
A New Identification Method Reveals the Resistance of an Extensive-Source Radish Collection to Plasmodiophora brassicae Race 4

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Plant Materials
2.2. Pathogen Preparation
2.3. Plant Cultivation and Treatment
2.4. Data Collection
2.5. Data Analysis
3. Results
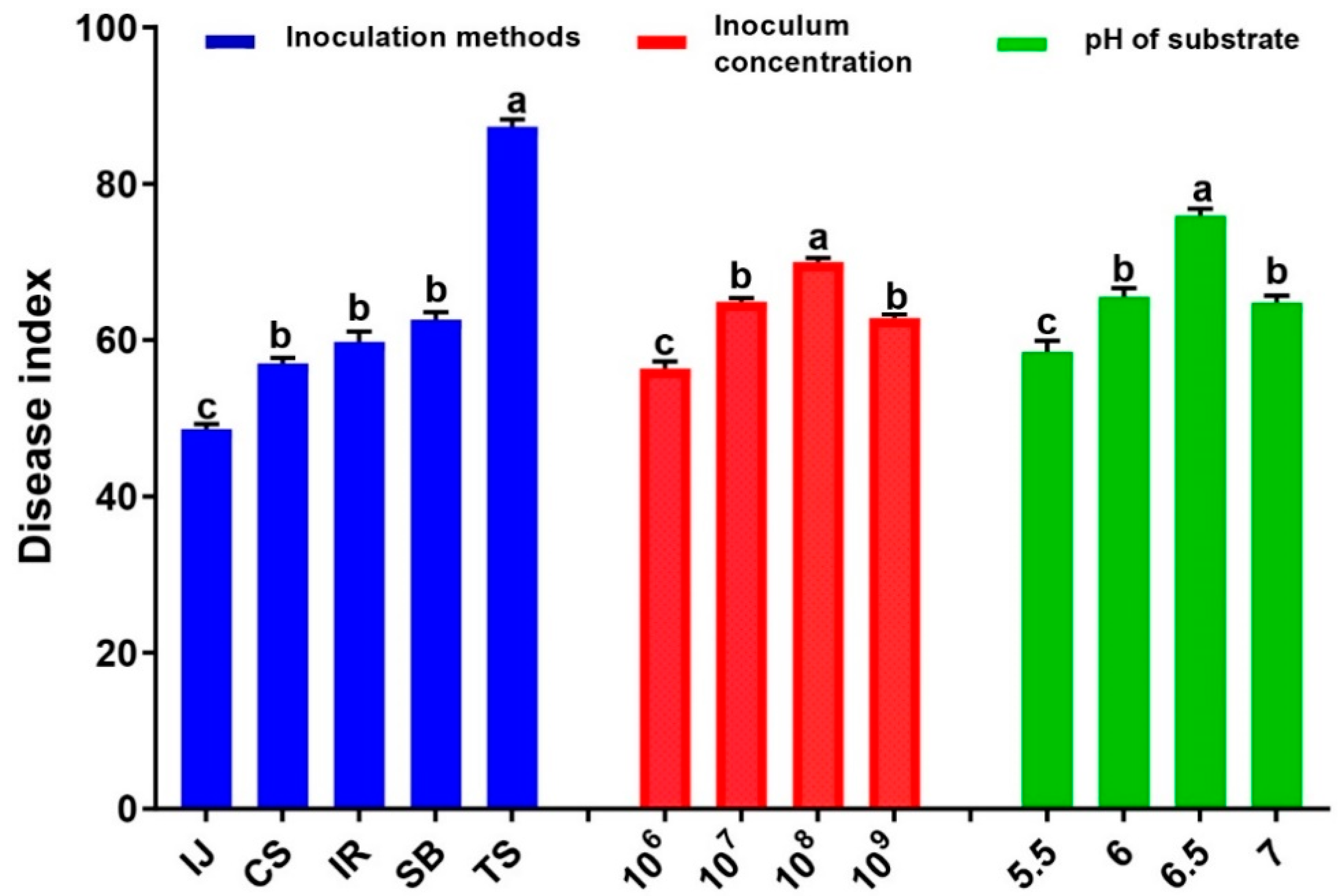
3.1. Influence of Different Inoculum Concentrations, Inoculation Methods, and pH of Substrate on Severity of Clubroot Disease in Radish
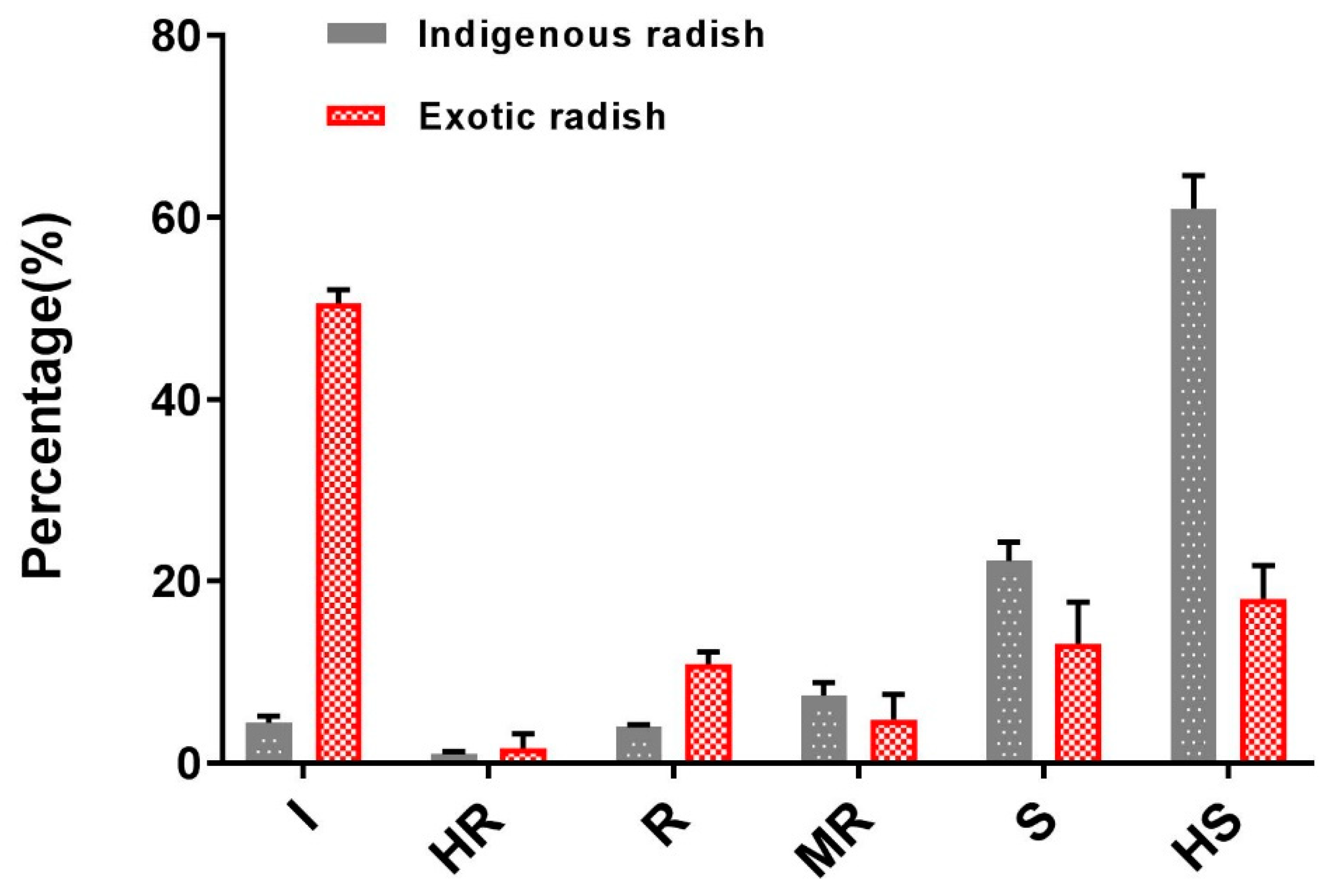
3.2. The Resistance of Radish Germplasms to P. brassicae Race 4
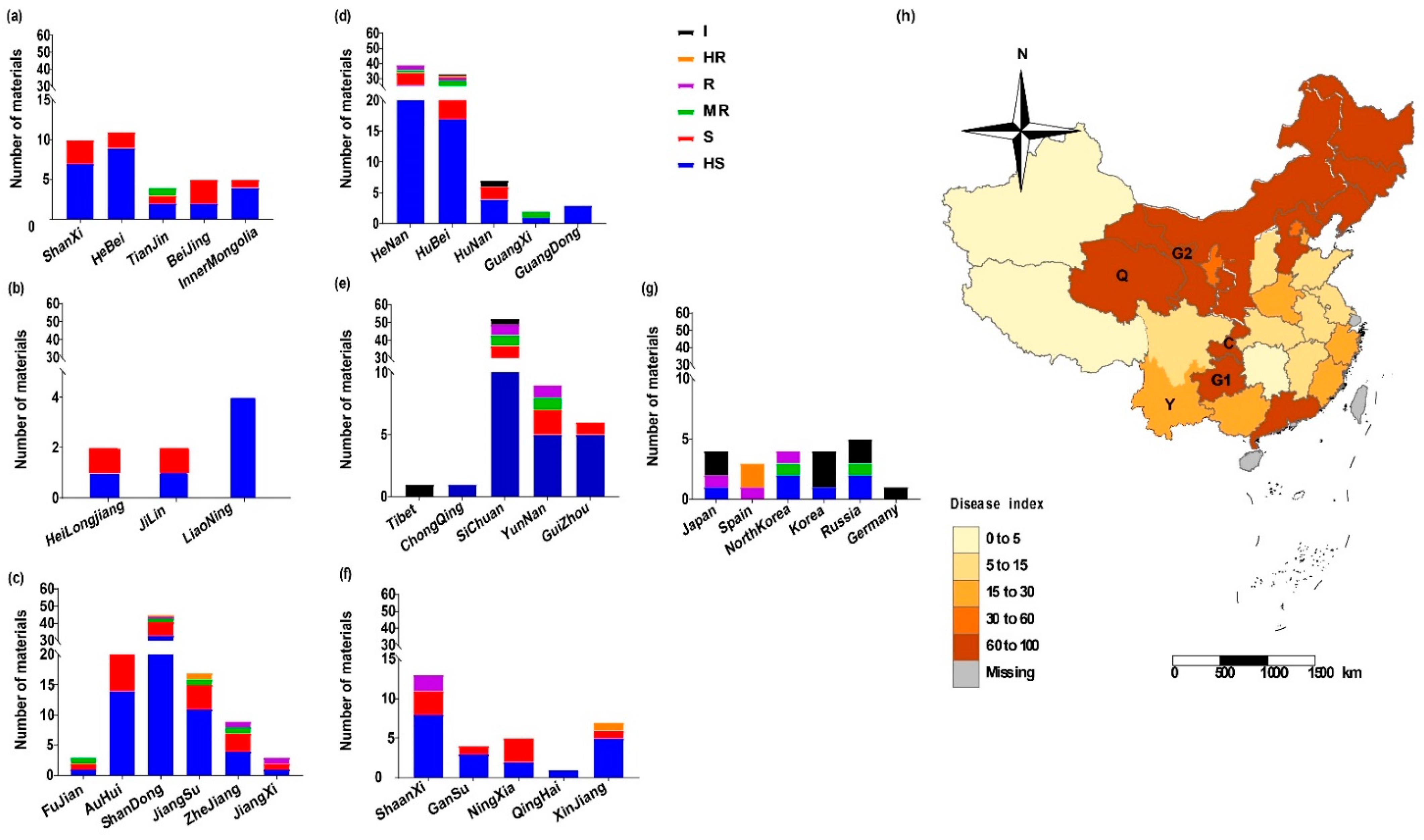
3.3. Geographic Distribution of the Radish Resources with Different Resistance to P. brassicae Race 4
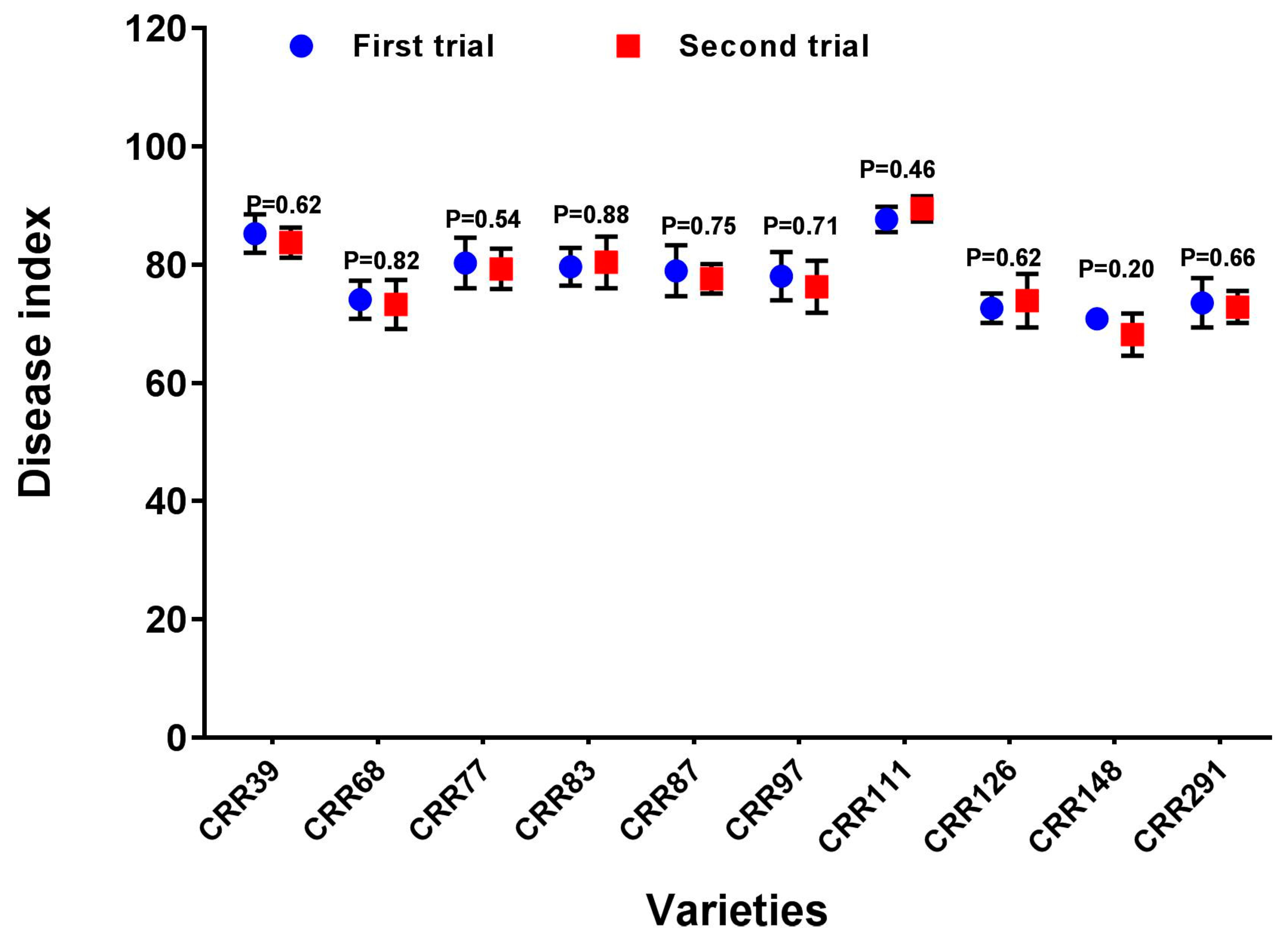
3.4. Verification of the New Method by Repeated Identification of the Resistance of Immune Materials and Representative Highly Susceptible Materials
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hirai, M. Genetic analysis of clubroot resistance in Brassica crops. Breed. Sci. 2006, 56, 223–229. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Huang, Y.L.; Li, X.L.; Li, H.Z. Research progress in clubroot of crucifers. Plant Prot. 2011, 37, 153–158. [Google Scholar]
- Chen, S.L.; Li, Y.L.; Wang, M.X.; Wang, P.S.; Ren, G.C.; Yi, Z.H. Occurrence of clubroot disease of radish in Lushan county. China Plant Prot. 2001, 1, 20. [Google Scholar]
- Zhang, R.B. Investigation on Plasmodiophora brassica in Chendu and Hungicides Screening in Field. Master’s Thesis, Sichuan Agricultural University, Chengdu, China, 2010. [Google Scholar]
- He, J.M.; Hu, J.F.; Dai, Y.J.; Wang, Q.; Wu, L.Y.; Li, S.K.; Li, W.F.; He, Z.R.; Yang, H.L.; Zhong, L. Breeding of club-root resistant ogura CMS backcross line and maintainer line for Brassica Campestris. Hunan Agric. Sci. 2011, 23, 1–5. [Google Scholar]
- Zhang, Z.M.; Zhao, Y.T. Identification of new symptom of clubroot disease in white radish. China Plant Prot. 2013, 32, 44–45. [Google Scholar]
- Matsumoto, E.; Yasui, C.; Ohi, M.; Tsukada, M. Linkage analysis of RFLP markers for clubroot resistance and pigmentation in Chinese cabbage. Euphytica 1998, 104, 79–86. [Google Scholar] [CrossRef]
- Sakamoto, K.; Saito, A.; Hayashida, N.; Taguchi, G.; Matsumoto, E. Mapping of isolate-specific QTL for clubroot resistance in Chinese cabbage (Brassica rapa L. ssp. pekinensis). Theor. Appl. Genet. 2008, 117, 759–767. [Google Scholar] [CrossRef]
- Nikolaev, S.I.; Berney, C.; Fahrni, J.F.; Bolivar, I.; Polet, S.; Mylnikov, A.P.; Aleshin, V.V.; Petrov, N.B.; Pawlowski, J. The twilight of Heliozoa and rise of Rhizaria, an emerging supergroup of amoeboid eukaryotes. Proc. Natl. Acad. Sci. USA 2004, 101, 8066–8071. [Google Scholar] [CrossRef] [Green Version]
- Hatakeyama, K.; Suwabe, K.; Tomita, R.N.; Kato, T.; Nunome, T.; Fukuoka, H.; Matsumoto, S. Identification and characterization of Crr1a, a gene for resistance to clubroot disease (Plasmodiophora brassicae Woronin) in Brassica rapa L. PLoS ONE 2013, 8, e54745. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Zhang, X.; Peng, G.; Falk, K.C.; Strelkov, S.E.; Gossen, B.D. Genotyping-by-sequencing reveals three QTL for clubroot resistance to six pathotypes of Plasmodiophora brassicae in Brassica rapa. Sci. Rep. 2017, 7, 4516. [Google Scholar] [CrossRef]
- Nguyen, M.L.; Monakhos, G.F.; Komakhin, R.A.; Monakhos, S.G. The new clubroot resistance locus is located on chromosome A05 in Chinese cabbage (Brassica rapa L.). Russ. J. Genet. 2018, 54, 296–304. [Google Scholar] [CrossRef]
- Ingram, D.S.; Tommerup, I.C. The life history of Plasmodiophora brassicae Woron. Proc. R. Soc. B Biol. Sci. 1972, 180, 103–112. [Google Scholar]
- McDonald, M.R.; Sharma, K.; Gossen, B.D.; Deora, A.; Feng, J.; Hwang, S.F. The role of primary and secondary infection in host response to Plasmodiophora brassicae. Phytopathology 2014, 104, 1078–1087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voorrips, R.E.; Jongerius, M.C.; Kanne, H.J. Quantitative trait loci for clubroot resistance in Brassica oleracea. In Brassicas and Legumes from Genome Structure to Breeding. Biotechnology in Agriculture and Forestry; Nagata, T., Tabata, S., Eds.; Springer: Berlin/Heidelberg, Germany, 2003; Volume 52, pp. 87–104. [Google Scholar]
- Pageau, D.; Lajeunesse, J.; Lafond, J. Impact of clubroot (Plasmodiophora brassicae) on the yield and quality of canola. Can. J. Plant Pathol. 2006, 28, 137–143. [Google Scholar] [CrossRef]
- Buczacki, S.T.; Ockendon, J.G.; White, J.G. General studies. In Report of the National Vegetable Research Station for 1974; 1974; p. 113. [Google Scholar]
- Wallenhammar, A.C. Prevalence of Plasmodiophora brassicae in spring oil-seed rape growing area in Central Sweden and factors influencing soil infestation levels. Plant Pathol. 1996, 45, 710–719. [Google Scholar] [CrossRef]
- Gossen, B.D.; Strelkov, S.E.; Manolii, V.P.; Rennie, D.C.; Cao, T.; Hwang, S.F.; Peng, G.; McDonald, M.R. Spread of Plasmodiophora brassicae on canola in Canada, 2003–2014: Old pathogen, new home. Can. J. Plant Pathol. 2015, 37, 403–413. [Google Scholar] [CrossRef]
- Hwang, S.F.; Howard, R.J.; Strelkov, S.E.; Gossen, B.D.; Peng, G. Management of clubroot (Plasmodiophora brassicae) on canola (Brassica napus) in western Canada. Can. J. Plant Pathol. 2014, 36, 49–65. [Google Scholar] [CrossRef]
- Botero, A.; García, C.; Gossen, B.D.; Strelkov, S.E.; Todd, C.D.; Bonham-Smith, P.C.; Pérez-López, E. Clubroot disease in Latin America: Distribution and management strategies. Plant Pathol. 2019, 68, 827–833. [Google Scholar] [CrossRef] [Green Version]
- Donald, C.; Porter, I. Integrated control of clubroot. J. Plant Growth Regul. 2009, 28, 289. [Google Scholar] [CrossRef]
- Hwang, S.F.; Ahmed, H.U.; Strelkov, S.E.; Zhou, Q.; Gossen, B.D.; McDonald, M.R.; Peng, G.; Turnbull, G.D. Suppression of clubroot by dazomet fumigant. Can. J. Plant Sci. 2017, 98, 172–182. [Google Scholar] [CrossRef]
- Myers, D.F.; Campbell, R.N.; Greathead, A.S. Clubroot of crucifers in California: Soils respond differently to lime for clubroot control. Phytopathology 1981, 71, 1005–1006. [Google Scholar]
- Gossen, B.D.; Kasinathan, H.; Cao, T.; Manolii, V.P.; Strelkov, S.E.; Hwang, S.F.; McDonald, M.R. Interaction of pH and temperature on infection and symptom development of clubroot in canola. Can. J. Plant Pathol. 2013, 35, 294–303. [Google Scholar] [CrossRef]
- Peng, G.; Pageau, D.; Strelkov, S.E.; Gossen, B.D.; Hwang, S.F.; Lahlali, R. A > 2-year crop rotation reduces resting spores of Plasmodiophora brassicae in soil and the impact of clubroot on canola. Eur. J. Agron. 2015, 70, 78–84. [Google Scholar] [CrossRef]
- Ernst, T.W.; Kher, S.; Stanton, D.; Rennie, D.C.; Hwang, S.F.; Strelkov, S.E. Plasmodiophora brassicae resting spore dynamics in clubroot resistant canola (Brassica napus) cropping systems. Plant Pathol. 2019, 68, 399–408. [Google Scholar] [CrossRef]
- Gossen, B.D.; Al-Daoud, F.; Dumonceaux, T.; Dalton, J.A.; Peng, G.; Pageau, D.; McDonald, M.R. Comparison of techniques for estimation of resting spores of Plasmodiophora brassicae in soil. Plant Pathol. 2019, 68, 954–961. [Google Scholar] [CrossRef]
- Peng, G.; McGregor, L.; Lahlali, R.; Gossen, B.D.; Hwang, S.F.; Adhikari, K.K.; Strelkov, S.E.; McDonald, M.R. Potential biological control of clubroot on canola and crucifer vegetable crops. Plant Pathol. 2011, 60, 566–574. [Google Scholar] [CrossRef]
- Wit, F.; Weg, M. Clubroot-resistance in turnips (Brassica campestris L.). Euphytica 1964, 13, 9–18. [Google Scholar] [CrossRef]
- Tjallingii, F. Testing clubroot-resistance of turnips in the Netherlands and the physiologic specialization of Plasmodiophora brassicae. Euphytica 1965, 14, 1–22. [Google Scholar]
- Scholze, P.; Hammer, K. Evaluation of resistance to Plasmodiophora brassicae, Alternaria and Phoma in Brassicaceae. ISHS Acta Horticulturae 459. In Proceedings of the International Symposium Brassica, Rennes, France, 23–27 September 1997; pp. 363–372. [Google Scholar]
- Ren, L.; Wang, J.; Jia, J.G.; He, Y.Q.; Liu, S.Y.; Fang, X.P. Preliminary study on resistance of cruciferous crops to clubroot disease. In Agricultural Reformation and Rural Progressing: Annual Conference Proceedings of China Agriculture System Engineering Society; Konglai, Z., Zhang, H., Eds.; Aussino Academic Publishing House: Sydney, Australia, 2010. [Google Scholar]
- Sharma, P.; Siddiqui, S.A.; Rai, P.K.; Meena, P.D.; Kumar, J.; Chauhan, J.S. Evaluation of Brassica germplasm for field resistance against clubroot (Plasmodiophora brassicae woron). Arch. Phytopathol. Plant Prot. 2012, 45, 356–359. [Google Scholar] [CrossRef]
- Tan, C.; Yue, Y.L. Identification of different Chinese cabbage [Brassica campestris L. ssp. pekinensis (Lour) Olsson] varieties resistance to clubroot (Plasmodiophora brassicae). China Veget 2013, 8, 91–94. [Google Scholar]
- Huang, X.L.; Ran, Z.H.; Peng, S.S.; Liu, M.J.; Xun, L.F.; Liu, E.M. Identification and evaluation of resistance of Chinese cabbage in Hunan province against clubroot disease. J. Hunan Agric. Univ. Nat. Sci. 2014, 40, 43–47. [Google Scholar]
- Shi, Q.N. Study on Screening of Clubroot Resistance Germplasm Resources and Hybrid Combinations of Chinese Cabbage. Master’s Thesis, Northwest A&F University, Shanxi, China, 2016. [Google Scholar]
- Crisp, P.; Crute, I.R.; Sutherland, R.A.; Angell, S.M.; Bloor, K.; Burgess, H.; Gordon, P.L. The exploitation of genetic resources of Brassica oleracea in breeding for resistance to clubfoot (Plasmodiophora brassicae). Euphytica 1989, 42, 215–226. [Google Scholar]
- Manzanares-Dauleux, M.J.; Divaret, I.; Baron, F.; Thomas, G. Evaluation of French Brassica oleracea landraces for resistance to Plasmodiophora brassicae. Euphytica 2000, 113, 211–218. [Google Scholar] [CrossRef]
- Carlsson, M.; Bothmer, R.V.; Merker, A. Screening and evaluation of resistance to downy mildew (Peronospora parasitica) and clubroot (Plasmodiophora brassicae) in genetic resources of Brassica oleracea. Hereditas 2004, 141, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Si, J.; Li, C.Q.; Song, H.Y.; Ren, X.S.; Song, M.; Wang, X.J. Identification and evaluation of resistance to clubroot in cabbage (Brassica oleracea var. Capitata L.). J. Southwest Univ. Nat. Sci. 2009, 31, 26–30. [Google Scholar]
- Hasan, M.J.; Strelkov, S.E.; Howard, R.J.; Rahman, H. Screening of Brassica germplasm for resistance to Plasmodiophora brassicae pathotypes prevalent in Canada for broadening diversity in clubroot resistance. Can. J. Plant Sci. 2012, 92, 501–515. [Google Scholar] [CrossRef] [Green Version]
- Ma, D.D.; Pang, S.Q.; Ding, Y.H.; Jian, Y.C.; Geng, L.H. Study on clubroot resistance of different Brassica crops. J. Shihezi Univ. Nat. Sci. 2016, 34, 164–169. [Google Scholar]
- Sun, C.; Ma, J.; Lei, L.; Liu, J.; Xie, J.M.; Kang, J.G. Identification of cabbage germplasm resources with resistance to clubroot and sequence analysis of CRa and Crr1a homologous genes from cabbage. J. Plant Genet. Resour. 2016, 17, 1058–1064. [Google Scholar]
- Karling, J.S. The Plasmodiophorales: Including a Complete Host Index, Bibliography, and a Description of Diseases Caused by Species of this Order, 2nd ed.; Hafner Publishing Company: New York, NY, USA, 1968. [Google Scholar]
- Dixon, G.R.; Doodson, J.K.; Beeney, B.W.; Davies, H.; Jemmett, J.L.; Moxon, R.H. Studies of resistance in swede seed stocks to clubroot (Plasmodiophora brassicae). Plant Var. Seeds 1972, 12, 456–463. [Google Scholar]
- Vigier, M.S.; Hum, D.J. Source of resistance to clubroot (Plasmodophom brassicae Wor.) in triazine-resistant spring canola (rapeseed). Can. Plant Dis. Surv. 1989, 69, 113. [Google Scholar]
- Gao, Y.; Gao, F.; Li, S.; Peng, S.; Chen, S.; Lin, L. Screening clubroot-resistant germplasm resources from 20 Brassica napus varieties. J. Yunnan Agric. Univ. 2015, 30, 346–356. [Google Scholar]
- Liu, Y.; Xu, A.; Liang, F.; Yao, X.; Wang, Y.; Liu, X.; Zhang, Y.; Dalelhan, J.; Zhang, B.; Qin, M.; et al. Screening of clubroot-resistant varieties and transfer of clubroot resistance genes to Brassica napus using distant hybridization. Breed Sci. 2018, 68, 258–267. [Google Scholar] [CrossRef] [Green Version]
- Liang, J.H.; Lang, L.N.; Liu, Y.P.; Wang, J.; Zhang, H.; Xu, A.X.; Huang, Z. Resource screening and germplasm creation of Brassica napus resistant to clubroot. Acta Agric. Boreali-Sin. 2019, 34, 219–225. [Google Scholar]
- Yoshikawa, H. Breeding for clubroot resistance in Chinese cabbage. In Proceedings of the International Symposium on Chinese Cabbage, Tsukuba, Japan, 31 March–5 April 1980; pp. 405–413. [Google Scholar]
- Suwabe, K.; Tsukazaki, H.; Iketani, H.; Hatakeyama, K.; Fujimura, M.; Nunome, T.; Fukuoka, H.; Matsumoto, S.; Hirai, M. Identification of two loci for resistance to clubroot (Plasmodiophora brassicae Woronin) in Brassica rapa L. Theor. Appl. Genet. 2003, 107, 997–1002. [Google Scholar] [CrossRef] [PubMed]
- Suwabe, K.; Tsukazaki, H.; Iketani, H.; Hatakeyama, K.; Kondo, M.; Fujimura, M.; Nunome, T.; Fukuoka, H.; Hirai, M.; Matsumoto, S. Simple sequence repeat-based comparative genomics between Brassica rapa and Arabidopsis thaliana: The genetic origin of clubroot resistance. Genetics 2006, 173, 309–319. [Google Scholar] [CrossRef] [Green Version]
- Hirai, M.; Harada, T.; Kubo, N.; Tsukada, M.; Suwabe, K.; Matsumoto, S. A novel locus for clubroot resistance in Brassica rapa and its linkage markers. Theor. Appl. Genet. 2004, 108, 639–643. [Google Scholar] [CrossRef] [PubMed]
- Piao, Z.Y.; Deng, Y.Q.; Choi, S.R.; Park, Y.J.; Lim, Y.P. SCAR and CAPS mapping of CRb, a gene conferring resistance to Plasmodiophora brassicae in Chinese cabbage (Brassica rapa ssp. pekinensis). Theor. Appl. Genet. 2004, 108, 1458–1465. [Google Scholar] [CrossRef]
- Chen, J.; Jing, J.; Zhan, Z.; Zhang, T.; Zhang, C.; Piao, Z. Identification of novel QTLs for isolate-specific partial resistance to Plasmodiophora brassicae in Brassica rapa. PLoS ONE 2013, 8, e85307. [Google Scholar] [CrossRef] [Green Version]
- Chu, M.; Song, T.; Falk, K.C.; Zhang, X.; Liu, X.; Chang, A.; Lahlali, R.; McGregor, L.; Gossen, B.D.; Yu, F.; et al. Fine mapping of Rcr1 and analyses of its effect on transcriptome patterns during infection by Plasmodiophora brassicae. BMC Genom. 2014, 15, 1166. [Google Scholar] [CrossRef] [Green Version]
- Pang, W.; Liang, S.; Li, X.; Li, P.; Yu, S.; Lim, Y.P.; Piao, Z. Genetic detection of clubroot resistance loci in a new population of Brassica rapa. Hortic. Environ. Biotechnol. 2014, 55, 540–547. [Google Scholar] [CrossRef]
- Huang, Z.; Peng, G.; Liu, X.; Deora, A.; Falk, K.C.; Gossen, B.D.; McDonald, M.R.; Yu, F. Fine mapping of a clubroot resistance gene in Chinese cabbage using SNP markers identified from bulked segregant RNA sequencing. Front. Plant Sci. 2017, 8, 1448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, W.; Fu, P.; Li, X.; Zhan, Z.; Yu, S.; Piao, Z. Identification and mapping of the clubroot resistance gene CRd in chinese cabbage (Brassica rapa ssp. pekinensis). Front. Plant Sci. 2018, 9, 653. [Google Scholar] [CrossRef] [Green Version]
- Laila, R.; Park, J.I.; Robin, A.H.; Natarajan, S.; Vijayakumar, H.; Shirasawa, K.; Isobe, S.; Kim, H.T.; Nou, I.S. Mapping of a novel clubroot resistance QTL using ddRAD-seq in Chinese cabbage (Brassica rapa L.). BMC Plant Biol. 2019, 19, 13. [Google Scholar] [CrossRef]
- Zhu, H.; Zhai, W.; Li, X.; Zhu, Y. Two QTLs controlling clubroot resistance identified from bulked segregant sequencing in pakchoi (Brassica campestris ssp. Chinensis Makino). Sci. Rep. 2019, 9, 9228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, S.R.; Oh, S.H.; Chhapekar, S.S.; Dhandapani, V.; Lee, C.Y.; Rameneni, J.J.; Ma, Y.; Choi, G.J.; Lee, S.S.; Lim, Y.P. Quantitative trait locus mapping of clubroot resistance and Plasmodiophora brassicae pathotype banglim-specific marker development in Brassica rapa. Int. J. Mol. Sci. 2020, 21, 4157. [Google Scholar] [CrossRef] [PubMed]
- Tomita, H.; Shimizu, M.; Doullah, M.A.; Fujimoto, R.; Okazaki, K. Accumulation of quantitative trait loci conferring broad-spectrum clubroot resistance in Brassica oleracea. Mol. Breed. 2013, 32, 889–900. [Google Scholar] [CrossRef]
- Landry, B.S.; Hubert, N.; Crete, R.; Chang, M.S.; Lincoln, S.E.; Etoh, T. A genetic map for Brassica oleracea based on RFLP markers detected with expressed DNA sequences and mapping of resistance gene to race 2 of Plasmodiophora brassicae. Genome 1992, 35, 409–420. [Google Scholar] [CrossRef]
- Figdore, S.S.; Ferreira, M.E.; Slocum, M.K.; Williams, P.H. Association of RFLP markers with trait loci affecting clubroot resistance and morphological characters in Brassica oleracea L. Euphytica 1993, 69, 33–44. [Google Scholar] [CrossRef]
- Grandclément, C.; Thomas, G. Detection and analysis of QTL based on RAPD markers for polygenic resistance to Plasmodiophora brassicae Wor. in Brassica oleracea L. Theor. Appl. Genet. 1996, 93, 86–90. [Google Scholar] [CrossRef] [PubMed]
- Voorrips, R.E.; Kanne, H.J. Genetic analysis of resistance to clubroot (Plasmodiophora brassicae) in Brassica oleracea. II. Quantitative analysis of root system measurements. Euphytica 1997, 93, 41–48. [Google Scholar] [CrossRef]
- Moriguchi, K.; Kimizuka-Takagi, C.; Ishii, K.; Nomura, K. A genetic map based on RAPD, RFLP, isozyme, morphological markers and QTL analysis for clubroot resistance in Brassica oleracea. Breed Sci. 1999, 49, 257–265. [Google Scholar] [CrossRef]
- Rocherieux, J.; Glory, P.; Giboulot, A.; Boury, S.; Barbeyron, G.; Thomas, G.; Manzanares-Dauleux, M.J. Isolate-specific and broads pectrum QTL are involved in the control of clubroot in Brassica oleracea. Theor. Appl. Genet. 2004, 108, 1555–1563. [Google Scholar] [CrossRef] [PubMed]
- Nagaoka, T.; Doullah, M.A.; Matsumoto, S.; Kawasaki, S.; Ishikawa, T.; Hori, H.; Okazaki, K. Identification of QTLs that control clubroot resistance in Brassica oleracea and comparative analysis of clubroot resistance genes between B. rapa and B. oleracea. Theor. Appl. Genet. 2010, 120, 1335–1346. [Google Scholar] [CrossRef]
- Lee, J.; Izzah, N.K.; Choi, B.S.; Joh, H.J.; Lee, S.C.; Perumal, S.; Seo, J.; Ahn, K.; Jo, E.J.; Choi, G.J.; et al. Genotyping-by-sequencing map permits identification of clubroot resistance QTLs and revision of the reference genome assembly in cabbage (Brassica oleracea L.). DNA Res. 2016, 23, 29–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, L.; Zhou, L.; Li, Q.; Wei, D.; Ren, X.; Song, H.; Mei, J.; Si, J.; Qian, W. Identification of quantitative trait loci for clubroot resistance in Brassica oleracea with the use of Brassica SNP microarray. Front. Plant Sci. 2018, 9, 822. [Google Scholar] [CrossRef]
- Dakouri, A.; Zhang, X.; Peng, G.; Falk, K.C.; Gossen, B.D.; Strelkov, S.E.; Yu, F. Analysis of genome-wide variants through bulked segregant RNA sequencing reveals a major gene for resistance to Plasmodiophora brassicae in Braassica oleracea. Sci. Rep. 2018, 8, 17657. [Google Scholar] [CrossRef]
- Piao, Z.; Ramchiary, N.; Lim, Y.P. Genetics of clubroot resistance in Brassica species. J. Plant Growth Regul. 2009, 28, 252–264. [Google Scholar] [CrossRef]
- Rahman, H.; Shakir, A.; Hasan, M.J. Breeding for clubroot resistant spring canola (Brassica napus L.) for the Canadian prairies: Can the European winter canola cv. Mendel be used as a source of resistance? Can. J. Plant Sci. 2011, 91, 447–458. [Google Scholar] [CrossRef] [Green Version]
- Werner, S.; Diederichsen, E.; Frauen, M.; Schondelmaier, J.; Jung, C. Genetic mapping of clubroot resistance genes in oilseed rape. Theor. Appl. Genet. 2008, 116, 363–372. [Google Scholar] [CrossRef]
- Li, L.; Luo, Y.; Chen, B.; Xu, K.; Zhang, F.; Li, H.; Huang, Q.; Xiao, X.; Zhang, T.; Hu, J.; et al. A genome-wide association study reveals new loci for resistance to clubroot disease in Brassica napus. Front. Plant Sci. 2016, 7, 1483. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Feng, J.; Hwang, S.F.; Strelkov, S.E.; Falak, I.; Huang, X.; Sun, R. Mapping of clubroot (Plasmodiophora brassicae) resistance in Brassica napus canola. Plant Pathol. 2016, 65, 435–440. [Google Scholar] [CrossRef] [Green Version]
- Scheijgrond, W.; Vos, H. Investigation on the susceptibility to club root. Euphytica 1954, 3, 125–139. [Google Scholar] [CrossRef]
- Yoshikawa, H. Studies on breeding of clubroot resistance in cole crops (in Japanese with English summary). Bull. Natl. Res. Inst. Veg. Ornam. Plants Tea Japan Ser. A 1993, 7, 1–165. [Google Scholar]
- Kamei, A.; Tsuro, M.; Kubo, N.; Hayashi, T.; Wang, N.; Fujimura, T.; Hirai, M. QTL mapping of clubroot resistance in radish (Raphanus sativus L.). Theor. Appl. Genet. 2010, 120, 1021–1027. [Google Scholar] [CrossRef]
- Johnston, T.D. Club root in brassica a standard inoculation technique and the specification of races. Plant Pathol. 1968, 17, 184–187. [Google Scholar] [CrossRef]
- Guo, X.H. Biological Characteristics and Pathogenesis of Plasmodiophora brassicae. Master’s Thesis, Southwest Agriculture University, Chongqing, China, 2001. [Google Scholar]
- Si, J.; Li, C.Q.; Xiao, C.G.; Ren, X.S.; Wang, X.J. Study on the inoculation methods of Plasmodiophora brassicae. J. Southwest Univ. (Nat. Sci.) 2003, 25, 216–219. [Google Scholar]
- Chen, X.; Wang, C.; Zhang, X.X.; Wang, S. The inoculation system of cabbage clubroot and Screening of resistant germplasm resources. Plant Prot. 2015, 41, 121–126. [Google Scholar]
- Zhang, S.X.; Yang, X.Y.; Si, C.G.; Zhang, Q.X.; Wang, Y. Comparision of artificial inoculation identification methods for Chinese cabbage clubroot. Shandong Agric. Sci. 2010, 1, 78–79. [Google Scholar]
- Zhao, Y.Y.; Jiang, W.S.; Yuan, Y.X.; Zhang, X.W.; Yao, Q.J.; Zhang, Q.; Wang, Z.Y.; Wei, X.C. Comparison of indoor artificial inoculation methods and conditions of clubroot disease and identification of resistance of different cultivars of Chinese cabbage. Acta Hortic. Sin. 2014, 41, 2675. [Google Scholar]
- Suo, H.; Chen, L.Z.; Xu, H.; Song, B.; Fan, X.X.; Yuan, X.H.; He, C.Z. Research on inoculation method of of Plasmodiophora brassicae in no-heading Chinese cabbage. Jiangsu Agric. Sci. 2016, 44, 193–195. [Google Scholar]
- Zhu, H.F.; Li, X.F.; Zhu, Y.Y.; Guo, X.X.; Liu, J.P.; Gao, Q.Q.; Zhai, W. Characterization of pak-choi (Brassica chinesis) resistance to clubroot (Plasmodiophora brassica). Acta Agric. Shanghai 2016, 32, 109–113. [Google Scholar]
- Zhang, H.; Zhang, B.; Wang, C.N.; Li, M.; Huang, Z.Y.; Liu, J.F. Study on inoculation methods and condition of clubroot in qingmaye chinese cabbage. Acta Agric. Boreali Sin. 2016, 31, 182–185. [Google Scholar]
- Zhang, X.L.; Chai, A.L.; Liu, Y.M.; Fang, Z.Y.; Li, B.J.; Li, Z.S. The development of artificial inoculation system for assessment of resistance to clubroot in broccoli seedlings. J. Plant Pro. 2017, 44, 110–114. [Google Scholar]
- Huang, R.; Huang, R.R.; Hua, J.L.; Liang, Y.Y.; Shao, J.Y. Study on Inoculation Methods and Condition of Clubroot in Brassica rapa chinensis. Chinese Agri. Sci. Bulletin 2012, 28, 252–255. [Google Scholar]
- Williams, P.H. A system for the determination of races of Plasmodiophora brassicae that infect cabbage and rutabaga. Phytopathology 1966, 56, 624–626. [Google Scholar]
- Luo, H.C.; Chen, G.K.; Liu, C.P.; Huang, Y.; Xiao, C.G. An improved culture solution technique for Plasmodiophora brassicae infection and the dynamic infection in the root hair. Australas Plant Pathol. 2013, 43, 53–60. [Google Scholar] [CrossRef]
- Yuan, Y.X.; Zhao, Y.Y.; Wei, X.C.; Yao, Q.J.; Jiang, W.S.; Wang, Z.Y.; Li, Y.; Xu, Q.; Yang, S.J.; Zhang, X.W. Pathotype identification of Plasmodiophora brassicae Woron. collected from Chinese cabbage in Henan province. J. Henan Agric. Sci. 2017, 46, 71–76. [Google Scholar]
- Hoagland, D.R.; Arnon, D.I. The water-culture method for growing plants without soil. Circ. Calif. Agric. Exp. Stn. 1950, 347, 357–359. [Google Scholar]
- Strelkov, S.E.; Tewari, J.P.; Smith-Degenhardt, E. Characterization of Plasmodiophora brassicae populations from Alberta, Canada. Can. J. Plant Pathol. 2006, 28, 467–474. [Google Scholar] [CrossRef]
- Hatakeyama, K.; Niwa, T.; Kato, T.; Ohara, T.; Kakizaki, T.; Matsumoto, S. The tandem repeated organization of NB-LRR genes in the clubroot-resistant CRb locus in Brassica rapa L. Mol. Genet. Genom. 2017, 292, 397–405. [Google Scholar] [CrossRef]
- Matsumoto, E.; Ueno, H.; Aruga, D.; Sakamoto, K.; Hayashida, N. Accumulation of three clubroot resistance genes through marker-assisted selection in Chinese Cabbage (Brassica rapa ssp. pekinensis). J. Japan Soc. Hort. Sci. 2012, 81, 184–190. [Google Scholar] [CrossRef] [Green Version]
- Crute, I.R.; Gray, A.R.; Crisp, P.; Buczacki, S.T. Variation in Plasmodiophora brassicae and resistance to clubroot disease in brassicas and allied crops—A critical review. Plant Breed. Abstr. 1980, 50, 91–104. [Google Scholar]
- Kuginuki, Y.; Yoshikawa, H.; Hirai, M. Variation in virulence of Plasmodiophora brassicae in Japan tested with clubroot-resistant cultivars of Chinese cabbage (Brassica rapa L. ssp. pekinensis). Eur. J. Plant Pathol. 1999, 105, 327–332. [Google Scholar] [CrossRef]
- Manzanares-Dauleux, M.J.; Divaret, I.; Baron, F.; Thomas, G. Assessment of biological and molecular variability between and within field isolates of Plasmodiophora brassicae. Plant Pathol. 2001, 50, 165–173. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, S.; Ito, S. Pathogenic and genetic diversity in Plasmodiophora brassicae (clubroot) from Japan. J. Genl. Plant Pathol. 2013, 79, 297–306. [Google Scholar] [CrossRef]
- Strelkov, S.E.; Hwang, S.F.; Manolii, V.P.; Cao, T.; Feindel, D. Emergence of new virulence phenotypes of Plasmodiophora brassicae on canola (Brassica napus) in Alberta, Canada. Eur. J. Plant Pathol. 2016, 145, 517–529. [Google Scholar] [CrossRef]
- Gan, C.; Deng, X.; Cui, L.; Yu, X.; Yuan, W.; Dai, Z.; Yao, M.; Pang, W.; Ma, Y.; Yu, X.; et al. Construction of a high-density genetic linkage map and identification of quantitative trait loci associated with clubroot resistance in radish (Raphanus sativus L.). Mol. Breed. 2019, 39, 116. [Google Scholar] [CrossRef]
- Li, B.J. Overview of occurrence of major vegetable diseases in 2013 and occurrence trend in 2014. China Veget. 2014, 2, 5–8. [Google Scholar]
- Sun, C. Effects of Soil Factors on Clubroot Disease in Main Rapeseed Production Areas; Oil Crops Research Institute, Chinese Academy of Agricultural Sciences: Wuhan, China, 2014. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, H.; Yuan, Y.; Wei, X.; Zhang, X.; Wang, H.; Song, J.; Li, X. A New Identification Method Reveals the Resistance of an Extensive-Source Radish Collection to Plasmodiophora brassicae Race 4. Agronomy 2021, 11, 792. https://doi.org/10.3390/agronomy11040792
Yang H, Yuan Y, Wei X, Zhang X, Wang H, Song J, Li X. A New Identification Method Reveals the Resistance of an Extensive-Source Radish Collection to Plasmodiophora brassicae Race 4. Agronomy. 2021; 11(4):792. https://doi.org/10.3390/agronomy11040792
Chicago/Turabian StyleYang, Haohui, Yuxiang Yuan, Xiaochun Wei, Xiaohui Zhang, Haiping Wang, Jiangping Song, and Xixiang Li. 2021. "A New Identification Method Reveals the Resistance of an Extensive-Source Radish Collection to Plasmodiophora brassicae Race 4" Agronomy 11, no. 4: 792. https://doi.org/10.3390/agronomy11040792