
Phytohormonal and Transcriptomic Response of Hulless Barley Leaf in Response to Powdery Mildew Infection

Abstract
:1. Introduction
2. Results
2.1. Phytohormonal Response of XL19 and ZYM1288
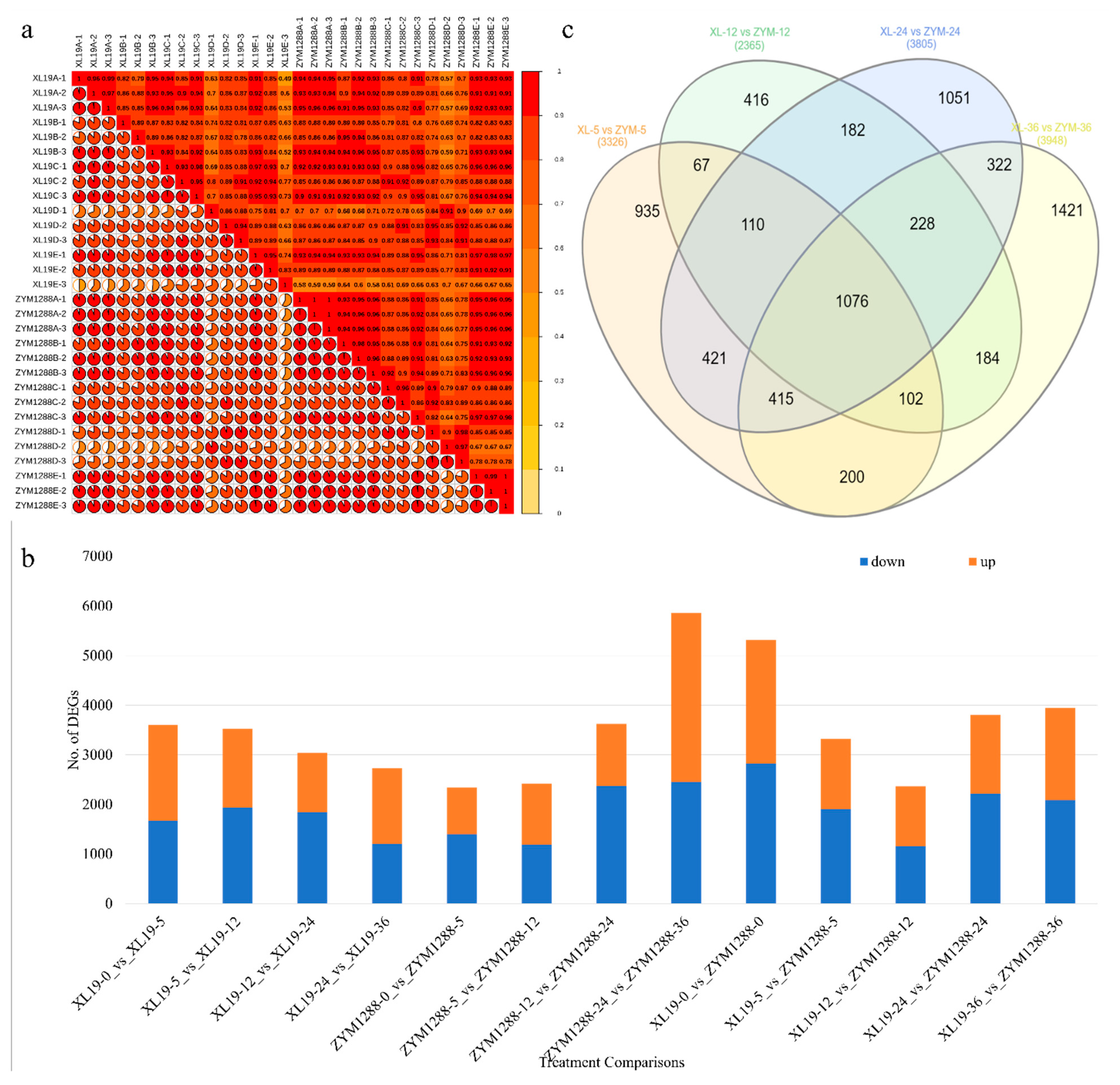
2.2. Transcriptomic Responses of XL19 and ZYM1288
2.3. Differential Gene Expression between XL19 and XYM1288
2.3.1. Signaling Responses
2.3.2. Transcriptional Changes in Energy Related Pathways
2.3.3. Transcriptional Changes in Defense Related Pathways
2.3.4. Transcriptional Changes in Known PM Related Genes
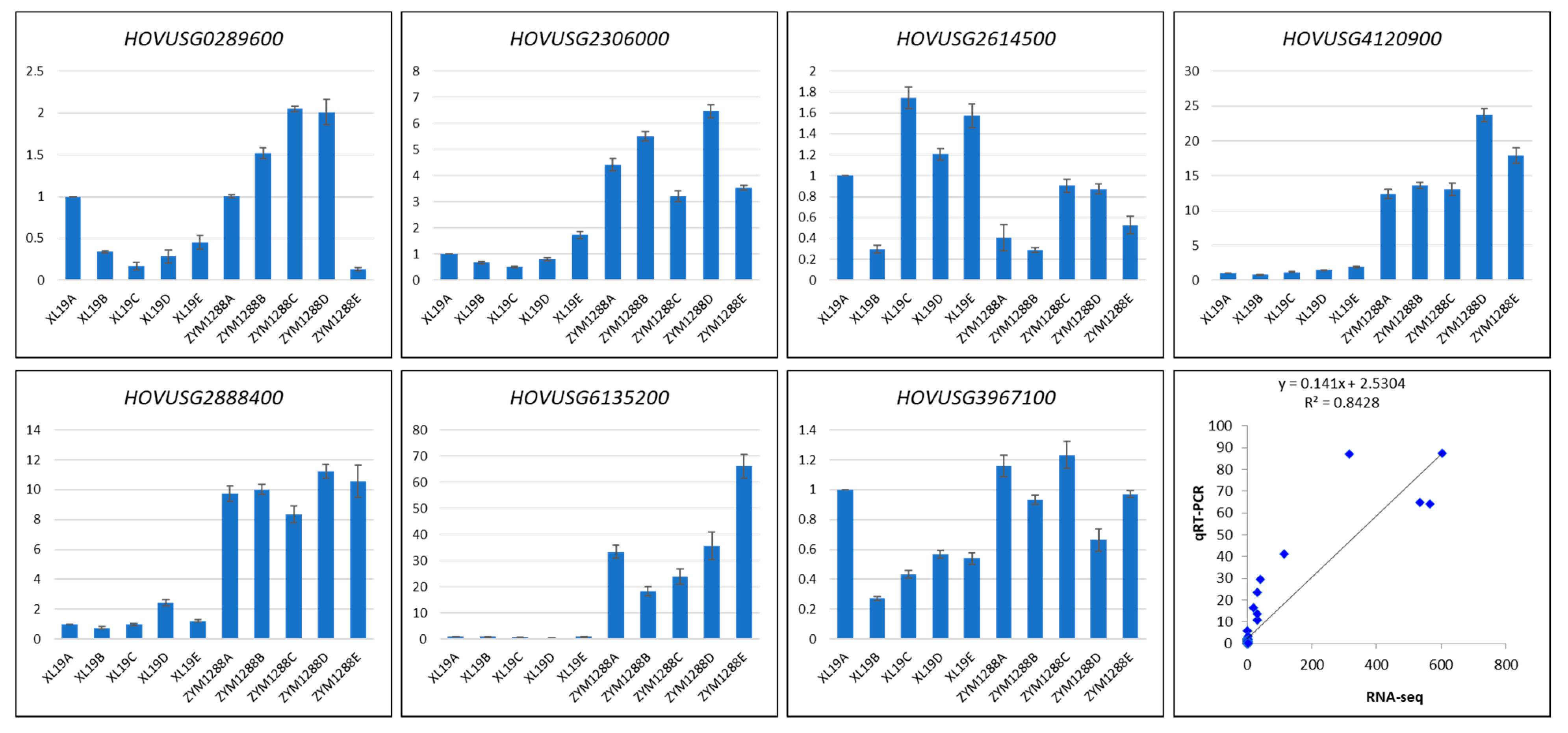
2.3.5. qRT-PCR Analysis
3. Discussion
3.1. Phytohormone Level and Signaling Is Differentially Affected in XL19 and ZYM1288
3.2. MAPK Signaling Pathway and Toll-Like Receptor Signaling Pathways Are Activated in Response to PM in Hulless Barley
3.3. PM Infection Modulates Changes in Energy Related Pathways in Hulless Barley
3.4. PM Triggers Changes in Defense-Associated Pathways in Hulless Barley
3.5. XL19 and ZYM1288 Differ in the Expression of Known PM Resistance Loci/Genes
4. Conclusions
5. Material and Method
5.1. Plant Materials, Growth Conditions, and Inoculation
5.2. Phytohormone Detection
5.3. Transcriptome Sequencing, Data Analyses, Differential Gene Expression, and Enrichment Analyses
5.4. qRT-PCR Analysis of Selected DEGs
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ABA | Abscisic acid |
| ABI5 | ABA-insensitive 5 |
| ALDOA | Fructose-bisphosphate aldolases |
| APXs | L-ascorbate peroxidases |
| AST | Aspartate aminotransferase |
| AUX | Auxin induced proteins |
| Bgh | Blumeria graminis (DC.) Golovin ex Speer f. sp. hordei Marchal |
| CDPK | Calcium Depedent Protein Kinase |
| COG | Clusters of Orthologous Groups |
| DEGs | Differentially expressed genes |
| ETI | Effector triggered immunity |
| FPKM | Fragments Per Kilobase of Transcript per Million fragments mapped |
| GGCTs | Gamma-glutamylcyclotransferase-plant |
| GO | Gene Ontology |
| GSS | Glutathione synthase |
| GSTs | Glutathione s-transferases |
| H2JA | Dihydrojasmonic acid |
| HK | Histidine kinases |
| HR | Hypersensitive response |
| IAA | Indole 3-acetic acid |
| ICA | Indole-3-carboxylic acid |
| ICA-Id | Indole-3-carboxaldehyde |
| IDH | Isocitrate dehydrogenase |
| iPPase | inorganic pyrophosphatases |
| IRAK | Interleukin-1 receptor-associated kinases |
| JA | Jasmonic acid |
| JA-ILE | Jasmonoyl-L-Isoleucine |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| KOG | Eukaryotic Orthologous Groups of proteins |
| LAPs | Leucyl aminopeptidases |
| MAPK | Mitogen-activated protein kinase |
| MDH | Malate dehydrogenase |
| MEIAA | Methylindole-3-acetic acid |
| MEJA | Methyl Jasmonate |
| Mlo | MILDEW RESISTANCE LOCUS O |
| MRPs | Multidrug resistance proteins |
| ORRs | Two-component response regulations |
| PAMP | Pathogen-associated molecular patterns |
| PCC | Pearson Correlation Coefficient |
| PEN | PENETRATION proteins |
| 6PGD | 6-phosphogluconate dehydrogenase |
| PEP | Phosphoenolpyruvate carboxylase |
| PHP5 | Phosphotransfer protein 5 |
| PLSC | 1-acyl-sn-glycerol-3-phosphate acyltransferase |
| PM | Powdery mildew |
| PP2A | Serine/threonine-protein phosphatase 2A activator |
| PRMC | Release factor glutamine methyltransferase |
| PS | Photosystem |
| PTI | PAMP-triggered immunity |
| RGA | Disease resistance gene analogs |
| RRM1s | Ribonucleoside-diphosphate reductase subunit M1 |
| RRM2 | cold-inducible RNA-binding protein |
| SA | Salicylic acid |
| SGT1 | Suppressor of the G2 allele of skp1 |
| SKP1 | S-phase kinase-associated protein 1 |
| SYP | SYNTAXIN OF PLANTS |
| T9L24.4 | Steroid 22-alpha-hydroxylase |
| TUBGCP6 | Gamma-tubulin complex component 6 |
| tZ | trans-zeatin |
References
- Zha, S.; Yang, C.; Zeng, X.; Li, Z.; Wang, Y.; Yuan, H.; Yu, M.; Xu, Q. Comparative analysis of H3K4 and H3K27 trimethylations in two contrasting Tibetan hulless barely varieties on powdery mildew infection. J. Plant Pathol. 2021, 103, 117–126. [Google Scholar] [CrossRef]
- Yuan, H.; Zeng, X.; Yang, Q.; Xu, Q.; Wang, Y.; Jabu, D.; Sang, Z.; Tashi, N. Gene coexpression network analysis combined with metabonomics reveals the resistance responses to powdery mildew in Tibetan hulless barley. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dreiseitl, A.; Yang, J. Powdery mildew resistance in a collection of Chinese barley varieties. Genet. Resour. Crop Evol. 2007, 54, 259–266. [Google Scholar] [CrossRef]
- Cao, X.-R.; Yao, D.-M.; Duan, X.-Y.; Wei, L.; Fan, J.-R.; Ding, K.-J.; Zhou, Y.-L. Effects of powdery mildew on 1000-kernel weight, crude protein content and yield of winter wheat in three consecutive growing seasons. J. Integr. Agric. 2014, 13, 1530–1537. [Google Scholar] [CrossRef]
- Zhou, M. Barley production and consumption. In Genetics and Improvement of Barley Malt Quality; Springer: Berlin/Heidelberg, Germany, 2009; pp. 1–17. [Google Scholar]
- Ridout, C.J.; Skamnioti, P.; Porritt, O.; Sacristan, S.; Jones, J.D.; Brown, J.K. Multiple avirulence paralogues in cereal powdery mildew fungi may contribute to parasite fitness and defeat of plant resistance. Plant Cell 2006, 18, 2402–2414. [Google Scholar] [CrossRef]
- Ramos, S.M.B.; Almeida, E.F.A.; Rocha, F.d.S.; Fernandes, M.d.F.G.; Santos, E.B.d. Organic fertilization and alternative products in the control of powdery mildew. Ornam. Hortic. 2020, 26, 57–68. [Google Scholar] [CrossRef]
- Zubrod, J.P.; Bundschuh, M.; Arts, G.; Brühl, C.A.; Imfeld, G.; Knäbel, A.; Payraudeau, S.; Rasmussen, J.J.; Rohr, J.; Scharmüller, A. Fungicides: An overlooked pesticide class? Environ. Sci. Technol. 2019, 53, 3347–3365. [Google Scholar] [CrossRef]
- Yang, F.; Yang, T.; Liu, K.; Yang, Q.; Wan, Y.; Wang, R.; Li, G. Analysis of Metabolite Accumulation Related to Pod Color Variation of Caragana intermedia. Molecules 2019, 24, 717. [Google Scholar] [CrossRef] [Green Version]
- Jones, J.D.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chelkowski, J.; Tyrka, M.; Sobkiewicz, A. Resistance genes in barley (Hordeum vulgare L.) and their identification with molecular markers. J. Appl. Genet. 2003, 44, 291–310. [Google Scholar]
- Piechota, U.; Czembor, P.C.; Słowacki, P.; Czembor, J.H. Identifying a novel powdery mildew resistance gene in a barley landrace from Morocco. J. Appl. Genet. 2019, 60, 243–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, X.; Kracher, B.; Saur, I.M.; Bauer, S.; Ellwood, S.R.; Wise, R.; Yaeno, T.; Maekawa, T.; Schulze-Lefert, P. Allelic barley MLA immune receptors recognize sequence-unrelated avirulence effectors of the powdery mildew pathogen. Proc. Natl. Acad. Sci. USA 2016, 113, E6486–E6495. [Google Scholar] [CrossRef] [Green Version]
- Ge, X.; Deng, W.; Lee, Z.Z.; Lopez-Ruiz, F.J.; Schweizer, P.; Ellwood, S.R. Tempered mlo broad-spectrum resistance to barley powdery mildew in an Ethiopian landrace. Sci. Rep. 2016, 6, 29558. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, H.; Lorek, J.; Kwaaitaal, M.; Consonni, C.; Becker, K.; Micali, C.; Ver Loren van Themaat, E.; Bednarek, P.; Raaymakers, T.M.; Appiano, M. Key components of different plant defense pathways are dispensable for powdery mildew resistance of the Arabidopsis mlo2 mlo6 mlo12 triple mutant. Front. Plant Sci. 2017, 8, 1006. [Google Scholar] [CrossRef] [PubMed]
- Jia, M.; Xu, H.; Liu, C.; Mao, R.; Li, H.; Liu, J.; Du, W.; Wang, W.; Zhang, X.; Han, R. Characterization of the powdery mildew resistance gene in the elite wheat cultivar Jimai 23 and its application in marker-assisted selection. Front. Genet. 2020, 11, 241. [Google Scholar] [CrossRef]
- Li, H.; Dong, Z.; Ma, C.; Tian, X.; Xiang, Z.; Xia, Q.; Ma, P.; Liu, W. Discovery of powdery mildew resistance gene candidates from Aegilops biuncialis chromosome 2Mb based on transcriptome sequencing. PLoS ONE 2019, 14, e0220089. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Guo, G.; Zhou, L.; Chen, Y.; Zong, Y.; Huang, J.; Lu, R.; Liu, C. Transcriptome Analysis Identifies Candidate Genes and Functional Pathways Controlling the Response of Two Contrasting Barley Varieties to Powdery Mildew Infection. Int. J. Mol. Sci. 2020, 21, 151. [Google Scholar] [CrossRef] [Green Version]
- Xin, M.; Wang, X.; Peng, H.; Yao, Y.; Xie, C.; Han, Y.; Ni, Z.; Sun, Q. Transcriptome comparison of susceptible and resistant wheat in response to powdery mildew infection. Genom. Proteom. Bioinform. 2012, 10, 94–106. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Q.; Gao, P.; Wan, Y.; Cui, H.; Fan, C.; Liu, S.; Luan, F. Comparative transcriptome profiling of genes and pathways related to resistance against powdery mildew in two contrasting melon genotypes. Sci. Hortic. 2018, 227, 169–180. [Google Scholar] [CrossRef]
- Saja, D.; Janeczko, A.; Barna, B.; Skoczowski, A.; Dziurka, M.; Kornaś, A.; Gullner, G. Powdery Mildew-Induced Hormonal and Photosynthetic Changes in Barley Near Isogenic Lines Carrying Various Resistant Genes. Int. J. Mol. Sci. 2020, 21, 4536. [Google Scholar] [CrossRef]
- Zeng, X.; Xu, T.; Ling, Z.; Wang, Y.; Li, X.; Xu, S.; Xu, Q.; Zha, S.; Qimei, W.; Basang, Y.; et al. An improved high-quality genome assembly and annotation of Tibetan hulless barley. Sci. Data 2020, 7, 1–9. [Google Scholar] [CrossRef]
- Guo, W.-L.; Chen, B.-H.; Chen, X.-J.; Guo, Y.-Y.; Yang, H.-L.; Li, X.-Z.; Wang, G.-Y. Transcriptome profiling of pumpkin (Cucurbita moschata Duch.) leaves infected with powdery mildew. PLoS ONE 2018, 13, e0190175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bari, R.; Jones, J.D. Role of plant hormones in plant defence responses. Plant Mol. Biol. 2009, 69, 473–488. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Zhang, L.-L.; Yang, X.-M.; Cao, X.-L.; Wang, Y.-G.; Ma, X.-F.; Chandran, V.; Fan, J.; Yang, H.; Shang, J. Transcriptome analysis reveals pathways facilitating the growth of tobacco powdery mildew in Arabidopsis. Phytopathol. Res. 2019, 1, 7. [Google Scholar] [CrossRef] [Green Version]
- Tian, X.; Zhang, L.; Feng, S.; Zhao, Z.; Wang, X.; Gao, H. Transcriptome analysis of apple leaves in response to powdery mildew (Podosphaera leucotricha) infection. Int. J. Mol. Sci. 2019, 20, 2326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fung, R.W.; Gonzalo, M.; Fekete, C.; Kovacs, L.G.; He, Y.; Marsh, E.; McIntyre, L.M.; Schachtman, D.P.; Qiu, W. Powdery mildew induces defense-oriented reprogramming of the transcriptome in a susceptible but not in a resistant grapevine. Plant Physiol. 2008, 146, 236–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bilgin, D.D.; Zavala, J.A.; Zhu, J.; Clough, S.J.; Ort, D.R.; DeLUCIA, E.H. Biotic stress globally downregulates photosynthesis genes. Plant Cell Environ. 2010, 33, 1597–1613. [Google Scholar] [CrossRef] [Green Version]
- Lyu, X.; Shen, C.; Xie, J.; Fu, Y.; Jiang, D.; Hu, Z.; Tang, L.; Tang, L.; Ding, F.; Li, K. A “footprint” of plant carbon fixation cycle functions during the development of a heterotrophic fungus. Sci. Rep. 2015, 5, 1–13. [Google Scholar] [CrossRef]
- Tanabe, S.; Ashikari, M.; Fujioka, S.; Takatsuto, S.; Yoshida, S.; Yano, M.; Yoshimura, A.; Kitano, H.; Matsuoka, M.; Fujisawa, Y. A novel cytochrome P450 is implicated in brassinosteroid biosynthesis via the characterization of a rice dwarf mutant, dwarf11, with reduced seed length. Plant Cell 2005, 17, 776–790. [Google Scholar] [CrossRef] [Green Version]
- Kang, J.-G.; Yun, J.; Kim, D.-H.; Chung, K.-S.; Fujioka, S.; Kim, J.-I.; Dae, H.-W.; Yoshida, S.; Takatsuto, S.; Song, P.-S. Light and brassinosteroid signals are integrated via a dark-induced small G protein in etiolated seedling growth. Cell 2001, 105, 625–636. [Google Scholar] [CrossRef] [Green Version]
- Ali, S.S.; Gunupuru, L.R.; Kumar, G.S.; Khan, M.; Scofield, S.; Nicholson, P.; Doohan, F.M. Plant disease resistance is augmented in uzu barley lines modified in the brassinosteroid receptor BRI1. BMC Plant Biol. 2014, 14, 227. [Google Scholar] [CrossRef] [Green Version]
- Singh, A. Glucosinolates and Plant Defense. In Glucosinolates; Mérillon, J.M., Ramawat, K.G., Eds.; Springer: Berlin/Heidelberg, Germany, 2017; pp. 237–246. [Google Scholar]
- Ghanashyam, C.; Jain, M. Role of auxin-responsive genes in biotic stress responses. Plant Signal. Behav. 2009, 4, 846–848. [Google Scholar] [CrossRef]
- Spartz, A.K.; Ren, H.; Park, M.Y.; Grandt, K.N.; Lee, S.H.; Murphy, A.S.; Sussman, M.R.; Overvoorde, P.J.; Gray, W.M. SAUR inhibition of PP2C-D phosphatases activates plasma membrane H+-ATPases to promote cell expansion in Arabidopsis. Plant Cell 2014, 26, 2129–2142. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.; Pizarro, L.; Leibman-Markus, M.; Marash, I.; Bar, M. Cytokinin response induces immunity and fungal pathogen resistance, and modulates trafficking of the PRR LeEIX2 in tomato. Mol. Plant Pathol. 2020, 21, 1287–1306. [Google Scholar] [CrossRef] [PubMed]
- Pham, J.; Liu, J.; Bennett, M.H.; Mansfield, J.W.; Desikan, R. Arabidopsis histidine kinase 5 regulates salt sensitivity and resistance against bacterial and fungal infection. New Phytol. 2012, 194, 168–180. [Google Scholar] [CrossRef] [PubMed]
- Hutchison, C.E.; Li, J.; Argueso, C.; Gonzalez, M.; Lee, E.; Lewis, M.W.; Maxwell, B.B.; Perdue, T.D.; Schaller, G.E.; Alonso, J.M. The Arabidopsis histidine phosphotransfer proteins are redundant positive regulators of cytokinin signaling. Plant Cell 2006, 18, 3073–3087. [Google Scholar] [CrossRef] [Green Version]
- Argueso, C.T.; Ferreira, F.J.; Epple, P.; To, J.P.; Hutchison, C.E.; Schaller, G.E.; Dangl, J.L.; Kieber, J.J. Two-component elements mediate interactions between cytokinin and salicylic acid in plant immunity. PLoS Genet. 2012, 8, e1002448. [Google Scholar] [CrossRef] [Green Version]
- Han, X.; Kahmann, R. Manipulation of phytohormone pathways by effectors of filamentous plant pathogens. Front. Plant Sci. 2019, 10, 822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brocard, I.M.; Lynch, T.J.; Finkelstein, R.R. Regulation and role of the Arabidopsis abscisic acid-insensitive 5 gene in abscisic acid, sugar, and stress response. Plant Physiol. 2002, 129, 1533–1543. [Google Scholar] [CrossRef] [Green Version]
- Guerreiro, A.; Figueiredo, J.; Sousa Silva, M.; Figueiredo, A. Linking jasmonic acid to grapevine resistance against the biotrophic oomycete Plasmopara viticola. Front. Plant Sci. 2016, 7, 565. [Google Scholar] [CrossRef] [Green Version]
- Shah, J. The salicylic acid loop in plant defense. Curr. Opin. Plant Biol. 2003, 6, 365–371. [Google Scholar] [CrossRef]
- Meng, X.; Zhang, S. MAPK cascades in plant disease resistance signaling. Annu. Rev. Phytopathol. 2013, 51, 245–266. [Google Scholar] [CrossRef] [PubMed]
- Pedley, K.F.; Martin, G.B. Role of mitogen-activated protein kinases in plant immunity. Curr. Opin. Plant Biol. 2005, 8, 541–547. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Yan, H.; Li, J.; Tang, D. BSK1, a receptor-like cytoplasmic kinase, involved in both BR signaling and innate immunity in Arabidopsis. Plant Signal. Behav. 2013, 8, e24996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; He, C. A review of redox signaling and the control of MAP kinase pathway in plants. Redox Biol. 2017, 11, 192–204. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo, O.; Piqueras, R.; Sánchez-Serrano, J.J.; Solano, R. ETHYLENE RESPONSE FACTOR1 integrates signals from ethylene and jasmonate pathways in plant defense. Plant Cell 2003, 15, 165–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jose, J.; Ghantasala, S.; Roy Choudhury, S. Arabidopsis Transmembrane Receptor-Like Kinases (RLKs): A Bridge between Extracellular Signal and Intracellular Regulatory Machinery. Int. J. Mol. Sci. 2020, 21, 4000. [Google Scholar] [CrossRef] [PubMed]
- Magyarosy, A.C.; Schürmann, P.; Buchanan, B.B. Effect of powdery mildew infection on photosynthesis by leaves and chloroplasts of sugar beets. Plant Physiol. 1976, 57, 486–489. [Google Scholar] [CrossRef] [Green Version]
- Wientjes, E.; van Stokkum, I.H.; van Amerongen, H.; Croce, R. The role of the individual Lhcas in photosystem I excitation energy trapping. Biophys. J. 2011, 101, 745–754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, X.-Q.; Luo, X.-M.; Wang, Y.-L.; Xu, Q.-J.; Bai, L.-J.; Yuan, H.-J.; Tashi, N. Transcriptome sequencing in a Tibetan barley landrace with high resistance to powdery mildew. Sci. World J. 2014, 2014, 594579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Chen, Y.; Niu, Y.; Zhang, X.; Zhao, J.; Sun, L.; Wang, H.; Xiao, J.; Wang, X. Characterization and fine mapping of a leaf yellowing mutant in common wheat. Plant Growth Regul. 2020, 92, 233–247. [Google Scholar] [CrossRef]
- Hu, Y.; Zhong, S.; Zhang, M.; Liang, Y.; Gong, G.; Chang, X.; Tan, F.; Yang, H.; Qiu, X.; Luo, L. Potential Role of Photosynthesis in the Regulation of Reactive Oxygen Species and Defence Responses to Blumeria graminis f. sp. tritici in Wheat. Int. J. Mol. Sci. 2020, 21, 5767. [Google Scholar] [CrossRef]
- Wang, L.-F.; Wang, M.; Zhang, Y. Effects of powdery mildew infection on chloroplast and mitochondrial functions in rubber tree. Trop. Plant Pathol. 2014, 39, 242–250. [Google Scholar] [CrossRef] [Green Version]
- Gordon, T.R.; Duniway, J.M. Effects of powdery mildew infection on the efficiency of CO2 fixation and light utilization by sugar beet leaves. Plant Physiol. 1982, 69, 139–142. [Google Scholar] [CrossRef] [Green Version]
- Uematsu, K.; Suzuki, N.; Iwamae, T.; Inui, M.; Yukawa, H. Increased fructose 1, 6-bisphosphate aldolase in plastids enhances growth and photosynthesis of tobacco plants. J. Exp. Bot. 2012, 63, 3001–3009. [Google Scholar] [CrossRef]
- Nunes-Nesi, A.; Carrari, F.; Lytovchenko, A.; Smith, A.M.; Loureiro, M.E.; Ratcliffe, R.G.; Sweetlove, L.J.; Fernie, A.R. Enhanced photosynthetic performance and growth as a consequence of decreasing mitochondrial malate dehydrogenase activity in transgenic tomato plants. Plant Physiol. 2005, 137, 611–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suh, S.W.; Cascio, D.; Chapman, M.S.; Eisenberg, D. A crystal form of ribulose-1, 5-bisphosphate carboxylase/oxygenase from Nicotiana tabacum in the activated state. J. Mol. Biol. 1987, 197, 363–365. [Google Scholar] [CrossRef]
- Staskawicz, B.J. Genetics of plant-pathogen interactions specifying plant disease resistance. Plant Physiol. 2001, 125, 73–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Zhou, J.-M. Plant immunity triggered by microbial molecular signatures. Mol. Plant 2010, 3, 783–793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, A.; Wang, X.; Leseberg, C.H.; Jia, J.; Mao, L. Biotic and abiotic stress responses through calcium-dependent protein kinase (CDPK) signaling in wheat (Triticum aestivum L.). Plant Signal. Behav. 2008, 3, 654–656. [Google Scholar] [CrossRef] [Green Version]
- Seo, Y.-S.; Lee, S.-K.; Song, M.-Y.; Suh, J.-P.; Hahn, T.-R.; Ronald, P.; Jeon, J.-S. The HSP90-SGT1-RAR1 molecular chaperone complex: A core modulator in plant immunity. J. Plant Biol. 2008, 51, 1–10. [Google Scholar] [CrossRef]
- Wang, Z.-Y. Brassinosteroids modulate plant immunity at multiple levels. Proc. Natl. Acad. Sci. USA 2012, 109, 7–8. [Google Scholar] [CrossRef] [Green Version]
- Radojčić Redovniković, I.; Glivetić, T.; Delonga, K.; Vorkapić-Furač, J. Glucosinolates and their potential role in plant. Period. Biol. 2008, 110, 297–309. [Google Scholar]
- Gullner, G.; Kömives, T. The role of glutathione and glutathione-related enzymes in plant-pathogen interactions. In Significance of Glutathione to Plant Adaptation to the Environment; Springer: Berlin/Heidelberg, Germany, 2001; pp. 207–239. [Google Scholar]
- Nakashita, H.; Yasuda, M.; Nitta, T.; Asami, T.; Fujioka, S.; Arai, Y.; Sekimata, K.; Takatsuto, S.; Yamaguchi, I.; Yoshida, S. Brassinosteroid functions in a broad range of disease resistance in tobacco and rice. Plant J. 2003, 33, 887–898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubreuil-Maurizi, C.; Poinssot, B. Role of glutathione in plant signaling under biotic stress. Plant Signal. Behav. 2012, 7, 210–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Zahaby, H.; Gullner, G.; Kiraly, Z. Effects of powdery mildew infection of barley on the ascorbate-glutathione cycle and other antioxidants in different host-pathogen interactions. Phytopathology 1995, 85, 1225–1230. [Google Scholar] [CrossRef]
- Gullner, G.; Komives, T.; Király, L.; Schröder, P. Glutathione S-transferase enzymes in plant-pathogen interactions. Front. Plant Sci. 2018, 9, 1836. [Google Scholar] [CrossRef] [Green Version]
- Harrach, B.D.; Fodor, J.; Pogány, M.; Preuss, J.; Barna, B. Antioxidant, ethylene and membrane leakage responses to powdery mildew infection of near-isogenic barley lines with various types of resistance. Eur. J. Plant Pathol. 2008, 121, 21–33. [Google Scholar] [CrossRef]
- Jambagi, S.; Dunwell, J.M. Global transcriptome analysis and identification of differentially expressed genes after infection of Fragaria vesca with powdery mildew (Podosphaera aphanis). Transcr. Open Access 2015, 3, 1000106. [Google Scholar] [CrossRef] [Green Version]
- Weng, K.; Li, Z.-Q.; Liu, R.-Q.; Wang, L.; Wang, Y.-J.; Xu, Y. Transcriptome of Erysiphe necator-infected Vitis pseudoreticulata leaves provides insight into grapevine resistance to powdery mildew. Hortic. Res. 2014, 1, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Molitor, A.; Zajic, D.; Voll, L.M.; Pons-Kühnemann, J.; Samans, B.; Kogel, K.-H.; Waller, F. Barley leaf transcriptome and metabolite analysis reveals new aspects of compatibility and Piriformospora indica–mediated systemic induced resistance to powdery mildew. Mol. Plant Microbe Interact. 2011, 24, 1427–1439. [Google Scholar] [CrossRef] [Green Version]
- Seeholzer, S.; Tsuchimatsu, T.; Jordan, T.; Bieri, S.; Pajonk, S.; Yang, W.; Jahoor, A.; Shimizu, K.K.; Keller, B.; Schulze-Lefert, P. Diversity at the Mla powdery mildew resistance locus from cultivated barley reveals sites of positive selection. Mol. Plant Microbe Interact. 2010, 23, 497–509. [Google Scholar] [CrossRef] [Green Version]
- Consonni, C.; Humphry, M.E.; Hartmann, H.A.; Livaja, M.; Durner, J.; Westphal, L.; Vogel, J.; Lipka, V.; Kemmerling, B.; Schulze-Lefert, P. Conserved requirement for a plant host cell protein in powdery mildew pathogenesis. Nat. Genet. 2006, 38, 716–720. [Google Scholar] [CrossRef] [PubMed]
- Sanderfoot, A.A.; Assaad, F.F.; Raikhel, N.V. The Arabidopsis genome. An abundance of soluble N-ethylmaleimide-sensitive factor adaptor protein receptors. Plant Physiol. 2000, 124, 1558–1569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bracuto, V.; Appiano, M.; Zheng, Z.; Wolters, A.-M.A.; Yan, Z.; Ricciardi, L.; Visser, R.G.; Pavan, S.; Bai, Y. Functional characterization of a syntaxin involved in tomato (Solanum lycopersicum) resistance against powdery mildew. Front. Plant Sci. 2017, 8, 1573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavan, S.; Jacobsen, E.; Visser, R.G.; Bai, Y. Loss of susceptibility as a novel breeding strategy for durable and broad-spectrum resistance. Mol. Breed. 2010, 25, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reiner, T.; Hoefle, C.; Hückelhoven, R. A barley SKP1-like protein controls abundance of the susceptibility factor RACB and influences the interaction of barley with the barley powdery mildew fungus. Mol. Plant Pathol. 2016, 17, 184–195. [Google Scholar] [CrossRef]
- Li, Y.; Qiu, L.; Liu, X.; Zhang, Q.; Zhuansun, X.; Fahima, T.; Krugman, T.; Sun, Q.; Xie, C. Glycerol-induced powdery mildew resistance in wheat by regulating plant fatty acid metabolism, plant hormones cross-talk, and pathogenesis-related genes. Int. J. Mol. Sci. 2020, 21, 673. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.; Liang, K.; Zhou, Z.; Wang, X.; Huang, G. UPLC-ESI-MS/MS-Based Widely Targeted Metabolomics Analysis of Wood Metabolites in Teak (Tectona grandis). Molecules 2020, 25, 2189. [Google Scholar] [CrossRef]
- Chen, L.; Wu, Q.; He, W.; He, T.; Wu, Q.; Miao, Y. Combined De Novo Transcriptome and Metabolome Analysis of Common Bean Response to Fusarium oxysporum f. sp. phaseoli Infection. Int. J. Mol. Sci. 2019, 20, 6278. [Google Scholar] [CrossRef] [Green Version]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat Biotech 2011, 29, 644. [Google Scholar] [CrossRef] [Green Version]
- Zeng, X.; Long, H.; Wang, Z.; Zhao, S.; Tang, Y.; Huang, Z.; Wang, Y.; Xu, Q.; Mao, L.; Deng, G.; et al. The draft genome of Tibian hulless barley reveals adaptive patterns to the high stressful Tibean Plateau. PNAS 2015, 112, 1095–1100. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varet, H.; Brillet-Guéguen, L.; Coppée, J.-Y.; Dillies, M.-A. SARTools: A DESeq2-and edgeR-based R pipeline for comprehensive differential analysis of RNA-Seq data. PLoS ONE 2016, 11, e0157022. [Google Scholar]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anders, S.; McCarthy, D.J.; Chen, Y.; Okoniewski, M.; Smyth, G.K.; Huber, W.; Robinson, M.D. Count-based differential expression analysis of RNA sequencing data using R and Bioconductor. Nat Protoc. 2013, 8, 1765. [Google Scholar] [CrossRef]
- Heberle, H.; Meirelles, G.V.; da Silva, F.R.; Telles, G.P.; Minghim, R. InteractiVenn: A web-based tool for the analysis of sets through Venn diagrams. BMC Bioinform. 2015, 16, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S.; Kawashima, S.; Okuno, Y.; Hattori, M. The KEGG resource for deciphering the genome. Nucleic Acids Res. 2004, 32, D277–D280. [Google Scholar] [CrossRef] [Green Version]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene Ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 25. [Google Scholar] [CrossRef] [Green Version]
- Tatusov, R.L.; Galperin, M.Y.; Natale, D.A.; Koonin, E.V. The COG Database: A tool for fenome-scale analysis of protein functions and evolution. Nucleic Acids Res. 2000, 28, 33–36. [Google Scholar] [CrossRef] [Green Version]
- Apweiler, R.; Bairoch, A.; Wu, C.H.; Barker, W.C.; Boeckmann, B.; Ferro, S.; Gasteiger, E.; Huang, H.; Lopez, R.; Magrane, M. UniProt: The universal protein knowledgebase. Nucleic Acids Res. 2004, 32, D115–D119. [Google Scholar] [CrossRef]
- Koonin, E.V.; Fedorova, N.D.; Jackson, J.D.; Jacobs, A.R.; Krylov, D.M.; Makarova, K.S.; Mazumder, R.; Mekhedov, S.L.; Nikolskaya, A.N.; Rao, B.S. A comprehensive evolutionary classification of proteins encoded in complete eukaryotic genomes. Genome. Biol. 2004, 5, R7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genom. Biol. 2009, 10, R25. [Google Scholar] [CrossRef] [Green Version]
- Xie, C.; Mao, X.; Huang, J.; Ding, Y.; Wu, J.; Dong, S.; Kong, L.; Gao, G.; Li, C.-Y.; Wei, L. KOBAS 2.0: A web server for annotation and identification of enriched pathways and diseases. Nucleic Acids Res. 2011, 39, W316–W322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Chen, H.; He, Y.; Xia, R. TBtools, a toolkit for biologists integrating various biological data handling tools with a user-friendly interface. BioRxiv 2018, 289660. [Google Scholar] [CrossRef]
- Walling, J.G.; Zalapa, L.A.; Vinje, M.A. Evaluation and selection of internal reference genes from two-and six-row US malting barley varieties throughout micromalting for use in RT-qPCR. PLoS ONE 2018, 13, e0196966. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primers |
|---|---|
| GAPDH | F-CTAAGTTTTTGGGTGT R-CTATGAATTGGTCTTCC |
| HOVUSG4120900 | F-GGCACCTCCAAGCACCAGAT R-ACACCCGAACCAAAGTAGCG |
| HOVUSG6135200 | F-CTTTTGGGAGAACGATGGA R-TGTTTGGTAGGTGCTCTTTATG |
| HOVUSG0289600 | F-TAATCTATTGGATGACACTGGGA R-CTGTGAGAGGCTTTGCTTGA |
| HOVUSG2614500 | F-CGCATTTGTCGCAAAACA R-GATGAGCGTGCCAACCC |
| HOVUSG2888400 | F-GGGTCCTTGTCTGCCTGG R-CGCTCTGTTTCTTGCTTCC |
| HOVUSG3967100 | F-GGATGAAGGGGAAGACGG R-ACAGAGGGTCGGCGGAGA |
| HOVUSG2306000 | F-TCAGTGCGGTAGAGCGAGC R-GGTGACGCCGAGGATGGG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sang, Z.; Zhang, M.; Mu, W.; Yang, H.; Yang, C.; Xu, Q. Phytohormonal and Transcriptomic Response of Hulless Barley Leaf in Response to Powdery Mildew Infection. Agronomy 2021, 11, 1248. https://doi.org/10.3390/agronomy11061248
Sang Z, Zhang M, Mu W, Yang H, Yang C, Xu Q. Phytohormonal and Transcriptomic Response of Hulless Barley Leaf in Response to Powdery Mildew Infection. Agronomy. 2021; 11(6):1248. https://doi.org/10.3390/agronomy11061248
Chicago/Turabian StyleSang, Zha, Minjuan Zhang, Wang Mu, Haizhen Yang, Chunbao Yang, and Qijun Xu. 2021. "Phytohormonal and Transcriptomic Response of Hulless Barley Leaf in Response to Powdery Mildew Infection" Agronomy 11, no. 6: 1248. https://doi.org/10.3390/agronomy11061248