Monitoring of Natural Occurrence and Severity of Leaf and Glume Blotch Diseases of Winter Wheat and Winter Triticale Incited by Necrotrophic Fungi Parastagonospora spp. and Zymoseptoria tritici

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
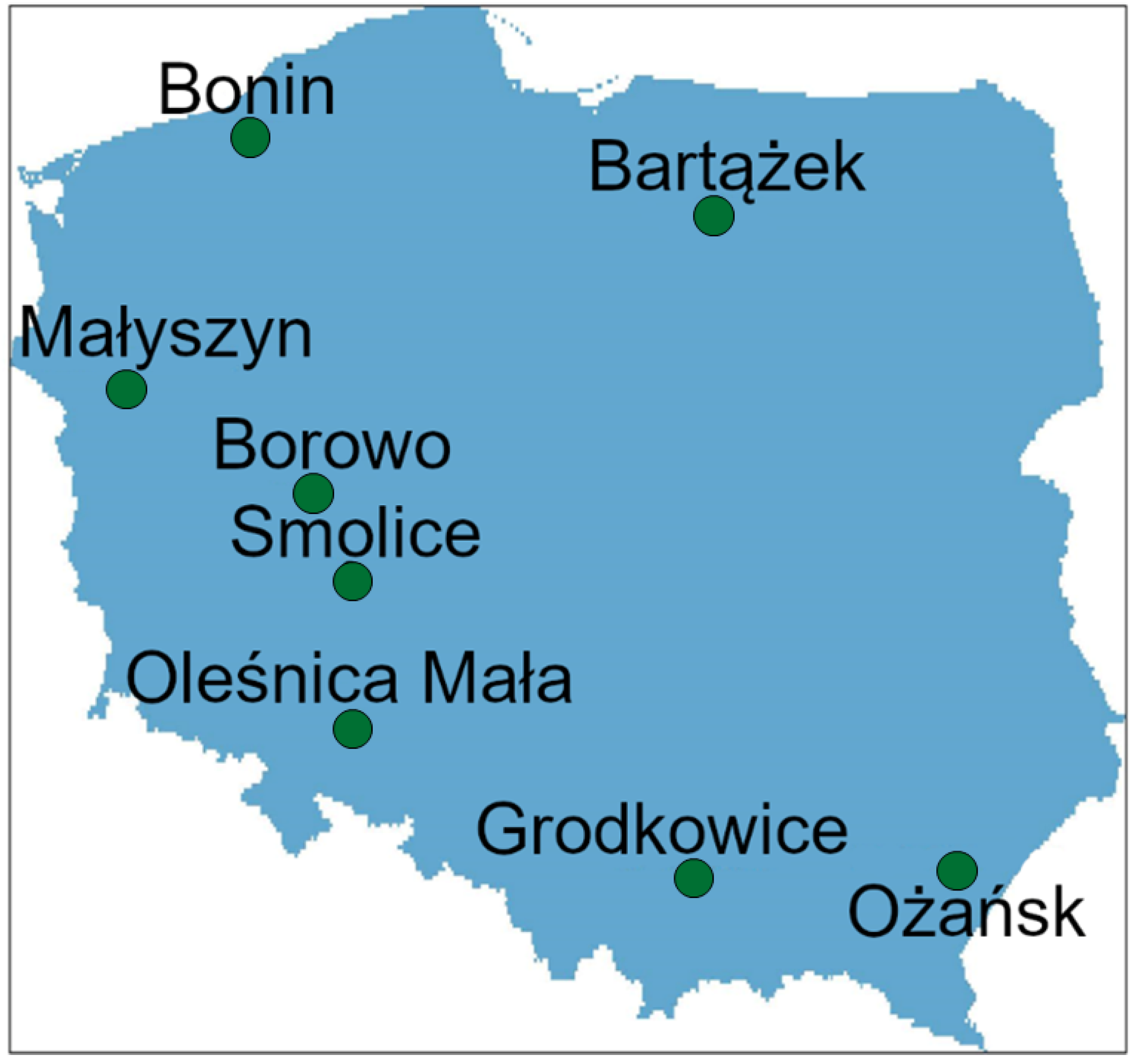
2.1. Plant Materials and Field Experiments
2.2. Mycological Analyses
2.3. Statistical Analysis
3. Results and Discussion
3.1. Occurrence of Pathogens on Wheat and Triticale
3.2. Correlation between Meteorological Factors and Occurrence of Pathogens
3.2.1. Correlation between Meteorological Factors and Occurrence of P. nodorum
3.2.2. Correlation between Meteorological Factors and Occurrence of P. avenae f. sp. triticea
3.2.3. Correlation between Meteorological Factors and Occurrence of Z. tritici
3.2.4. Summary of Correlation between Meteorological Factors and Occurrence of Pathogens
3.3. The Pathogen Occurrence Models
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Johnson, T. A form of leptosphaeria avenaria on wheat in canada. Can. J. Res. 1947, 25, 259–270. [Google Scholar] [CrossRef]
- Arseniuk, E.; Fried, P.M.; Winzeler, H.; Czembor, H.J. Comparison of resistance of triticale, wheat and spelt to Septoria nodorum blotch at the seedling and adult plant stages. Euphytica 1991, 55, 43–48. [Google Scholar] [CrossRef]
- Leath, S.; Scharen, A.L.; Lund, R.E.; Dietz-Holmes, M.E. Factors associated with global occurrences of Septoria nodorum blotch and Septoria tritici blotch of wheat. Plant Dis. 1993, 77, 1266–1270. [Google Scholar] [CrossRef]
- El Harrak, A.; Mergoum, M.; Saadaoui, E. Major Foliar Diseases of Triticale in Morocco; CIMMYT: Mexico City, Mexico, 1997; pp. 150–156. [Google Scholar]
- Małkus, A.; Reszka, E.; Chang, C.J.; Arseniuk, E.; Linda Chang, P.L.; Ueng, P.P. Sequence diversity of β-tubulin (tubA) gene in Phaeosphaeria nodorum and P. avenaria. FEMS Microbiol. Lett. 2005, 249, 49–56. [Google Scholar] [CrossRef] [Green Version]
- Reszka, E.; Chung, K.R.; Tekauz, A.; Malkus, A.; Arseniuk, E.; Krupinsky, J.M.; Tsang, H.; Ueng, P.P. Presence of β-glucosidase (bgl1) gene in Phaeosphaeria nodorum and Phaeosphaeria avenaria f.sp. triticea. Can. J. Bot. 2005, 83, 1001–1014. [Google Scholar] [CrossRef]
- Tian, S.; Wolf, G.A.; Weinert, J. Accurate Assessment of Wheat and Triticale Cultivar Resistance to Septoria tritici and Stagonospora nodorum Infection by Biotin/Avidin ELISA. Plant Dis. 2005, 89, 1229–1234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quaedvlieg, W.; Verkley, G.; Shin, H.-D.; Barreto, R.; Alfenas, A.; Swart, W.; Groenewald, J.; Crous, P. Sizing up Septoria. Stud. Mycol. 2013, 75, 307–390. [Google Scholar] [CrossRef] [Green Version]
- Gilon, M.; Arseniuk, E. Natural field infections of wheat and triticale by fungi from the complex of fungi Stagonospora nodorum/Septoria tritici under climatic conditions of Poland. Commun. Agric. Appl. Biol. Sci. 2014, 79, 216–227. [Google Scholar] [PubMed]
- Downie, R.C.; Lin, M.; Corsi, B.; Ficke, A.; Lillemo, M.; Oliver, R.P.; Phan, H.; Tan, K.-C.; Cockram, J. Septoria nodorum blotch of wheat: Disease management and resistance breeding in the face of shifting disease dynamics and a changing environment. Phytopathology 2020. [Google Scholar] [CrossRef]
- Bhathal, J.; Loughman, R.; Speijers, J. Yield Reduction in Wheat in Relation to Leaf Disease From Yellow (tan) Spot and Septoria Nodorum Blotch. Eur. J. Plant Pathol. 2003, 109, 435–443. [Google Scholar] [CrossRef]
- Bearchell, S.J.; Fraaije, B.A.; Shaw, M.W.; Fitt, B.D.L. Wheat archive links long-term fungal pathogen population dynamics to air pollution. Proc. Natl. Acad. Sci. USA 2005, 102, 5438–5442. [Google Scholar] [CrossRef] [Green Version]
- Lin, M.; Ficke, A.; Cockram, J.; Lillemo, M. Genetic Structure of the Norwegian Parastagonospora nodorum Population. Front. Microbiol. 2020, 11, 1280. [Google Scholar] [CrossRef]
- Hekuran, V.; Belul, G.; Foto, K.; Halit, S.; Thanas, R. The Relationship between diseases index of septoria leaf blotch, leaf rust and yield losses in bread wheat cultivar in Albania. J. Environ. Sci. Eng. B 2012, 1, 957–965. [Google Scholar]
- Witkowska, K.; Śmiałowski, T.; Witkowski, E. Dependence of the Yield of Winter Wheat Sows on the Degree of Infection by Stagonospora Nodorum and Puccinia Triticina under Different Field Conditions. J. Bull. Plant Breed. Acclim. Inst. 2011, 262, 47–57. [Google Scholar]
- Torriani, S.F.; Melichar, J.P.; Mills, C.; Pain, N.; Sierotzki, H.; Courbot, M. Zymoseptoria tritici: A major threat to wheat production, integrated approaches to control. Fungal Genet. Biol. 2015, 79, 8–12. [Google Scholar] [CrossRef] [PubMed]
- Głazek, M.; Krzyzińska, B.; Maczyńska, A. Occurrence of Stagonospora nodorum glume blotch of wheat in the region of middle-southern Poland. Acta Agrobot. 2012, 58, 23–28. [Google Scholar] [CrossRef] [Green Version]
- Arseniuk, E.; Gilon, M. Monitoring of pathogenicity changes in populations of necrotrophic pathogens of cereals (Stagonospora spp.; Septoria tritici). Monographs and Scientific Papers. In Improving the Plant for Sustainable Agro-Ecosystems, High-Quality Food and Non-Food Plant Production; Monographs and Scientific Papers IHAR-PIB: Błonie, Poland, 2014; Volume 48, pp. 272–278. [Google Scholar]
- Cowger, C.; Silva-Rojas, H.V. Frequency of Phaeosphaeria nodorum, the Sexual Stage of Stagonospora nodorum, on Winter Wheat in North Carolina. Phytopathology 2006, 96, 860–866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solomon, P.S.; Lowe, R.G.T.; Tan, K.-C.; Waters, O.D.C.; Oliver, R.P. Stagonospora nodorum: Cause of stagonospora nodorum blotch of wheat. Mol. Plant Pathol. 2006, 7, 147–156. [Google Scholar] [CrossRef] [PubMed]
- McDonald, M.C.; Razavi, M.; Friesen, T.L.; Brunner, P.C.; McDonald, B.A. Phylogenetic and population genetic analyses of Phaeosphaeria nodorum and its close relatives indicate cryptic species and an origin in the Fertile Crescent. Fungal Genet. Biol. 2012, 49, 882–895. [Google Scholar] [CrossRef]
- Arseniuk, E.; Góral, T.; Scharen, A.L. Seasonal Patterns of Spore Dispersal of Phaeosphaeria spp. and Stagonospora spp. Plant Dis. 1998, 82, 187–194. [Google Scholar] [CrossRef] [Green Version]
- Mirzwa-Mróz, E.; Tvaruzek, L.; Zamorski Cz Nowicki, B. Research on the development of Mycosphaerella graminicola (Fuckel) Schroeter telemorph on wheat leaves from Poland and Czech Republic. Acta Agrobot. 2005, 58, 59–65. [Google Scholar] [CrossRef] [Green Version]
- Pietravalle, S.; Shaw, M.W.; Parker, S.R.; Bosch, F.V.D. Modeling of Relationships Between Weather and Septoria tritici Epidemics on Winter Wheat: A Critical Approach. Phytopathology 2003, 93, 1329–1339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahtour, A.; El Jarroudi, M.; Delobbe, L.; Hoffmann, L.; Maraite, H.; Tychon, B. Site-Specific Septoria Leaf Blotch Risk Assessment in Winter Wheat Using Weather-Radar Rainfall Estimates. Plant Dis. 2011, 95, 384–393. [Google Scholar] [CrossRef] [PubMed]
- West, J.S.; Townsend, J.A.; Stevens, M.; Fitt, B.D. Comparative biology of different plant pathogens to estimate effects of climate change on crop diseases in European. J. Plant Pathol. 2012, 133, 315–331. [Google Scholar] [CrossRef] [Green Version]
- Fones, H.; Gurr, S. The impact of Septoria tritici Blotch disease on wheat: An EU perspective. Fungal Genet. Biol. 2015, 79, 3–7. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, M.R.; Stevenson, C.F.; Hodge, K.; Dokken-Bouchard, F.; Pearse, P.G.; Waelchli, F.; Brown, A.; Peluola, C. Assessing Effects of Climatic Change, Region and Agronomic Practices on Leaf Spotting of Bread and Durum Wheat in the Western Canadian Prairies, from 2001 to 2012. Agron. J. 2016, 108, 1180–1195. [Google Scholar] [CrossRef] [Green Version]
- Shaw, M.W.; Bearchell, S.J.; Fitt BD, L.; Fraaije, B.A. Long-term relationships between environment and abundance in wheat of Phaeosphaeria nodorum and Mycosphaerella graminicola. New Phytol. Found. 2007, 77, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Crook, A.D.; Friesen, T.L.; Liu, Z.H.; Ojiambo, P.S.; Cowger, C. Novel Necrotrophic Effectors from Stagonospora nodorum and Corresponding Host Sensitivities in Winter Wheat Germplasm in the Southeastern United States. Phytopathology 2012, 102, 498–505. [Google Scholar] [CrossRef] [Green Version]
- Heick, T.M.; Fejer, A.; Nistrup Jørgensen, J.L. Resistance of wheat pathogen Zymoseptoria tritici to DMI and QoI fungicides in the Nordic-Baltic region-a status. Eur. J. Plant Pathol. 2017, 149, 669–682. [Google Scholar] [CrossRef]
- Semaskiene, R.; Ronis, A. Leaf spotting diseases in winter and spring wheat and resistance to fungicides in Mycosphaerella graminicola. In Proceedings of the 8th International Symposium on Mycosphaerella and Stagonospora Diseases of Cereals, Mexico City, Mexico, 19 September 2011. [Google Scholar]
- Solomon, P.; Parker, K.; Loughman, R.; Oliver, R. Both Mating types of Phaeosphaeria (anamorph Stagonospora) nodorum are Present in Western Australia. Eur. J. Plant Pathol. 2004, 110, 763–766. [Google Scholar] [CrossRef]
- Shaner, G.; Buechley, C. Epidemiology of leaf blotch of soft red winter wheat caused by Septoria tritici and Stagonospora nodorum. Plant Dis. 1995, 19, 928–938. [Google Scholar] [CrossRef]
- Shaw, M.W.; Royle, D.J. Factors determining the severity of epidemics of Mycosphaerella graminicola (Septoria tritici) on winter wheat in the UK. Plant Pathol. 1993, 42, 882–899. [Google Scholar] [CrossRef]
- Bouma, E. Development of comparable agro-climatic zones for the international exchange of data on the efficacy and crop safety of plant protection products. Bulletin OEPP/EPPO 2005, 35, 233–238. [Google Scholar] [CrossRef]
- Hailemariam, B.N.; Kidane, Y.G.; Ayalew, A. Epidemiological factors of Septoria triticim blotch (Zymoseptoria tritici) in durum wheat (Triticum turgidum) in the highlands of Wollo, Ethiopia. Ecol. Process. 2020, 9, 61. [Google Scholar] [CrossRef]
- Henze, M.; Beyer, M.; Klink, H.; Verreet, J.-A. Characterizing Meteorological Scenarios Favorable for Septoria tritici Infections in Wheat and Estimation of Latent Periods. Plant Dis. 2007, 91, 1445–1449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDonald, B.A.; Mundt, C.C. How Knowledge of Pathogen Population Biology Informs Management of Septoria tritici Blotch. Phytopathology 2016, 106, 948–955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shearer, B.L.; Zadoks, J.C. Observations on the host range of an isolate of Septoria nodorum from wheat. Eur. J. Plant Pathol. 1972, 78, 153–159. [Google Scholar] [CrossRef]
- Eyal, Z.; Scharen, A.L.; Prescott, J.M.; van Ginkel, M. The Septoria Diseases of Wheat: Concepts and Methods of Disease Management; CIMMYT: Mexico City, Mexico, 1987. [Google Scholar]
- John, E.; Lopez-Ruiz, F.; Rybak, K.; Mousley, C.J.; Oliver, R.P.; Tan, K.-C. Dissecting the role of histidine kinase and HOG1 mitogen-activated protein kinase signalling in stress tolerance and pathogenicity of Parastagonospora nodorum on wheat. Microbiology 2016, 162, 1023–1036. [Google Scholar] [CrossRef]
- Saidi, A.; Eslahi, M.R.; Safaie, N. Efficiency of Septoria tritici sporulation on different culture media. Trakia J. Sci. 2012, 3, 15–18. [Google Scholar]
- Czembor, P.C.; Arseniuk, E. Study of genetic variability among monopycnidial and monopycnidiospore isolates derived from single pycnidia of Stagonospora ssp. and Septoria tritici with the use of RAPD-PCR, MP-PCR and rep-PCR Techniques. J. Phytopathol. 2008, 147, 539–546. [Google Scholar] [CrossRef]
- Pedregosa, F.; Varoquaux, G.; Gramfort, A.V.M.; Thirion, B.; Grisel, O.; Blondel, M.; Prettenhofer, P.; Weiss, R.; Dubourg, V.; Vanderplas, J.; et al. Scikit-learn: Machine Learning in Python. J. Mach. Learn. Res. 2011, 12, 2825–2830. [Google Scholar]
- The Pandas Development Team. Pandas-Dev/Pandas: Pandas; Zenodo: Geneva, Switzerland, 2020. [Google Scholar] [CrossRef]
- Harris, C.R.; Millman, K.J.; Van Der Walt, S.J.; Gommers, R.; Virtanen, P.; Cournapeau, D.; Wieser, E.; Taylor, J.; Berg, S.; Smith, N.J.; et al. Array programming with NumPy. Nature 2020, 585, 357–362. [Google Scholar] [CrossRef]
- Deisenroth, M.P.; Faisal, A.A.; Ong, C.S. Mathematics for Machine Learning; Cambridge University Press: Cambridge, UK, 2020. [Google Scholar]
- Sapkota, R.; Knorr, K.; Jørgensen, L.N.; O’Hanlon, K.A.; Nicolaisen, M. Host genotype is an important determinant of the cereal phyllosphere mycobiome. New Phytol. 2015, 207, 1134–1144. [Google Scholar] [CrossRef]
- Jones, D.G.; Cooke, B. The Epidemiology of Septoria tritici and S. nodorum: I. A tentative key for assessing Septoria tritici infection on wheat heads. Trans. Br. Mycol. Soc. 1969, 53, 39–46. [Google Scholar] [CrossRef]
- Arseniuk, E.; Walczewski, J.; Ochodzki, P. Protein toxins of Parastagonospora nodorum and their relation to pathogenicity and resistance of triticale and wheat to leaf and glume blotch. Commun. Agric. Appl. Biol. Sci. 2019, 286, 71–73. [Google Scholar]
- Mehra, L.K.; Cowger, C.; Gross, K.; Ojiambo, P.S. Predicting Pre-planting Risk of Stagonospora nodorum blotch in Winter Wheat Using Machine Learning Models. Front. Plant Sci. 2016, 7, 390. [Google Scholar] [CrossRef] [PubMed]
- Mehra, L.K.; Cowger, C.; Ojiambo, P.S. A Model for Predicting Onset of Stagonospora nodorum Blotch in Winter Wheat Based on Preplanting and Weather Factors. Phytopathology 2017, 107, 635–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghaderi, F.; Sharifnabi, B.; Javan-Nikkhah, M.; Brunner, P.C.; McDonald, B.A. SnToxA, SNTox1 and SnTox3 originated in Parastagonospora nodorum in the Fertile Crescent. Plant Pathol. 2020, 69, 1482–1491. [Google Scholar] [CrossRef]
- Chaloner, T.M.; Fones, H.; Varma, N.V.; Bebber, D.P.; Gurr, S.J. A new mechanistic model of weather-dependent Septoria tritici blotch disease risk. Philos. Trans. B 2019, 374, 20180266. [Google Scholar] [CrossRef] [Green Version]
- Eyal, Z. The Septoria tritici and Stagonospora Nodorum Blotch Diseases of Wheat. Eur. J. Plant Pathol. 1999, 105, 629–641. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Grouping Variable | Pathogen Pairs | Mean Number of Isolations | t | df | p | ||
|---|---|---|---|---|---|---|---|
| Winter wheat | Z. tritici | P. nodorum | 159.3 | 16.0 | −8.8 | 94 | 0.000 |
| Z. tritici | P. avenae | 159.3 | 1.8 | −10.2 | 94 | 0.000 | |
| P. nodorum | P. avenae | 16.0 | 1.8 | 2.9 | 94 | 0.005 | |
| Winter triticale | Z. tritici | P. nodorum | 7.8 | 25.2 | 3.2 | 94 | 0.002 |
| Z. tritici | P. avenae | 7.8 | 6.8 | −0.5 | 94 | 0.647 | |
| P. nodorum | P. avenae | 25.2 | 6.8 | 3.4 | 94 | 0.001 | |
| Month | Meteorological Factors | r | p |
|---|---|---|---|
| 1 | Number of days with snow cover | 0.202 | 0.049 |
| 2 | Number of days with rainfall | 0.247 | 0.015 |
| 2 | First day of maximum rainfall | 0.225 | 0.028 |
| 5 | Absolute maximum temperature | 0.232 | 0.023 |
| 5 | First day of maximum rainfall | 0.328 | 0.001 |
| 6 | Daily maximum sum of rainfall | 0.290 | 0.004 |
| 6 | Monthly sum of rainfall | 0.287 | 0.005 |
| 7 * | Average maximum temperature | −0.267 | 0.009 |
| 7 * | Monthly average temperature | −0.276 | 0.006 |
| 10 * | Absolute maximum temperature | −0.431 | 0.000 |
| 10 * | Average maximum temperature | −0.253 | 0.013 |
| 10 * | Monthly average temperature | −0.272 | 0.007 |
| 11 * | Number of days with snowfall | 0.257 | 0.012 |
| 11 * | Monthly sum of rainfall | 0.281 | 0.006 |
| 12 * | Number of days with snowfall | −0.247 | 0.015 |
| 12 * | Average maximum temperature | 0.276 | 0.006 |
| 12 * | Monthly average temperature | 0.297 | 0.003 |
| 12 * | Average minimum temperature | 0.285 | 0.005 |
| - | Geographical latitude | 0.292 | 0.004 |
| Month | Meteorological Factors | r | p |
|---|---|---|---|
| 2 | Absolute minimum temperature | 0.214 | 0.036 |
| 2 | Number of days with rainfall | 0.292 | 0.004 |
| 2 | Daily maximum sum of rainfall | 0.208 | 0.042 |
| 2 | Monthly sum of rainfall | 0.249 | 0.015 |
| 2 | Minimum near-surface temperature | 0.287 | 0.005 |
| 2 | First day of maximum rainfall | 0.293 | 0.004 |
| 2 | Monthly average temperature | 0.227 | 0.026 |
| 2 | Average minimum temperature | 0.241 | 0.018 |
| 3 | Number of days with rainfall | 0.264 | 0.009 |
| 5 | First day of maximum rainfall | 0.296 | 0.003 |
| 6 | Absolute maximum temperature | 0.205 | 0.045 |
| 7 * | Minimum near-surface temperature | −0.268 | 0.008 |
| 8 * | Absolute maximum temperature | 0.341 | 0.001 |
| 8 * | Number of days with rainfall | −0.325 | 0.001 |
| 8 * | Monthly sum of rainfall | −0.298 | 0.003 |
| 8 * | Average maximum temperature | 0.368 | 0.000 |
| 8 * | Monthly average temperature | 0.350 | 0.000 |
| 8 * | Average minimum temperature | 0.267 | 0.009 |
| 9 * | Absolute maximum temperature | 0.342 | 0.001 |
| 9 * | Daily maximum sum of rainfall | −0.235 | 0.021 |
| 10 * | Monthly sum of rainfall | −0.209 | 0.041 |
| 10 * | Monthly average temperature | −0.258 | 0.011 |
| 10 * | Average minimum temperature | −0.279 | 0.006 |
| 12 * | Absolute maximum temperature | 0.264 | 0.009 |
| 12 * | Number of days with snowfall | −0.294 | 0.004 |
| 12 * | Average maximum temperature | 0.399 | 0.000 |
| 12 * | Monthly average temperature | 0.394 | 0.000 |
| 12 * | Average minimum temperature | 0.362 | 0.000 |
| - | Plant species (host) | 0.307 | 0.002 |
| Month | Meteorological Factors | r | p |
|---|---|---|---|
| 1 | Monthly average temperature | −0.208 | 0.042 |
| 1 | Average minimum temperature | −0.218 | 0.033 |
| 2 | Absolute maximum temperature | 0.243 | 0.017 |
| 2 | Monthly sum of rainfall | 0.206 | 0.044 |
| 6 | Average maximum temperature | 0.237 | 0.020 |
| 6 | Monthly average temperature | 0.262 | 0.010 |
| 6 | Average minimum temperature | 0.252 | 0.013 |
| 8 * | Number of days with rainfall | −0.332 | 0.001 |
| 8 * | Average maximum temperature | 0.252 | 0.013 |
| 8 * | Monthly average temperature | 0.232 | 0.023 |
| 9 * | Absolute maximum temperature | 0.250 | 0.014 |
| 10 * | Average minimum temperature | −0.201 | 0.049 |
| - | Plant species (host) | −0.710 | 0.000 |
| Size of Training Dataset | Model for Pathogen | RMSE | r | ||
|---|---|---|---|---|---|
| Train Dataset | Test Dataset | Train Dataset | Test Dataset | ||
| 80% | P. nodorum | 16.86 | 30.43 | 0.88 | 0.58 |
| P. avenae | 2.94 | 7.20 | 0.95 | 0.50 | |
| Z. tritici | 44.11 | 60.19 | 0.91 | 0.84 | |
| 90% | P. nodorum | 17.23 | 27.62 | 0.87 | 0.62 |
| P. avenae | 2.82 | 6.86 | 0.95 | 0.53 | |
| Z. tritici | 47.71 | 53.80 | 0.89 | 0.88 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bartosiak, S.F.; Arseniuk, E.; Szechyńska-Hebda, M.; Bartosiak, E. Monitoring of Natural Occurrence and Severity of Leaf and Glume Blotch Diseases of Winter Wheat and Winter Triticale Incited by Necrotrophic Fungi Parastagonospora spp. and Zymoseptoria tritici. Agronomy 2021, 11, 967. https://doi.org/10.3390/agronomy11050967
Bartosiak SF, Arseniuk E, Szechyńska-Hebda M, Bartosiak E. Monitoring of Natural Occurrence and Severity of Leaf and Glume Blotch Diseases of Winter Wheat and Winter Triticale Incited by Necrotrophic Fungi Parastagonospora spp. and Zymoseptoria tritici. Agronomy. 2021; 11(5):967. https://doi.org/10.3390/agronomy11050967
Chicago/Turabian StyleBartosiak, Sławomir Franciszek, Edward Arseniuk, Magdalena Szechyńska-Hebda, and Ewa Bartosiak. 2021. "Monitoring of Natural Occurrence and Severity of Leaf and Glume Blotch Diseases of Winter Wheat and Winter Triticale Incited by Necrotrophic Fungi Parastagonospora spp. and Zymoseptoria tritici" Agronomy 11, no. 5: 967. https://doi.org/10.3390/agronomy11050967