
Growing Environment and Heat Treatment Effects on Intra- and Interspecific Pollination in Chile Pepper (Capsicum spp.)

Abstract
:1. Introduction
2. Materials and Methods
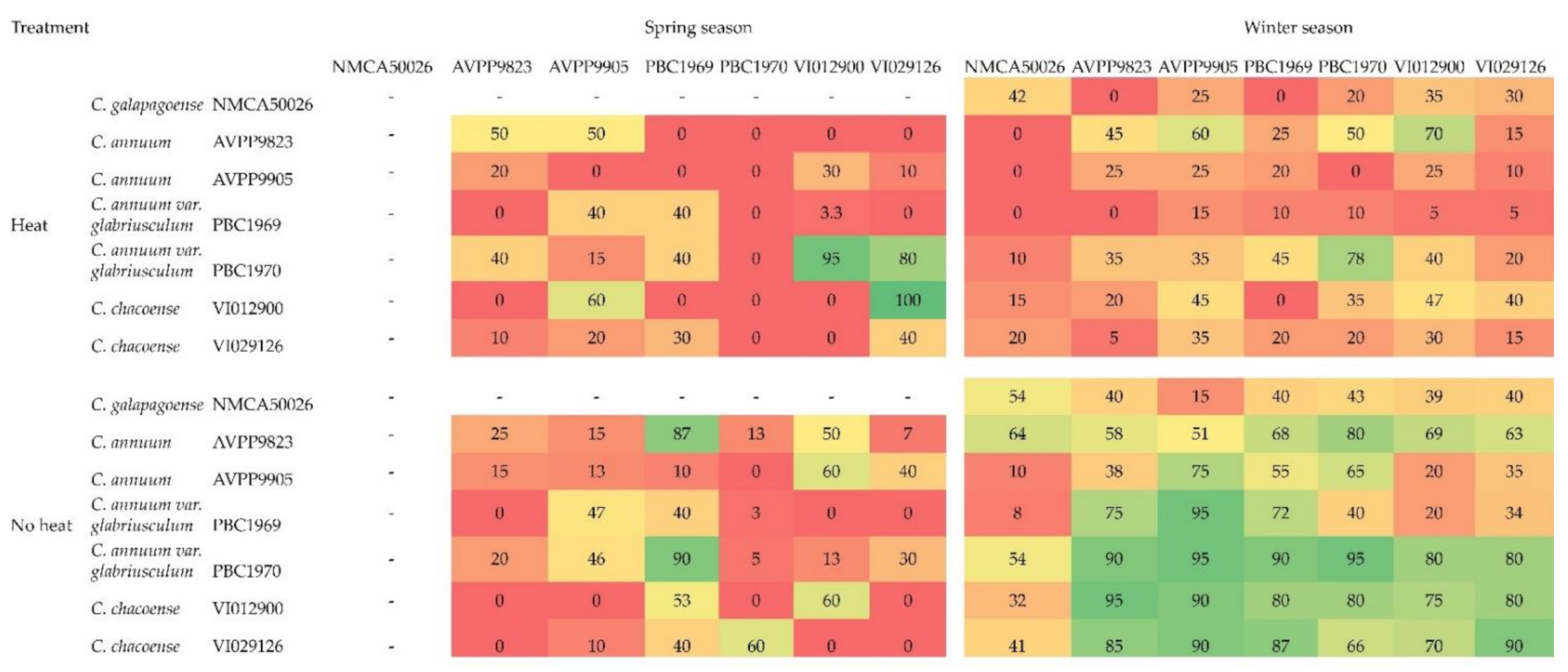
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Food and Agriculture Organization of the United Nations. FAOSTAT Statistical Database. 2019. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 22 January 2020).
- Barchenger, D.W.; Naresh, P.; Kumar, S. The Capsicum Genome; Ramchiary, N., Kole, C., Eds.; Springer Nature Switzerland AG: Cham, Switzerland, 2019; Chapter 2; pp. 9–23. [Google Scholar]
- Sanders, D.C.; Kirk, H.J.; van der Brink, C. Growing Degree Days in North Carolina; North Carolina Agricultural Extension Service: Sparta, NC, USA, 1980. [Google Scholar]
- Hedhly, A.; Hormaza, J.I.; Herrero, M. Global warming and sexual plant reproduction. Trends Plant Sci. 2009, 14, 30–36. [Google Scholar] [CrossRef]
- Giorno, F.; Wolters-Arts, M.; Mariani, C.; Rieu, I. Ensuring reproduction at high temperatures: The heat stress response during anther and pollen development. Plants 2013, 2, 489–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Driedonks, N.; Wolters-Arts, M.; Huber, H.; de Boer, G.J.; Vriezen, W.; Mariani, C.; Rieu, I. Exploring the natural variation for reproductive thermotolerance in wild tomato species. Euphytica 2018, 214, 67. [Google Scholar] [CrossRef] [Green Version]
- Barchenger, D.W.; Bosland, P.W. Cultivated Wild Relatives of North America; Greene, S., Williams, K., Khoury, C.K., Kantar, M.B., Marek, L., Eds.; Springer Nature Switzerland AG: Cham, Switerzland, 2019; Volume 2, Chapter 7; pp. 225–242. [Google Scholar]
- Khoury, C.K.; Carver, D.; Barchenger, D.W.; Barboza, G.; van Zonneveld, M.; Jarret, R.; Bohs, L.; Kantar, M.; Uchanski, M.; Mercer, K.; et al. Modeled distributions and conservation status of the wild relatives of chile peppers (Capsicum L.). Divers. Distrib. 2020, 26, 209–225. [Google Scholar] [CrossRef] [Green Version]
- Lin, T.; Zhu, G.; Zhang, J.; Xu, X.; Yu, Q.; Zheng, Z.; Zhang, Z.; Lun, Y.; Li, S.; Wang, X.; et al. Genomic analyses provide insights into the history of tomato breeding. Nat. Genet. 2014, 46, 1220–1226. [Google Scholar] [CrossRef] [PubMed]
- Hirsch, C.N.; Hirsch, C.D.; Felcher, K.; Coombs, J.; Zarka, D.; van Deynze, A.; de Jong, W.; Veilleux, R.E.; Jansky, S.; Bethke, P.; et al. Retrospective view of North American potato (Solanum tuberosum L.) breeding in the 20th and 21st centuries. G3 Genes Genom Genet. 2013, 3, 1003–1013. [Google Scholar] [CrossRef] [Green Version]
- Aguilar Meléndez, A.; Morrell, P.L.; Roose, M.L.; Kim, S.C. Genetic diversity and structure in semiwild and domesticated chiles (Capsicum annuum; Solanaceae) from Mexico. Am. J. Bot. 2009, 96, 1190–1202. [Google Scholar] [CrossRef] [PubMed]
- Carrizo García, C.; Barfuss, M.H.J.; Sehr, E.M.; Barboza, G.E.; Samuel, R.; Moscone, E.A.; Ehrendorfer, F. Phylogenetic relationships, diversification and expansion of chili peppers (Capsicum, Solanaceae). Ann. Bot. 2016, 118, 35–51. [Google Scholar] [CrossRef] [Green Version]
- Cheng, J.; Qin, C.; Tang, X.; Zhou, H.; Hu, Y.; Zhao, Z.; Cui, J.; Li, B.; Wu, Z.; Yu, J.; et al. Development of a SNP array and its application to genetic mapping and diversity assessment in pepper (Capsicum spp.). Sci. Rep. 2016, 6, 33293. [Google Scholar] [CrossRef]
- Oyama, K.; Hernández-Verdugo, S.; Sánchez, C.; González-Rodríguez, A.; Sánchez-Peña, P.; Garzón-Tiznado, J.A.; Casas, A. Genetic structure of wild and domesticated populations of Capsicum annuum (Solanaceae) from northwestern Mexico analyzed by RAPDs. Genet. Resour. Crop Evol. 2006, 53, 553–562. [Google Scholar] [CrossRef]
- Votava, E.J.; Nabhan, G.P.; Bosland, P.W. Genetic diversity and similarity revealed via molecular analysis among and within an in situ population and ex situ accessions of chiltepín (Capsicum annuum var. glabriusculum). Conserv. Genet. 2002, 3, 123–129. [Google Scholar] [CrossRef]
- Parry, C.; Wang, Y.W.; Lin, S.W.; Barchenger, D.W. Reproductive compatibility in Capsicum is not necessarily reflected in genetic or phenotypic similarity between species complexes. PLoS ONE 2021, 16, e0243689. [Google Scholar] [CrossRef]
- Reddy, K.R.; Kakani, V.G. Screening Capsicum species of different origins for high temperature tolerance by in vitro pollen germination and pollen tube length. Sci. Hort. 2007, 112, 130–135. [Google Scholar] [CrossRef]
- Heidmann, I.; Schade-Kampmann, G.; Lambalk, J.; Ottiger, M.; Di Berardino, M. Impedance flow cytometry: A novel technique in pollen analysis. PLoS ONE 2016, 11, e0165531. [Google Scholar] [CrossRef] [PubMed]
- Kenyon, L.; Hanson, P.; Kumar, S.; Shih, S.L.; Hsieh, M.H.; Chen, H.Y.; Lu, S.F.; Wang, Y.Y.; Lin, S.W.; Cheng, Y.H. Treatment for Cleaning Small Seed lots of Tomato and Pepper Seeds of Surface Contamination with Viroids. The World Vegetable Center. 2017. Available online: https://worldveg.tind.io/record/74053?ln=en (accessed on 19 December 2020).
- Berke, T.; Black, L.L.; Talekar, N.S.; Wang, J.F.; Gniffke, P.; Green, S.K.; Wang, T.C.; Morris, R. Suggested Cultural Practices for Chili Pepper; World Vegetable Center: Tainan, Taiwan, 2005. [Google Scholar]
- Lin, S.W.; Lin, T.H.; Wang, Y.W.; Barchenger, D.W. High throughput pollen phenotyping using impedance flow cytometry for heat tolerance selection in pepper (Capsicum sp.). HortScience 2020, 55, S156. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 17 February 2021).
- Aloni, B.; Peet, M.M.; Pharr, M.; Karni, L. The effect of high temperature and high atmospheric CO2 on carbohydrate changes in bell pepper (Capsicum annuum) pollen in relation to its germination. Physiol. Plant. 2001, 112, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Erickson, A.N.; Markhart, A.H. Flower developmental stage and organ sensitivity of bell pepper (Capsicum annuum L.) to elevated temperature. Plant Cell Environ. 2002, 25, 123–130. [Google Scholar] [CrossRef]
- Guo, M.; Yin, Y.X.; Ji, J.J.; Ma, B.P.; Lu, M.H.; Gong, Z.H. Cloning and expression analysis of heat-shock transcription factor gene CaHsfA1 from pepper (Capsicum annuum L.). Genetic. Mol. Res. 2014, 13, 1865–1875. [Google Scholar] [CrossRef]
- Polowick, P.L.; Sawhney, V.K. Temperature effects on male fertility and flower and fruit development in Capsicum annuum L. Sci. Hort. 1985, 25, 117–127. [Google Scholar] [CrossRef]
- Pressman, E.; Peet, M.M.; Pharr, D.M. The effect of heat stress on tomato pollen characteristics is associated with changes in carbohydrates concentration in developing anthers. Ann. Bot. 2002, 90, 631–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wubs, A.M.; Heuvelink, E.; Marcelis, L.F.M. Abortion of reproductive organs in sweet pepper (Capsicum annuum L.): A review. J. Hortic. Sci. Biotech. 2009, 84, 467–475. [Google Scholar] [CrossRef]
- Müller, F.; Rieu, I. Acclimation to high temperature during pollen development. Plant Reproduct. 2016, 29, 107–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotak, S.; Larkindale, J.; Lee, U.; von Koskull-Doring, P.; Vierling, E.; Scharf, K.D. Complexity of the heat stress response in plants. Curr. Opin. Plant Biol. 2007, 10, 310–316. [Google Scholar] [CrossRef]
- Firon, N.; Shaked, R.; Preet, M.M.; Pharr, D.M.; Zamski, E.; Rosenfeld, K.; Althan, L.; Pressman, E. Pollen grains of heat tolerant tomato cultivars retain higher carbohydrate concentration under heat stress conditions. Sci. Hortic. 2006, 109, 212–217. [Google Scholar] [CrossRef]
- Gajanayake, B.; Trader, B.W.; Raja Reddy, K.; Harkess, R.L. Screening ornamental pepper cultivars for temperature tolerance using pollen and physiological parameters. HortScience 2011, 46, 878–884. [Google Scholar] [CrossRef] [Green Version]
- Salem, M.A.; Kakani, V.G.; Koti, S.; Reddy, K.R. Pollen-based screening of soybean genotypes for high temperatures. Crop Sci. 2007, 47, 219–231. [Google Scholar] [CrossRef] [Green Version]
- Usman, M.G.; Radii, M.Y.; Ismail, M.R.; Malek, M.A.; Latif, M.A. Expression of target gene Hsp70 and membrane stability heat tolernace in chili pepper. J. Am. Soc. Hortic. Sci. 2015, 140, 144–150. [Google Scholar] [CrossRef] [Green Version]
- Costa, L.V.; Lopes, R.; Lopes, M.T.G.; de Figueiredo, A.F.; Barros, W.S.; Alves, R.R.M. Cross compatibility of domesticated hot pepper and cultivated sweet pepper. Crop Breed. Appl. Biotechnol. 2009, 9, 37–44. [Google Scholar] [CrossRef]
- da Silva Montero, C.E.; Santana Pereira, T.N.; de Campos, K.P. Reproductive characterization of interspecific hybrids among Capsicum species. Crop Breed. Appl. Biotechnol. 2011, 11, 3. [Google Scholar] [CrossRef]
- Pickersgill, B. Relationships between weedy and cultivated forms in some species of chili peppers (genus Capsicum). Evolution 1971, 25, 683–691. [Google Scholar] [CrossRef]
- Tong, N.; Bosland, P.W. Capsicum tovarii, a new member of the Capsicum baccatum complex. Euphytica 1999, 109, 71–77. [Google Scholar] [CrossRef]
- Li, T.; Xu, X.; Li, Y.; Wang, H.; Li, Z.; Li, Z. Comparative transcriptome analysis reveals differential transcription in heat-susceptible and heat-tolerant pepper (Capsicum annum L.) cultivars under heat stress. J. Plant Biol. 2015, 58, 411–424. [Google Scholar] [CrossRef]
- Zhai, Y.; Guo, M.; Wang, H.; Lu, J.; Liu, J.; Zhang, C.; Gong, Z.; Lu, M. Autophagy, a conserved mechanism for protein degradation, responds to heat, and other abiotic stresses in Capsicum annuum L. Front. Plant Sci. 2016, 7, 131. [Google Scholar] [CrossRef] [Green Version]
- Feng, X.H.; Zhang, H.X.; Ali, M.; Gai, W.X.; Cheng, G.X.; Yu, Q.H.; Yang, S.B.; Li, X.X.; Gong, Z.H. A small heat shock protein CaHsp25.9 positively regulates heat, salt, and drought stress tolerance in pepper (Capsicum annuum L.). Plant Physiol. Biochem. 2019, 142, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.; Muhammad, I.; Haq, S.U.; Alam, M.; Khattak, A.M.; Akhtar, K.; Ullah, H.; Khan, A.; Lu, G.; Gong, Z.H. The CaChiVI2 gene of Capsicum annuum L. confers resistance against heat stress and infection of Phytophthora capsici. Front. Plant Sci. 2020, 11, 219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajametov, S.; Yang, E.Y.; Cho, M.C.; Chae, S.Y.; Jeong, H.B. Morpho-physiological pesponse to heat stress during seedling stage and interrelation with agronomical traits in pepper (Capsicum annuum L.) cultivars differing in heat tolerance. Res. Sq. 2021. [Google Scholar] [CrossRef]
- Usman, I.S.; Mamat, A.S.; Syed Mohd, H.S.Z.; Aishan, H.S.; Anuar, R.A. The non-impairment of pollination and fertilization in the abscission of chilli (Capsicum annuum L. Var. Kulai) flowers under high temperature and humid conditions. Sci. Hortic. 1999, 79, 1–11. [Google Scholar] [CrossRef]
- Motamedi, M.; Haghighi, M.; Goli, A. Physiological changes of sweet and hot peppers in vegetative and reproductive growth stages treated by Ca and H2O2 under unforeseen heat stresses. Sci. Hortic. 2018, 249, 306–313. [Google Scholar] [CrossRef]


{kind=link}
| Season | Month | Plant Growth Stage | Mean Temperature (°C) | Maximum Temperature (°C) | Minimum Temperature (°C) | Mean RH (%) 1 | Maximum RH (%) | Minimum RH (%) |
|---|---|---|---|---|---|---|---|---|
| Summer | 4 | Vegetative | 26.8 | 41 | 17.5 | 77.7 | 100 | 33.8 |
| 5 | Vegetative | 27.7 | 43.8 | 19.8 | 83.2 | 100 | 38.7 | |
| 6 | Reproductive | 30 | 42.7 | 22.9 | 81.6 | 100 | 35.1 | |
| 7 | Reproductive | 29.5 | 43.9 | 24.1 | 86.1 | 100 | 42.4 | |
| 8 | Reproductive | 28.7 | 42.9 | 23.9 | 90.4 | 100 | 42.3 | |
| Winter | 10 | Vegetative | 27.5 | 44.3 | 19.4 | 70.2 | 100 | 30.3 |
| 11 | Vegetative | 24.8 | 41.4 | 14.4 | 74 | 100 | 29.4 | |
| 12 | Reproductive | 20.6 | 36.9 | 9.7 | 81.4 | 100 | 30 | |
| 1 | Reproductive | 19.9 | 38.3 | 5.4 | 80.5 | 100 | 24.8 | |
| 2 | Reproductive | 20.9 | 39.2 | 7.9 | 76.6 | 100 | 28.6 |
| Factor | DF 1 | Mean Squares | F-Value | p-Value |
|---|---|---|---|---|
| Season | 1 | 6.2 | 11.8 | 0.001 |
| Treatment | 1 | 46.9 | 90.2 | <0.001 |
| Female accession | 6 | 4.1 | 7.1 | <0.001 |
| Male accession | 6 | 1.7 | 2.9 | 0.009 |
| Female species | 3 | 0.5 | 0.7 | 0.559 |
| Male species | 3 | 2.6 | 4.3 | 0.006 |
| Season × treatment | 1 | 4.3 | 8.3 | 0.004 |
| Female × male | 36 | 0.8 | 1.3 | 0.106 |
| Female sp. ×male sp. | 9 | 1.7 | 2.7 | 0.005 |
| Replication(treatment) | 6 | 3.7 | 9.1 | <0.001 |
| Residuals | 406 | 0.4 |
| Summer Season | Winter Season | Heat Treatment | No Heat Treatment | |
|---|---|---|---|---|
| Summer season | 0.122 | 0.005 | 0.031 | |
| Winter season | 0.26 | <0.001 | <0.001 | |
| Heat treatment | 0.46 | 0.77 | 0.005 | |
| No-heat treatment | 0.36 | 0.86 | 0.39 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, T.H.; Lin, S.W.; Wang, Y.W.; van Zonneveld, M.; Barchenger, D.W. Growing Environment and Heat Treatment Effects on Intra- and Interspecific Pollination in Chile Pepper (Capsicum spp.). Agronomy 2021, 11, 1275. https://doi.org/10.3390/agronomy11071275
Lin TH, Lin SW, Wang YW, van Zonneveld M, Barchenger DW. Growing Environment and Heat Treatment Effects on Intra- and Interspecific Pollination in Chile Pepper (Capsicum spp.). Agronomy. 2021; 11(7):1275. https://doi.org/10.3390/agronomy11071275
Chicago/Turabian StyleLin, Tsung Han, Shih Wen Lin, Yen Wei Wang, Maarten van Zonneveld, and Derek W. Barchenger. 2021. "Growing Environment and Heat Treatment Effects on Intra- and Interspecific Pollination in Chile Pepper (Capsicum spp.)" Agronomy 11, no. 7: 1275. https://doi.org/10.3390/agronomy11071275