
The Effect of Biochar-Based Organic Amendments on the Structure of Soil Bacterial Community and Yield of Maize (Zea mays L.)

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. The Study Site
2.2. Identification of Soil Microorganisms
2.2.1. DNA Extraction
2.2.2. PCR Amplification
2.3. Soil Enzymes Analyses
2.4. Plant Analyses
2.5. Statistical and Bioinformatics Analyses
3. Results and Discussion
3.1. Abundance and Composition of Bacterial Community in Soil
3.2. Changes in Soil pH and Dehydrogenase Activity
3.3. Plant Parameters
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| DHA | Dehydrogenase activity |
| TTC | 2, 3, 5-triphenyltetrazolium chloride |
| CAT | Catalase activity |
| BIF | Biological Index of Fertility |
| OTU | Bacterial operational taxonomic units |
| CVA | Canonical variate analysis |
| AM | Arbuscular mycorrhiza |
References
- Davydov, R.; Sokolov, M.; Hogland, W.; Glinushkin, A.; Markaryan, A. The application of pesticides and mineral fertilizers in agriculture. In Proceedings of the EECE—2018, MATEC Web of Conferences, St. Petersburg, Russia, 19–20 November 2018; EDP Sciences: Les Ulis, France, 2018; p. 11003. [Google Scholar]
- Zhong, W.; Gu, T.; Wang, W.; Zhang, B.; Lin, X.; Huang, Q.; Shen, W. The effects of mineral fertilizer and organic manure on soil microbial community and diversity. Plant Soil 2010, 326, 511–522. [Google Scholar] [CrossRef]
- Geisseler, D.; Scow, K.M. Long-term effects of mineral fertilizers on soil microorganisms–A review. Soil Biol. Biochem. 2014, 75, 54–63. [Google Scholar] [CrossRef]
- Alkharabsheh, H.M.; Seleiman, M.F.; Battaglia, M.L.; Shami, A.; Jalal, R.S.; Alhammad, B.A.; Almutairi, K.F.; Al-Saif, A.M. Biochar and its broad impacts in soil quality and fertility, nutrient leaching and crop productivity: A review. Agronomy 2021, 11, 993. [Google Scholar] [CrossRef]
- Grzyb, A.; Wolna-Maruwka, A.; Niewiadomska, A. Environmental factors affecting the mineralization of crop residues. Agronomy 2020, 10, 1951. [Google Scholar] [CrossRef]
- Butterly, C.R.; Baldock, J.A.; Tang, C. The contribution of crop residues to changes in soil ph under field conditions. Plant Soil 2013, 366, 185–198. [Google Scholar] [CrossRef]
- Whalen, J.K. Managing soil biota-mediated decomposition and nutrient mineralization in sustainable agroecosystems. Adv. Agric. 2014, 2014, 1–13. [Google Scholar] [CrossRef]
- Gorovtsov, A.V.; Minkina, T.M.; Mandzhieva, S.S.; Perelomov, L.V.; Soja, G.; Zamulina, I.V.; Rajput, V.D.; Sushkova, S.N.; Mohan, D.; Yao, J. The mechanisms of biochar interactions with microorganisms in soil. Environ. Geochem. Health 2019, 42, 1–24. [Google Scholar] [CrossRef]
- Gul, S.; Whalen, J.K.; Thomas, B.W.; Sachdeva, V.; Deng, H. Physico-chemical properties and microbial responses in biochar-amended soils: Mechanisms and future directions. Agric. Ecosys. Environ. 2015, 206, 46–59. [Google Scholar] [CrossRef]
- Uchimiya, M.; Wartelle, L.H.; Klasson, K.T.; Fortier, C.A.; Lima, I.M. Influence of pyrolysis temperature on biochar property and function as a heavy metal sorbent in soil. J. Agric. Food Chem. 2011, 59, 2501–2510. [Google Scholar] [CrossRef]
- Cao, X.; Harris, W. Properties of dairy-manure-derived biochar pertinent to its potential use in remediation. Bioresour. Technol. 2010, 101, 5222–5228. [Google Scholar] [CrossRef]
- Quilliam, R.S.; Glanville, H.C.; Wade, S.C.; Jones, D.L. Life in the ‘Charosphere’–Does biochar in agricultural soil provide a significant habitat for microorganisms? Soil Biol. Biochem. 2013, 65, 287–293. [Google Scholar] [CrossRef]
- Man, Y.; Wang, B.; Wang, J.; Slanỳ, M.; Yan, H.; Li, P.; El-Naggar, A.; Shaheen, S.M.; Rinklebe, J.; Feng, X. Use of biochar to reduce mercury accumulation in Oryza sativa L: A trial for sustainable management of historically polluted farmlands. Environ. Int. 2021, 153, 106527. [Google Scholar] [CrossRef]
- Blackwell, P.; Krull, E.; Butler, G.; Herbert, A.; Solaiman, Z. Effect of banded biochar on dryland wheat production and fertiliser use in south-western Australia: An agronomic and economic perspective. Soil Res. 2010, 48, 531–545. [Google Scholar] [CrossRef]
- Palencia, P.; Martínez, F.; Pestana, M.; Oliveira, J.A.; Correia, P.J. Effect of Bacillus velezensis and Glomus intraradices on fruit quality and growth parameters in strawberry soilless growing system. J. Hortic. 2015, 84, 122–130. [Google Scholar] [CrossRef] [Green Version]
- Chandanie, W.A.; Kubota, M.; Hyakumachi, M. Interactions between the arbuscular mycorrhizal fungus Glomus mosseae and plant growth-promoting fungi and their significance for enhancing plant growth and suppressing damping-off of cucumber (Cucumis sativus L.). Appl. Soil Ecol. 2009, 41, 336–341. [Google Scholar] [CrossRef]
- Elsharkawy, M.M.; Shimizu, M.; Takahashi, H.; Hyakumachi, M. The plant growth-promoting fungus fusarium equiseti and the arbuscular mycorrhizal fungus Glomus mosseae induce systemic resistance against cucumber mosaic virus in cucumber plants. Plant Soil 2012, 361, 397–409. [Google Scholar] [CrossRef]
- Colla, G.; Rouphael, Y.; Di Mattia, E.; El-Nakhel, C.; Cardarelli, M. Co-inoculation of Glomus intraradices and Trichoderma atroviride acts as a biostimulant to promote growth, yield and nutrient uptake of vegetable crops. J. Sci. Food Agric. 2015, 95, 1706–1715. [Google Scholar] [CrossRef]
- Jemil, I.; Jridi, M.; Nasri, R.; Ktari, N.; Salem, R.B.S.-B.; Mehiri, M.; Hajji, M.; Nasri, M. Functional, antioxidant and antibacterial properties of protein hydrolysates prepared from fish meat fermented by Bacillus subtilis A26. Process Biochem. 2014, 49, 963–972. [Google Scholar] [CrossRef]
- Lim, J.-H.; Kim, S.-D. Synergistic plant growth promotion by the indigenous auxins-producing PGPR Bacillus subtilis AH18 and Bacillus licheniforims K11. J. Korean Soc. Appl. Biol. Chem. 2009, 52, 531–538. [Google Scholar] [CrossRef]
- Grover, M.; Madhubala, R.; Ali, S.Z.; Yadav, S.K.; Venkateswarlu, B. Influence of Bacillus spp. strains on seedling growth and physiological parameters of sorghum under moisture stress conditions. J. Basic Microbiol. 2014, 54, 951–961. [Google Scholar] [CrossRef]
- Jones, N.; Ray, B.; Ranjit, K.T.; Manna, A.C. Antibacterial activity of ZnO nanoparticle suspensions on a broad spectrum of microorganisms. FEMS Microbiol. Lett. 2008, 279, 71–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tate, J.J.; Gutierrez-Wing, M.T.; Rusch, K.A.; Benton, M.G. The Effects of plant growth substances and mixed cultures on growth and metabolite production of green algae Chlorella sp.: A review. J. Plant Growth Regul. 2013, 32, 417–428. [Google Scholar] [CrossRef]
- Win, T.T.; Barone, G.D.; Secundo, F.; Fu, P. Algal Biofertilizers and plant growth stimulants for sustainable agriculture. Ind. Biotechnol. 2018, 14, 203–211. [Google Scholar] [CrossRef]
- Bhagavathy, S.; Sumathi, P.; Bell, I.J.S. Green algae Chlorococcum humicola—A new source of bioactive compounds with antimicrobial activity. Asian Pac. J. Trop. Biomed. 2011, 1, S1–S7. [Google Scholar] [CrossRef]
- Abu-Ghannam, N.; Rajauria, G. Antimicrobial activity of compounds isolated from algae. In Functional Ingredients from Algae for Foods and Nutraceuticals; Domínguez, H., Ed.; Woodhead Publishing: Cambridge, UK, 2013; pp. 287–306. [Google Scholar]
- Ducey, T.F.; Novak, J.M.; Johnson, M.G. Effects of biochar blends on microbial community composition in two coastal plain soils. Agriculture 2015, 5, 1060–1075. [Google Scholar] [CrossRef] [Green Version]
- Sheng, Y.; Zhu, L. Biochar alters microbial community and carbon sequestration potential across different soil pH. Sci. Total Environ. 2018, 622, 1391–1399. [Google Scholar] [CrossRef]
- Cheng, H.; Wang, J.; Tu, C.; Lin, S.; Xing, D.; Hill, P.; Chadwick, D.; Jones, D.L. Arbuscular mycorrhizal fungi and biochar influence simazine decomposition and leaching. GCB Bioenergy 2021, 13, 708–718. [Google Scholar] [CrossRef]
- Liu, L.; Li, J.; Yue, F.; Yan, X.; Wang, F.; Bloszies, S.; Wang, Y. Effects of arbuscular mycorrhizal inoculation and biochar amendment on maize growth, cadmium uptake and soil cadmium speciation in Cd-contaminated soil. Chemosphere 2018, 194, 495–503. [Google Scholar] [CrossRef]
- Palanisamy, M.; Iniyakumar, M.; Elaiyaraju, P.; Uthandi, S.M.S.; Sivasubramanian, V. Effect of application of algal biochar on soil enzymes. J Algal Biomass Util. 2017, 8, 1–9. [Google Scholar]
- European Parliament. Communication from the Commission to the European Parliament, The Council, the European Economic and Social Committee and the Committee of the Regions EU—Biodiversity Strategy for 2030—Bringing Nature Back into Our Lives (COM/2020/380); European Parliament: Brussels, Belgium, 2020. [Google Scholar]
- Camiña, F.; Trasar-Cepeda, C.; Gil-Sotres, F.; Leirós, C. Measurement of dehydrogenase activity in acid soils rich in organic matter. Soil Biol. Biochem. 1998, 30, 1005–1011. [Google Scholar] [CrossRef]
- Rodríguez-Kábana, R.; Truelove, B. Effects of crop rotation and fertilization on catalase activity in a soil of the southeastern United States. Plant Soil 1982, 69, 97–104. [Google Scholar] [CrossRef]
- Stefanic, G.; Eliade, G.; Chirnogeanu, I. Researches concerning a biological index of soil fertility. In Proceedings of the 5. Symposium on Soil Biology, Jassy, Romania, 17 February 1981. [Google Scholar]
- EPPO. Fusarium Ear Rot of Maize. EPPO Bull. 2015, 45, 336–339. [Google Scholar] [CrossRef]
- Commission Regulation (EC) No 152/2009 of 27 January 2009 laying down the methods of sampling and analysis for the official control of feed. Off. J. Eur. Union I 2009, 54, 1–130.
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Murali, A.; Bhargava, A.; Wright, E.S. IDTAXA: A novel approach for accurate taxonomic classification of microbiome sequences. Microbiome 2018, 6, 140. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. Phyloseq: An r package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [Green Version]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef]
- Větrovský, T.; Baldrian, P.; Morais, D. SEED 2: A user-friendly platform for amplicon high-throughput sequencing data analyses. Bioinformatics 2018, 34, 2292–2294. [Google Scholar] [CrossRef]
- Kayzer, D. A Note on testing hypotheses concerning interaction with special reference to a graphical presentation in the space of canonical variates. Biom. Lett. 2019, 56, 89–104. [Google Scholar] [CrossRef] [Green Version]
- Kayzer, D.; Frankowski, P.; Zbierska, J.; Staniszewski, R. Evaluation of trophic parameters in newly built reservoir using canonical variates analysis. In Proceedings of the XLVIII Seminar of Applied Mathematics, Boguszów-Gorce, Poland, 9–11 September 2018; Volume 23, p. 00019. [Google Scholar]
- Staniszewski, R.; Frankowski, P.; Kayzer, D.; Zbierska, J.; Achtenberg, K. Reconstruction of ancient lake after peat excavation—A case study about water quality. Appl. Sci. 2021, 11, 4213. [Google Scholar] [CrossRef]
- Campbell, N.A.; Atchley, W.R. The geometry of canonical variate analysis. Syst. Biol. 1981, 30, 268–280. [Google Scholar] [CrossRef]
- Lejeune, M.; Caliński, T. Canonical analysis applied to multivariate analysis of variance. J. Multivar. Anal. 2000, 72, 100–119. [Google Scholar] [CrossRef] [Green Version]
- Kayzer, D.; Borowiak, K.; Budka, A.; Zbierska, J. Study of interaction in bioindication research on tobacco plant injuries caused by ground level ozone. Environmetrics 2009, 20, 666–675. [Google Scholar] [CrossRef]
- Furtak, K.; Grządziel, J.; Gałązka, A.; Niedźwiecki, J. Prevalence of unclassified bacteria in the soil bacterial community from floodplain meadows (fluvisols) under simulated flood conditions revealed by a metataxonomic approachss. Catena 2020, 188, 104448. [Google Scholar] [CrossRef]
- Mhuantong, W.; Wongwilaiwalin, S.; Laothanachareon, T.; Eurwilaichitr, L.; Tangphatsornruang, S.; Boonchayaanant, B.; Limpiyakorn, T.; Pattaragulwanit, K.; Punmatharith, T.; McEvoy, J. Survey of microbial diversity in flood areas during Thailand 2011 Flood Crisis using high-throughput tagged amplicon pyrosequencing. PLoS ONE 2015, 10, e0128043. [Google Scholar] [CrossRef] [PubMed]
- Delgado-Baquerizo, M. Obscure soil microbes and where to find them. ISME 2019, 13, 2120–2124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breidenbach, B.; Brenzinger, K.; Brandt, F.B.; Blaser, M.B.; Conrad, R. The effect of crop rotation between wetland rice and upland maize on the microbial communities associated with roots. Plant Soil 2017, 419, 435–445. [Google Scholar] [CrossRef] [Green Version]
- Francioli, D.; Schulz, E.; Lentendu, G.; Wubet, T.; Buscot, F.; Reitz, T. Mineral vs. organic amendments: Microbial community structure, activity and abundance of agriculturally relevant microbes are driven by long-term fertilization strategies. Front. Microbiol. 2016, 7, 1446. [Google Scholar] [CrossRef] [Green Version]
- Mori, K.; Yamaguchi, K.; Sakiyama, Y.; Urabe, T.; Suzuki, K. Caldisericum exile gen. nov., sp. nov., an anaerobic, thermophilic, filamentous bacterium of a novel bacterial phylum, Caldiserica phyl. nov., originally called the Candidate phylum op5, and description of Caldisericaceae fam. nov., Caldisericales ord. nov. and Caldisericia classis nov. Int. J. Syst. Evol. Microbiol. 2009, 59, 2894–2898. [Google Scholar]
- Poddar, N.; Badilla, C.; Maghool, S.; Osborne, T.H.; Santini, J.M.; Maher, M.J. Structural and functional investigation of the periplasmic arsenate-binding protein arrx from Chrysiogenes arsenatis. Biochemistry 2021, 60, 465–476. [Google Scholar] [CrossRef]
- Abdul Rahman, N.; Parks, D.H.; Vanwonterghem, I.; Morrison, M.; Tyson, G.W.; Hugenholtz, P. A Phylogenomic analysis of the bacterial phylum Fibrobacteres. Front. Microbiol. 2016, 6, 1469. [Google Scholar] [CrossRef] [Green Version]
- De Paula Silveira Mello, L.; dos Santos, A.C.; dos Santos, R.M.; Kandasamy, S.; Lazarovits, G.; Rigobelo, E.C. Application of the bacterial strains Ruminobacter amylophilus, Fibrobacter succinogenes and Enterococcus faecium for growth promotion in maize and soybean plants. Aust. J. Crop Sci. 2020, 14, 2020–2027. [Google Scholar] [CrossRef]
- Sharma, M.; Mishra, V.; Rau, N.; Sharma, R.S. Increased iron-stress resilience of maize through inoculation of siderophore-producing Arthrobacter globiformis from mine. J. Basic Microbiol. 2016, 56, 719–735. [Google Scholar] [CrossRef]
- Yadav, A.N.; Verma, P.; Kumar, S.; Kumar, V.; Kumar, M.; Sugitha, T.C.K.; Singh, B.P.; Saxena, A.K.; Dhaliwal, H.S. Actinobacteria from rhizosphere: Molecular diversity, distributions, and potential biotechnological applications. In New and Future Developments in Microbial Biotechnology and Bioengineering; Gupta, K., Ed.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 13–41. [Google Scholar]
- Baudoin, E.; Benizri, E.; Guckert, A. Impact of artificial root exudates on the bacterial community structure in bulk soil and maize rhizosphere. Soil Biol. Biochem. 2003, 35, 1183–1192. [Google Scholar] [CrossRef]
- Wolna-Maruwka, A.; Niewiadomska, A.; Klama, J. Biological activity of grey-brown podzolic soil organically fertilized for maize cultivation in monoculture. Pol. J. Environ. Stud. 2009, 18, 931–939. [Google Scholar]
- Tian, J.; Wang, J.; Dippold, M.; Gao, Y.; Blagodatskaya, E.; Kuzyakov, Y. Biochar affects soil organic matter cycling and microbial functions but does not alter microbial community structure in a paddy soil. Sci. Total Environ. 2016, 556, 89–97. [Google Scholar] [CrossRef]
- Elzobair, K.A.; Stromberger, M.E.; Ippolito, J.A.; Lentz, R.D. Contrasting effects of biochar versus manure on soil microbial communities and enzyme activities in an aridisol. Chemosphere 2016, 142, 145–152. [Google Scholar] [CrossRef]
- Chen, J.; Li, S.; Liang, C.; Xu, Q.; Li, Y.; Qin, H.; Fuhrmann, J.J. Response of microbial community structure and function to short-term biochar amendment in an intensively managed bamboo (Phyllostachys praecox) plantation soil: Effect of particle size and addition rate. Sci. Total Environ. 2017, 574, 24–33. [Google Scholar] [CrossRef]
- Li, G.L.; Zhou, C.H.; Fiore, S.; Yu, W.H. Interactions between microorganisms and clay minerals: New insights and broader applications. Appl. Clay Sci. 2019, 177, 91–113. [Google Scholar] [CrossRef]
- Gryndler, M. Interactions of arbuscular mycorrhizal fungi with other soil organisms. In Arbuscular mycorrhizas: Physiology and Function; Koltai, H., Kapulnik, Y., Eds.; Springer: Berlin/Heidelberg, Germany, 2000; pp. 239–262. [Google Scholar]
- Rangel-Castro, J.I.; Danell, E.; Pfeffer, P.E. A 13C-NMR study of exudation and storage of carbohydrates and amino acids in the ectomycorrhizal edible mushroom Cantharellus cibarius. Mycologia 2002, 94, 190–199. [Google Scholar]
- Landeweert, R.; Hoffland, E.; Finlay, R.D.; Kuyper, T.W.; van Breemen, N. Linking plants to rocks: Ectomycorrhizal fungi mobilize nutrients from minerals. Trends Ecol. Evol. 2001, 16, 248–254. [Google Scholar] [CrossRef]
- Olsson, P.A.; Chalot, M.; Baath, E.; Finlay, R.D.; Söderström, B. Ectomycorrhizal mycelia reduce bacterial activity in a sandy soil. FEMS Microbiol. Ecol. 1996, 21, 77–86. [Google Scholar] [CrossRef]
- Rillig, M.C. Arbuscular mycorrhizae and terrestrial ecosystem processes. Ecol. Lett. 2004, 7, 740–754. [Google Scholar] [CrossRef]
- Guangming, L.; Xuechen, Z.; Xiuping, W.; Hongbo, S.; Jingsong, Y.; Xiangping, W. Soil Enzymes as indicators of saline soil fertility under various soil amendments. Agric. Ecosyst. Environ. 2017, 237, 274–279. [Google Scholar] [CrossRef]
- Joergensen, R.G.; Wichern, F. Alive and kicking: Why dormant soil microorganisms matter. Soil Biol. Biochem. 2018, 116, 419–430. [Google Scholar] [CrossRef]
- Vázquez, M.M.; César, S.; Azcón, R.; Barea, J.M. Interactions between arbuscular mycorrhizal fungi and other microbial inoculants (Azospirillum, Pseudomonas, Trichoderma) and their effects on microbial population and enzyme activities in the rhizosphere of maize plants. Appl. Soil Ecol. 2000, 15, 261–272. [Google Scholar] [CrossRef]
- Wolna-Maruwka, A.; Piechota, T.; Dach, J.; Szczech, M.; Szczerbal, I.; Niewiadomska, A.; Budka, A.; Gaj, R. The Influence of Trichoderma on the phytosanitary status of soil and yield of red beets (Beta vulgaris L. subsp. vulgaris). Pol. J. Environ. Stud. 2017, 26, 847–859. [Google Scholar] [CrossRef]
- Niewiadomska, A.; Sulewska, H.; Wolna-Maruwka, A.; Klama, J. Effect of organic fertilization on development of proteolytic bacteria and activity of proteases in the soil for cultivation of maize (Zea mays L.). Arch. Environ. Prot. 2010, 36, 47–56. [Google Scholar]
- Hupe, A.; Schulz, H.; Bruns, C.; Haase, T.; Heß, J.; Joergensen, R.G.; Wichern, F. Even Flow? Changes of carbon and nitrogen release from pea roots over time. Plant Soil 2018, 431, 143–157. [Google Scholar] [CrossRef]
- Pińskwar, I.; Choryński, A.; Kundzewicz, Z.W. Severe drought in the spring of 2020 in Poland—More of the same? Agronomy 2020, 10, 1646. [Google Scholar] [CrossRef]
- Tomczyk, A.M.; Bednorz, E. The Extreme Year—Analysis of thermal conditions in Poland in 2018. Theor. Appl. Climatol. 2020, 139, 251–260. [Google Scholar] [CrossRef] [Green Version]
- Jalal, F.; Arif, M.; Akhtar, K.; Khan, A.; Naz, M.; Said, F.; Zaheer, S.; Hussain, S.; Imtiaz, M.; Khan, M.A. Biochar integration with legume crops in summer gape synergizes nitrogen use efficiency and enhance maize yield. Agronomy 2020, 10, 58. [Google Scholar] [CrossRef] [Green Version]
- Guo, M. The 3R principles for applying biochar to improve soil health. Soil Systems 2020, 4, 9. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Xie, W.; Chen, B. Arbuscular mycorrhiza improved drought tolerance of maize seedlings by altering Photosystem II efficiency and the levels of key metabolites. Chem. Biol. Technol. Agric. 2020, 7, 1–14. [Google Scholar] [CrossRef]
- Nyamwange, M.M.; Njeru, E.M.; Mucheru-Muna, M.; Ngetich, F. Soil management practices affect arbuscular mycorrhizal fungi propagules, root colonization and growth of rainfed maize. AIMS Agric. Food 2018, 3, 120–134. [Google Scholar] [CrossRef]
- Moitzi, G.; Thünauer, G.; Gronauer, A. Energy use and energy efficiency of corn production with different mineral fertilization in south Styria. In Proceedings of the International Conferenceof Agricultural Engineering, Zurich, Switzerland, 6–10 July 2014. [Google Scholar]
- Pfordt, A.; Ramos Romero, L.; Schiwek, S.; Karlovsky, P.; von Tiedemann, A. Impact of environmental conditions and agronomic practices on the prevalence of fusarium species associated with ear-and stalk rot in maize. Pathogens 2020, 9, 236. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
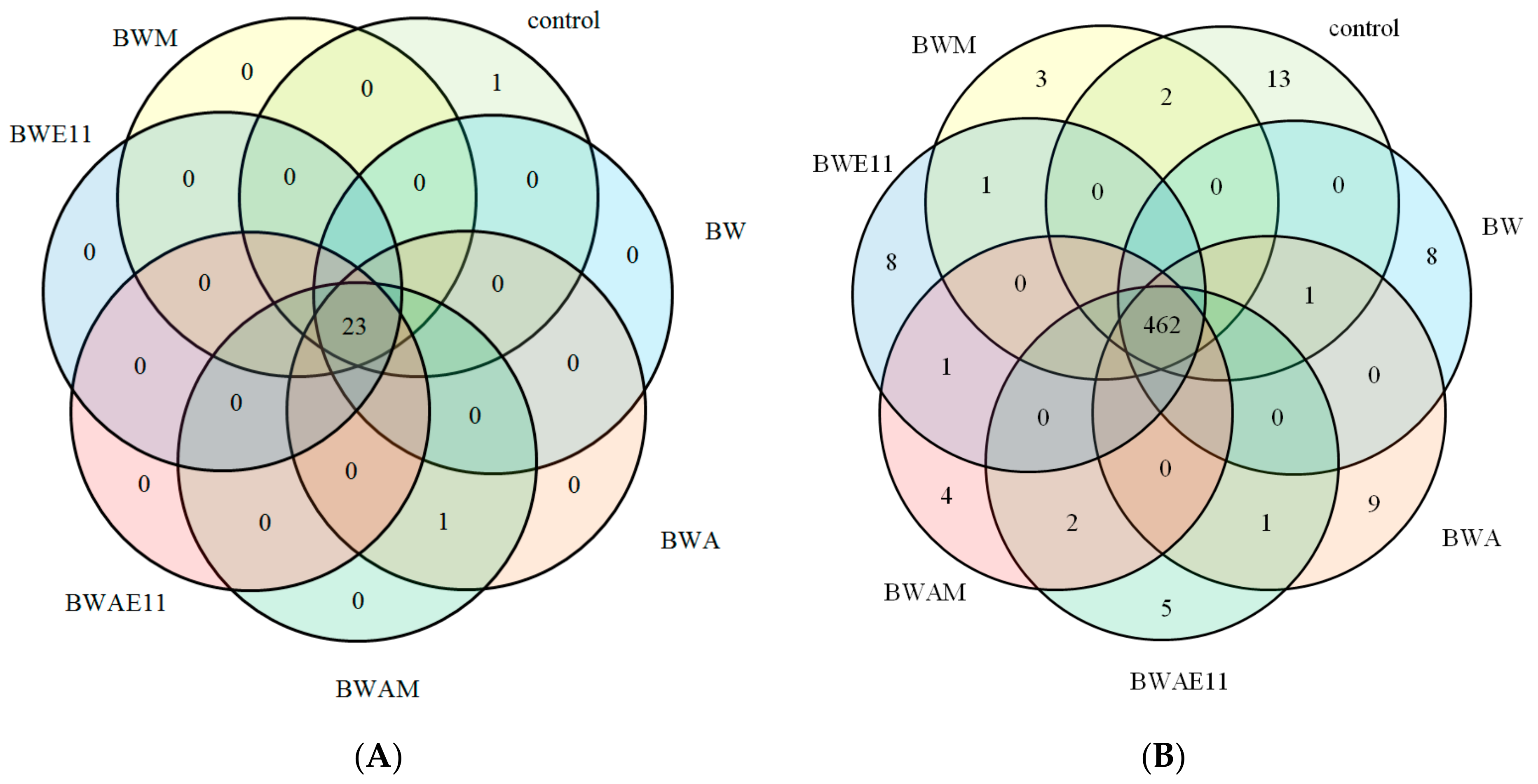{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Properties | Content |
|---|---|
| Organic C (g kg−1) | 5.61 |
| Total N (g kg−1) | 0.57 |
| Ca (g kg−1) | 2.61 |
| P (mg kg−1) | 50.8 |
| K (mg kg−1) | 11.85 |
| Mg (g kg−1) | 31.01 |
| pH KCl | 6.95 |
| Properties | Content |
|---|---|
| Organic C (g kg−1) | 580.11 |
| N (g kg−1) | 12.35 |
| P (g kg−1) | 50.8 |
| K (g kg−1) | 11.85 |
| Mg (g kg−1) | 31.01 |
| pH KCl | 6.78 |
| Variant | |||||||
|---|---|---|---|---|---|---|---|
| Control ‘0’ | Control | BW | BWM | BWE11 | BWA | BWAE11 | BWAM |
| Number of Unique Taxa | |||||||
| 8 | 13 | 8 | 3 | 8 | 9 | 5 | 4 |
| Dethiobacter | Bizionia | Candidatus Az. | Phormidium | Erwinia | Muricauda | Cellvibrio | Pseudochrobactrum |
| Alloscardovia | Butyrivibrio | Ignatzschineria | Anaerostipes | Anaerotruncus | Acidianus | Desulfobacterium | Elizabethkingia |
| Desulfosporomusa | Fibrobacter | Arsenicicoccus | Geovibrio | Mogibacterium | Anaeromusa | Nitrospina | Halanaerobacter |
| Desulfurococcus | Fusobacterium | Azorhizophilus | Planktothrix | Anaeroplasma | Ruminococcus | Herpetosiphon | |
| Providencia | Melissococcus | Candidatus Ph. | Dickeya | Martelella | Sediminibacterium | ||
| Salisaeta | Nevskia | Kurthia | Pseudanabaena | Natranaerobius | |||
| Trabulsiella | Nitrincola | Porphyromonas | Reinekea | Rothia | |||
| Vagococcus | Rapidithrix | Stenoxybacter | Verrucomicrobium | Succinivibrio | |||
| Sporanaerobacte | Trichococcus | ||||||
| Thiovirga | |||||||
| Alkanindiges | |||||||
| Citrobacter | |||||||
| Tepidimicrbium | |||||||
| Variants | Yield of Seeds (t ha−1) | Plant Density (No. m−2) | Number of Cobs per Plant | Number Of Seeds per Cob | Thousand Kernel Weight (g) | Moisture Content at Harvest (%) | Number of Cobs per m2 |
|---|---|---|---|---|---|---|---|
| control | 6.94 b (±1.36) | 6.65 a (± 0.26) | 0.87 a (±0.02) | 360 a (±60) | 281 a (±8) | 22.8 a (±1.6) | 5.8 a (±0.13) |
| BW | 6.98 ab (±1.09) | 7.20 a (±0.17) | 0.81 a (±0.04) | 352 a (±58) | 279 a (±4) | 19.3 ab (±0.5) | 5.8 a (±0.19) |
| BWM | 8.83 a (±1.39) | 7.10 a (±0.17) | 0.89 a (±0.01) | 437 a (±67) | 289 a (±9) | 18.0 b (±2.7) | 5.9 a (±0.12) |
| BWE11 | 7.40 ab (±1.42) | 6.73 a (±0.16) | 0.87 a (±0.02) | 385 a (±80) | 280 a (±5) | 22.4 a (±0.9) | 5.9 a (±0.13) |
| BWA | 7.89 ab (±1.32) | 6.42 a (±0.10) | 0.91 a (±0.02) | 429 a (±53) | 279 a (±16) | 21.5 ab (±1.2) | 5.8 a (±0.14) |
| BWAE11 | 7.46 ab (±1.32) | 7.15 a (±0.24) | 0.82 a (±0.02) | 386 a (±58) | 277 a (±12) | 20.0 ab (±1.1) | 5.8 a (±0.17) |
| BWAM | 8.02 ab (±1.25) | 6.85 a (±0.21) | 0.88 a (±0.03) | 407 a (±48) | 283 a (±14) | 21.3 ab (±0.7) | 6.0 a (±0.10) |
| LSD (0.05) | 1.88 | ns | ns | ns | Ns | 4.3 | ns |
| Variants | N Content (%) | Protein Content (%) | Starch Content (%) | Ash Content (%) | Stem Rot (%) | Ear Hanging (%) | Ear Rot (%) |
|---|---|---|---|---|---|---|---|
| control | 1.41 a | 8.84 a | 69.2 c | 1.25 ab | 0.0 a | 0.2 a | 0.0 a |
| BW | 1.42 a | 8.87 a | 70.5 abc | 1.24 ab | 0.0 a | 0.3 a | 0.0 a |
| BWM | 1.42 a | 8.96 a | 71.6 a | 1.18 b | 0.3 a | 0.0 a | 0.0 a |
| BWE11 | 1.44 a | 8.98 a | 69.7 bc | 1.27 a | 0.0 a | 0.2 a | 0.0 a |
| BWA | 1.42 a | 8.85 a | 70.5 abc | 1.25 ab | 0.3 a | 0.0 a | 0.0 a |
| BWAE11 | 1.42 a | 8.90 a | 70.4 abc | 1.24 ab | 0.0 a | 0.0 a | 0.0 a |
| BWAM | 1.38 a | 8.63 a | 71.2 ab | 1.21 ab | 0.0 a | 0.0 a | 0.0 a |
| LSD (0.05) | ns | ns | 1.6 | 0.07 | ns | ns | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wolna-Maruwka, A.; Piechota, T.; Niewiadomska, A.; Kamiński, A.; Kayzer, D.; Grzyb, A.; Pilarska, A.A. The Effect of Biochar-Based Organic Amendments on the Structure of Soil Bacterial Community and Yield of Maize (Zea mays L.). Agronomy 2021, 11, 1286. https://doi.org/10.3390/agronomy11071286
Wolna-Maruwka A, Piechota T, Niewiadomska A, Kamiński A, Kayzer D, Grzyb A, Pilarska AA. The Effect of Biochar-Based Organic Amendments on the Structure of Soil Bacterial Community and Yield of Maize (Zea mays L.). Agronomy. 2021; 11(7):1286. https://doi.org/10.3390/agronomy11071286
Chicago/Turabian StyleWolna-Maruwka, Agnieszka, Tomasz Piechota, Alicja Niewiadomska, Adam Kamiński, Dariusz Kayzer, Aleksandra Grzyb, and Agnieszka A. Pilarska. 2021. "The Effect of Biochar-Based Organic Amendments on the Structure of Soil Bacterial Community and Yield of Maize (Zea mays L.)" Agronomy 11, no. 7: 1286. https://doi.org/10.3390/agronomy11071286