2021. "From Traditional Food to Functional Food? Evaluation of Malvaceae Species as Novel Food Crops" Agronomy 11, no. 7: 1294. https://doi.org/10.3390/agronomy11071294