1. Introduction
Tomato (
Solanum lycopersicum L.), a staple vegetable crop with global distribution, exhibits obvious heterosis, especially with respect to early maturity and yield [
1]. Heterosis, a common natural phenomenon, is the biological basis of crossbreeding; it depends on genetic differences between the parents [
2], is the most profound in F
1, and gradually diminishes starting in F
2. It is an extremely important means of genetic improvement and has been widely used in field crops (e.g., rice and maize), as well as vegetable crops (e.g., species of Cruciferae and Solanaceae) [
3]. For example, in Europe, ≈100% of sugar beet, >90% of rapeseed, and >70% of rye varieties are hybrids, as are >70% of rice varieties in China and >80% of varieties cotton in India [
4]. Utilizing heterosis in agricultural production can significantly improve yield [
5,
6,
7], quality [
8,
9,
10], and resistance [
11,
12,
13]. The utilization of crop heterosis is considered a landmark innovation of modern agriculture and has yielded great economic gains [
4].
Analyzing heterosis and combining ability are two important considerations in the utilization of heterosis; it is the first step in breeding inbred lines to develop commercial hybrids. Progeny selection is one of the most important stages in plant breeding, but producing excellent progeny depends on the chosen parents. Combining ability is useful for successfully predicting the genetic capabilities of parental lines and crosses [
14,
15]. Therefore, the level of combining ability is an important criterion for selecting the parents of hybrids, and it directly affects hybrid quality [
16]. For some traits, parents with high GCA effects produce hybrids with low SCA effects. On the other hand, parents with low GCA effects can produce hybrids with high SCA effects. Therefore, when selecting elite parents for crosses, understanding and accounting for the relationship between the GCA of the parents, the SCA of the cross, and the dominant type of combining ability are key to improving breeding efficiency based on combining ability [
3]. Some studies indicate that there is no fixed relationship between GCA and SCA [
17,
18,
19,
20]. Xiang et al. revealed a correlation between parental traits and GCA [
21], but parental performance is not correlated with heterosis [
22]. Therefore, parental performance per se and GCA are not necessarily reliable predictors of heterosis.
Many studies have shown that parental GCA may be a good predictor of hybrid performance [
23,
24,
25,
26]. The relative contributions of the GCA and SCA effects to the performance of hybrids depend on traits and crosses; for aflatoxin and grain yield [
27], the GCA effect is more important than the SCA effect. However, crosses with higher SCA effects were useful for obtaining high-performing hybrids. Therefore, hybrid performance may depend on the GCA and SCA effects, and different traits may be dominated by either of the combining abilities (GCA or SCA). In maize, heterosis correlates with the genetic distance between the parental inbred lines and increasing genetic distances (variability) between parents increases heterosis [
2,
28]. Heterosis is also related to the heterogeneity and diversity of the parents. The general combining ability is mainly determined by the additive effect of the parental genes, SCA is controlled by dominance and nonallelic gene interactions, and heterosis is mainly caused by dominance and nonallelic interactions, i.e., heterosis mainly depends on the effect of SCA. Therefore, the strength of the SCA in a given cross plays a decisive role in determining the strength of heterosis on the cross’ yield [
29] and quality [
30]. Zhou et al. also have found that the SCA effect in rice is very important for yield heterosis: the loci
Sd1,
Ghd7,
Ghd8, and
DEP1 show a strong GCA effect on many agronomic traits, though not on yield or the seed setting rate; and the QTLs
Ghd8,
S5 and
qS12 show strong SCA effects on yield and thus overdominance, which also indicates that different traits may rely on different types of combining ability [
31]. Since there is no direct relationship between the additive effect of genes and dominance or epistasis, high-yield combinations may be produced in combinations with high GCA or high SCA. Breeding practice shows that the GCA levels of potential crossing parents should be determined beforehand to avoid blind selection, and the selected parents should have high levels of SCA so that high-quality crosses can be obtained.
Although ubiquitous, heterosis does not necessarily occur in every hybridization between two parents or materials, and good crosses do not necessarily come from good parents. Given this variance, the degrees of heterosis in different crosses or different traits within the same cross also vary. Therefore, selecting parents based on past performance does not always produce the desired outcome [
32]; the parents need to be judged based on their potential to produce excellent hybrids rather than on their own performance [
33]. To identify parents with high crossing potential, their combining abilities should be assessed, and ideal parents and crosses can be selected on this basis. Assessing the strength of heterosis requires determining the portion of the genetic effects of heterosis that are utilizable but not easily fixed between different crosses or traits while evaluating their practical value in heterosis breeding, and this assessment provides a basis for parental choice [
16]. Thus, we chose 10 highly inbred homozygous tomato parents with diverse traits as the test material and made 45 crosses by adopting the Griffing IV complete diallel design (P(P − 1)/2), and in order to guarantee the commodity value of the selected combination, we used TY-301 as the control. We analyzed the combining ability, heritability, and heterosis of eight traits, i.e., plant height, early yield, total yield, fruit weight, fruit number per plant, first ripening stage, fruit hardness, and soluble solid content, of the parents and their hybrid progeny. Through this experimental approach, we aimed to screen for crosses with high levels of heterosis and usability, as well as parental materials with good overall performance, thus providing high-quality germplasm resources for studies on the mechanism of heterosis and to promote the breeding of tomato hybrids.
4. Discussion
Evaluating combining ability and heterosis is the first step in breeding inbred lines to develop commercial hybrids [
36]. The genetic bases of heterosis and combining ability remain unclear [
37], but this lack of clarity does not negate the importance of heterosis and combining ability in crop breeding. In the utilization of heterosis, the level of combining ability is an important basis for the choice of crossing parents and directly affects the quality of the hybrid [
38,
39]. Combining ability is measured via two genetic parameters, GCA and SCA, which may be respectively controlled by the additive genetic effects and nonallelic interactions of the parents [
40]. The individual traits of different parents have different GCA values, while the SCA values of different parental crosses also differ, suggesting that the additive and the nonadditive genetic effects are fundamentally different. In this study, we found that for all traits, the variances in the GCA of the parents and the SCA of the crosses were all statistically significant and that the 2σ
2GCA/2σ
2GCA+σ
2SCA ratio was close to 1. These findings indicate that the traits are affected by the joint action of the additive and nonadditive effects of the genes, and the additive effect of the genes plays a dominant role in the inheritance of the traits [
29].
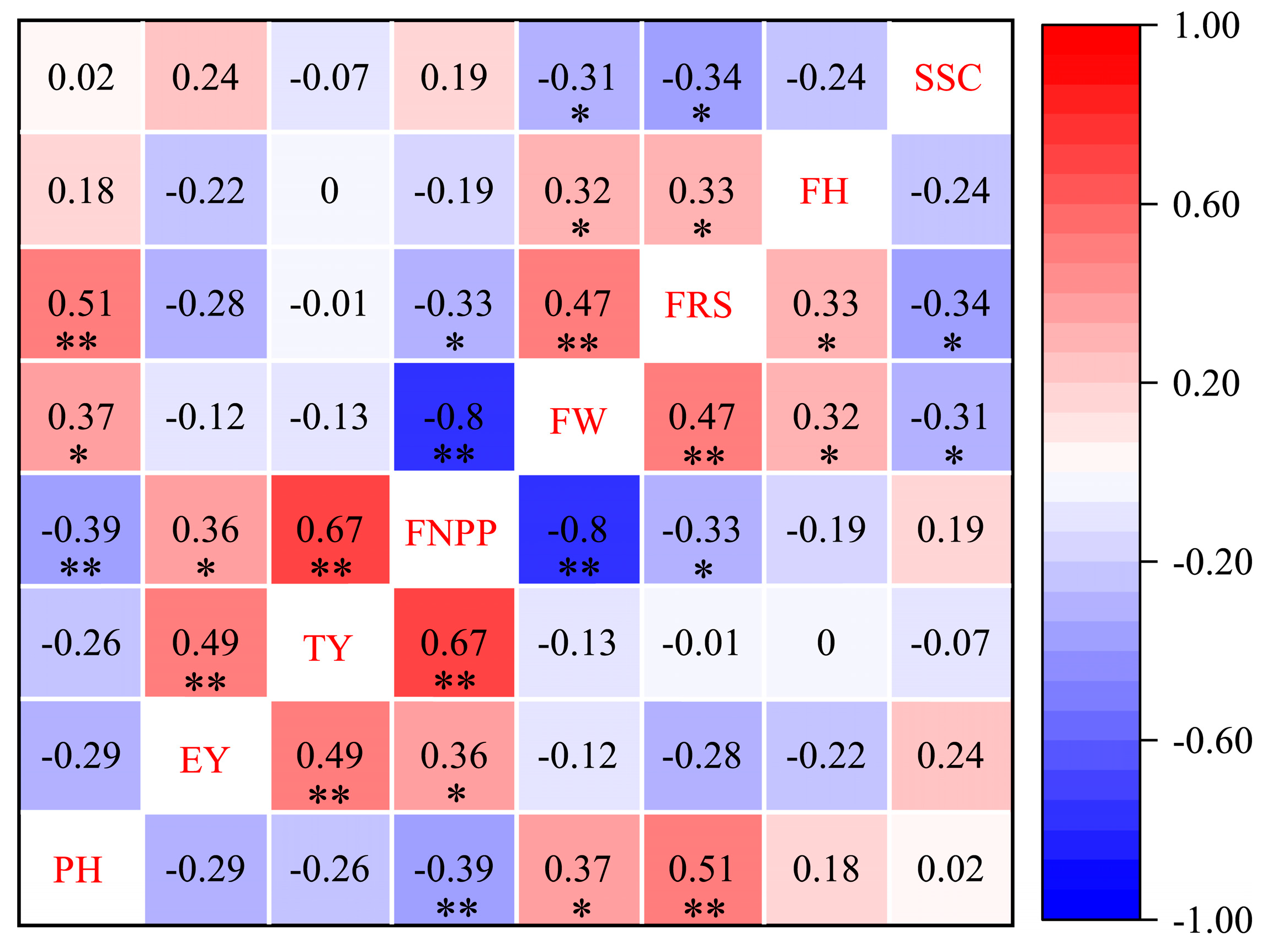
Integrating combining ability, phenotypic performance, and heterosis helps identify crosses with comparatively high levels of heterosis and thus provides valuable insights for crop improvement. For most traits, the GCA effect is strongly correlated with hybrid performance [
23], which is inconsistent with the results of this study. The present study showed that the relationship between the GCA effect and hybrid performance was not statistically significant, but the relationship between the sum of parental GCA values and hybrid performance was generally stronger than that between the SCA effect and hybrid performance, except in the cases of the EY and SSC. Therefore, the sum of parental GCA values plays a comparatively important role in predicting hybrid performance in tomato breeding. Heterosis is mainly caused by dominance effects and nonallelic interactions. Therefore, SCA is highly important for heterosis. We found that for eight traits, GCA and SCA did not correlate, which is consistent with the findings of Zhang et al. [
17] in rice and Han et al. [
18] in soybean. SCA can be used to predict MPH; this conclusion is also supported by previous studies [
41,
42,
43]. The results show that F
1 heterosis may not directly correlate with parental performance. Heterosis depends on the nature of genetic variation [
22]. However, heterosis may depend on the sum of the parents’ GCAs and SCA. Through large-scale analysis of combining ability and heterosis of a hybrid maize population, Yu et al. found that the sum of parental GCAs was either negatively correlated or not correlated with heterosis [
23]. This finding was inconsistent with the conclusion of the present study, which found that the sum of parental GCA values has a highly significant positive correlation with heterosis for some traits. Combining ability and heterosis relationship is more complex, the establishment of heterotic group might be more help to research heterosis. The utilization of heterosis is the theory basis of the heterotic group and heterosis model, the division of heterotic group and the determination of heterosis pattern by many research results show that for corn, rice, and other crops, there is an obviously improved breeding efficiency, speeding up the progress of the breeding effect [
44,
45]. The tomato heterotic group division and establishment has been the subject of a few studie; Jin et al. attempted tomato heterosis group division and the establishment of a heterotic group [
46]. This will be the future research emphasis.
The early maturity and yield of tomato show significant heterosis [
1], and the combination of high yield and early maturity can be obtained through hybridization. This result is consistent with our finding that the EY of most crosses and the TY of all crosses exhibited a positive MPH, while the FRS of most crosses showed a negative MPH. The EY and FRS are important indicators of early maturity in tomato [
47]. We found that the variation coefficient of the FRS was low, suggesting that the FRS was relatively stable in all crosses, which is consistent with the result of a previous study showing that a few hybrids may be superior to their parents and their maturity date advances by only 1–5 days [
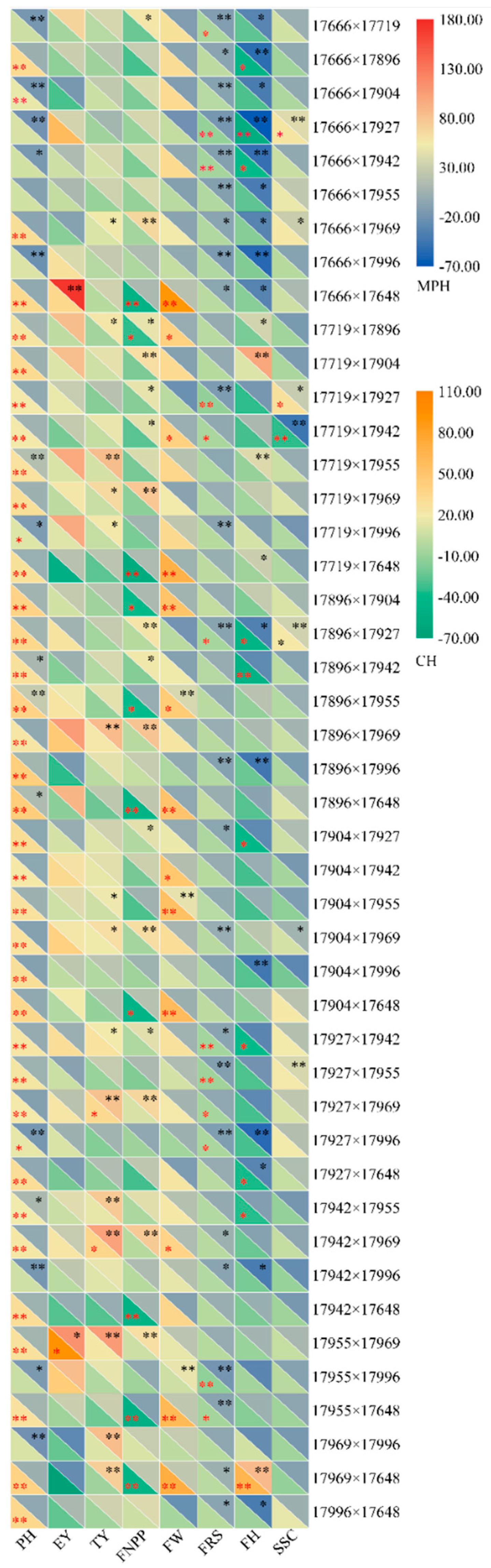
48]. The EY is very important, since it can strongly increase economic efficiency. Therefore, the 17,955 × 17,969 cross, with EY heterosis, would be more valuable to production than the 17,955 × 17,996 cross, which shows heterosis for FRS. In terms of GCA value, 17,969 was superior to 17,996, also indicating that the increase in the yield of the hybrid may result from choosing high-yield parents [
49]; this result was also observed in the analysis of heterosis with respect to yield, i.e., 17,942 × 17,969 and 17,927 × 17,969. The EY of tomato negatively correlated with PH and FW but significantly positively correlated with the FNPP, which was consistent with previous results [
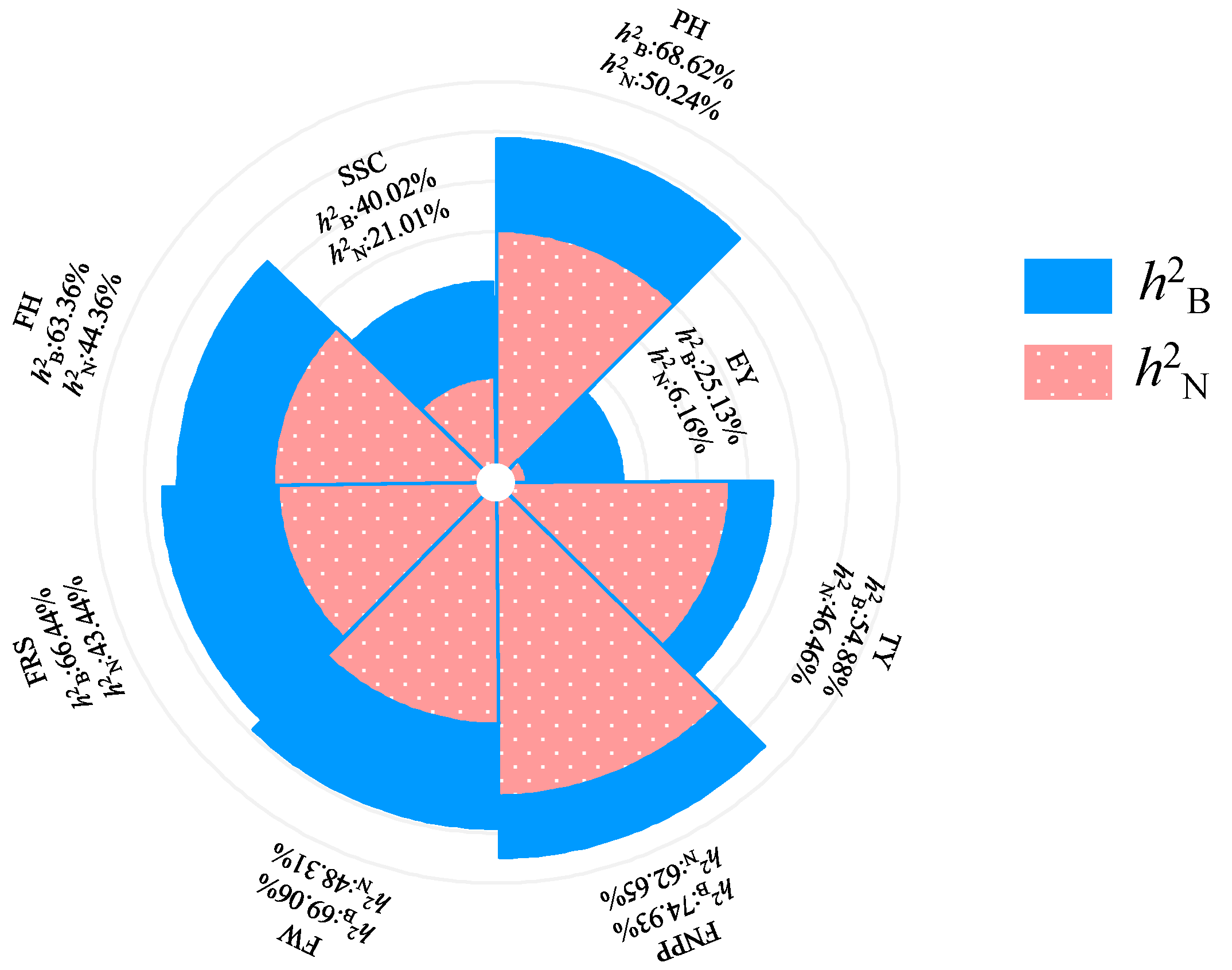
50]. Therefore, based on PH, FW, and FNPP, we can preliminarily predict the level of the EY. Offspring selection is one of the most important stages in plant breeding. Previous studies have shown that early yield is a trait with low heritability [
51,
52,
53], which is consistent with our research results, indicating that it was strongly affected by the environment. In pedigree breeding, selection based on EY is not reliable in early generations, but we found that the heritability of the FNPP was high and correlated closely with EY. Therefore, FNPP can be used as an indirect indicator to improve selection efficiency. This result is inconsistent with that of a previous study that found that the EY of tomato is not significantly associated with the environment and that FNPP is ideally used as a selection indicator in late generations [
50]; this difference is likely because the two studies used different F
1 populations and environments.
The relationship between the components of yield and early maturity is rather complicated. It is difficult to comprehensively balance these factors so that the hybrid has a combination of desirable traits. Nonetheless, the present study has provided an opportunity to select a complex trait through an associated simple trait. To be commercially advantageous, hybrids must be superior to their parents in terms of agronomic traits, especially traits related to yield. Rice and tomato are similar in that most heterotic phenotypes are related to yield [
54,
55]. In this study, we found that EY had a highly significantly positive correlation with Ty, which is consistent with a previous result [
56], indicating that increasing EY helps increase TY. The FNPP had a highly significantly positive correlation with TY and a significantly positive correlation with EY, indicating that heterosis for FNPP is an important indicator for improving yield potential [
29]. The contribution of FNPP to TY is greater than that of EY, which is likely because EY affects TY through the FNPP while also having a direct negative effect on the yield per plant [
57]. In some cases, the FNPP is often negatively affected by other traits, such as FW. Therefore, when selecting based on EY, we should pay attention to the FNPP, especially when breeding tomato for high yield. The improvement in FNPP-related traits is an effective way to breed for heterosis for yield. The results and inferences of this study are only based on data from one year and one environment. In order to be more reliable and accurate, the repeated work for many years and in multiple environments will be the focus of the next step.
5. Conclusions
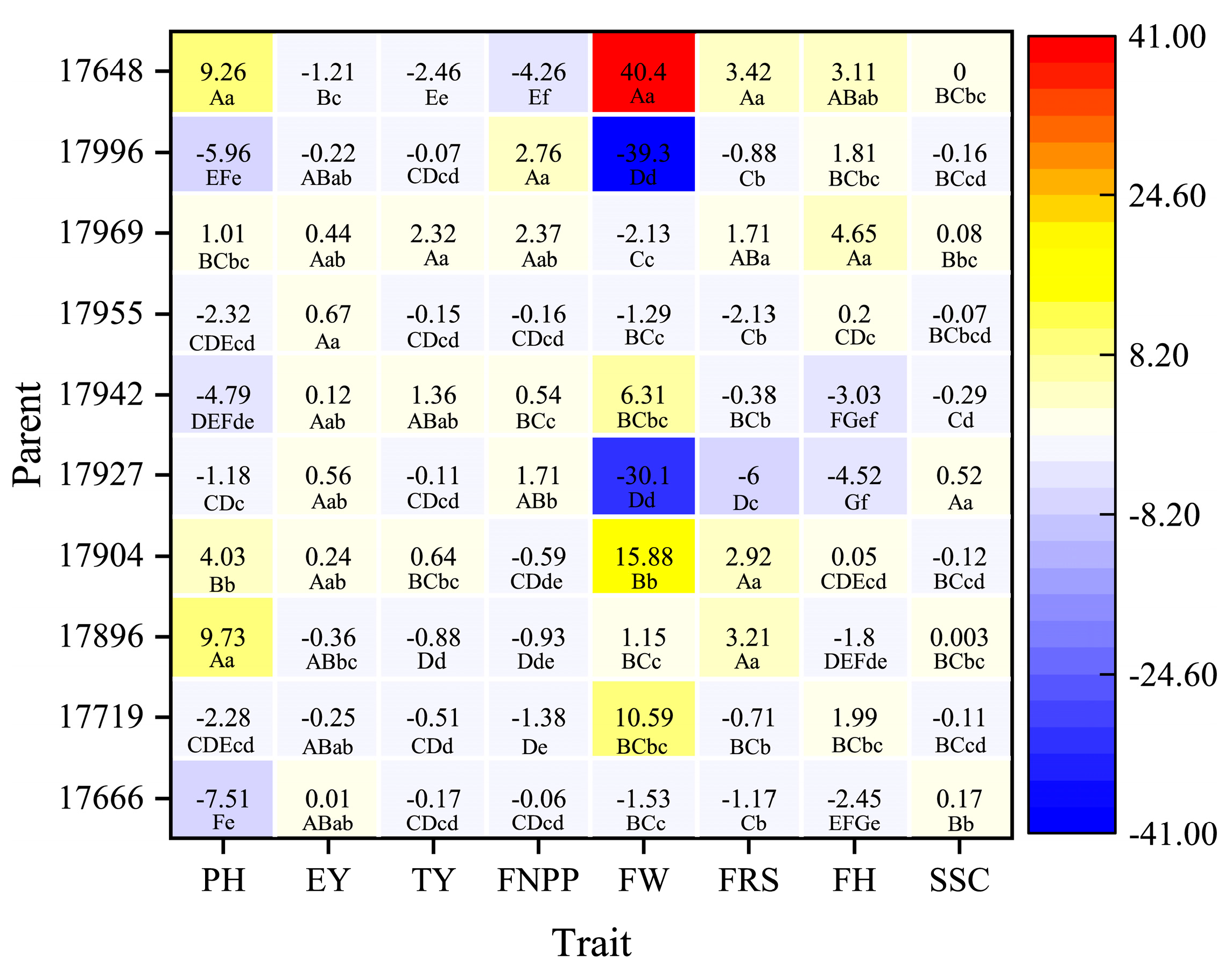
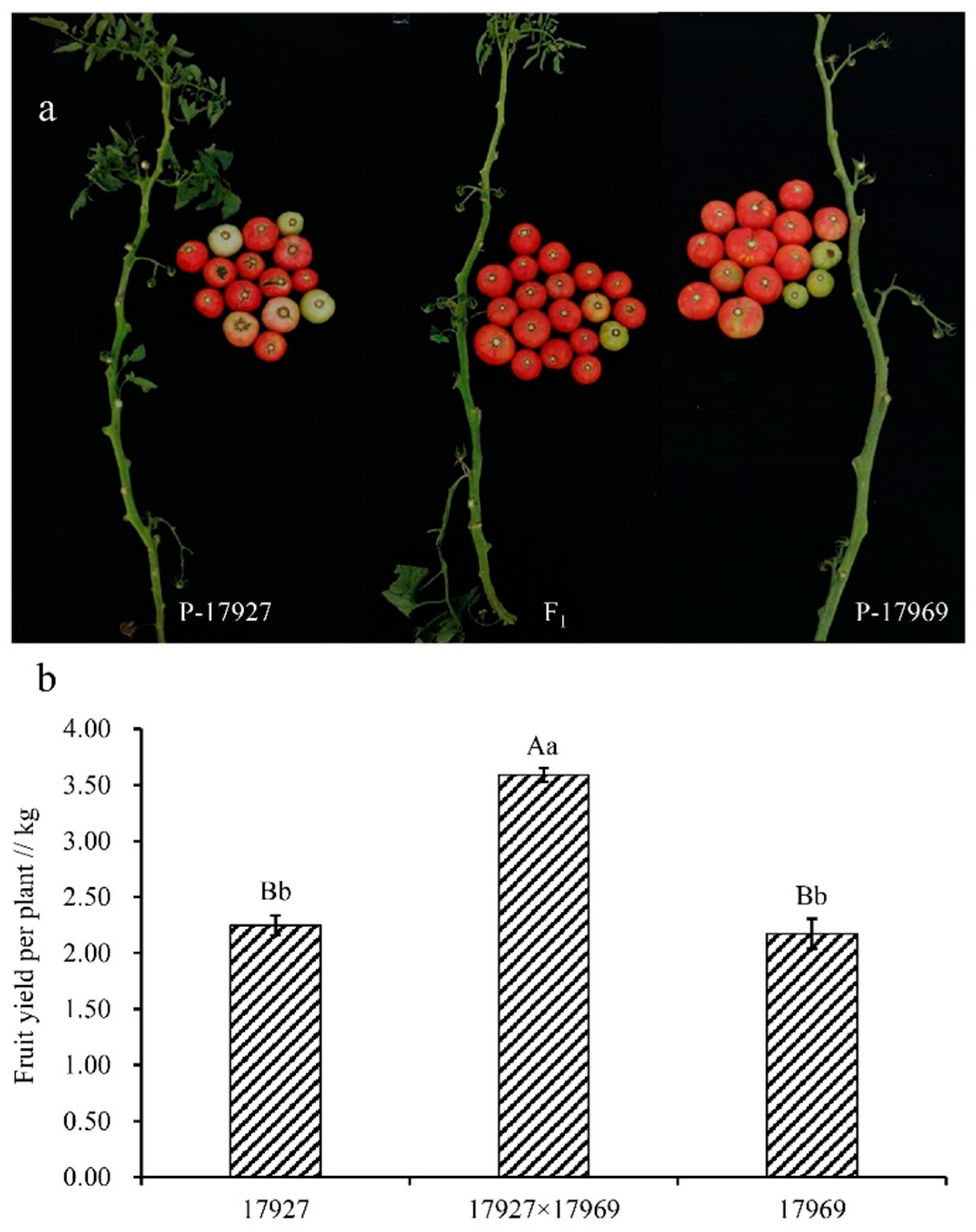
The present results showed clear heterosis for precocity and yield in tomato. Both additive and nonadditive genetic effects were involved in the expression of the traits, and the additive genetic effect was dominant in trait inheritance. Although GCA and SCA were not correlated, and the strength of heterosis depends on SCA, GCAsum did predict heterosis for some traits with higher predictive accuracy than did SCA. Compared with heterosis, GCAsum can better predict hybrid performance. Parent 17,969 showed the best overall trait performance, especially in terms of yield. Parents 17,955 and 17,927 showed strong potential for early maturity. The overall characteristics were higher plant height, early maturity, lower early yield, and slightly worse hardness. The best cross for yield per plot and fruit number per plant was 17,927 × 17,969, and the best for fruit ripening stage and soluble solid content was 17,719 × 17,927.

 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}