
Metabonomic Profiling Analyses Reveal ANS Upregulation to Enhance the Flavonoid Pathway of Purple-Fleshed Sweet Potato Storage Root in Response to Deep Shading


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Untargeted Metabolomics Analysis of Storage Roots
2.3. Measurement of Soluble Sugar, Starch, and Anthocyanin Contents in Sweet Potato
2.4. Measurement of Amino Acid Contents in Developing Storage Roots
2.5. Measurement of CHS, ANS, and UFGT Enzyme Activities
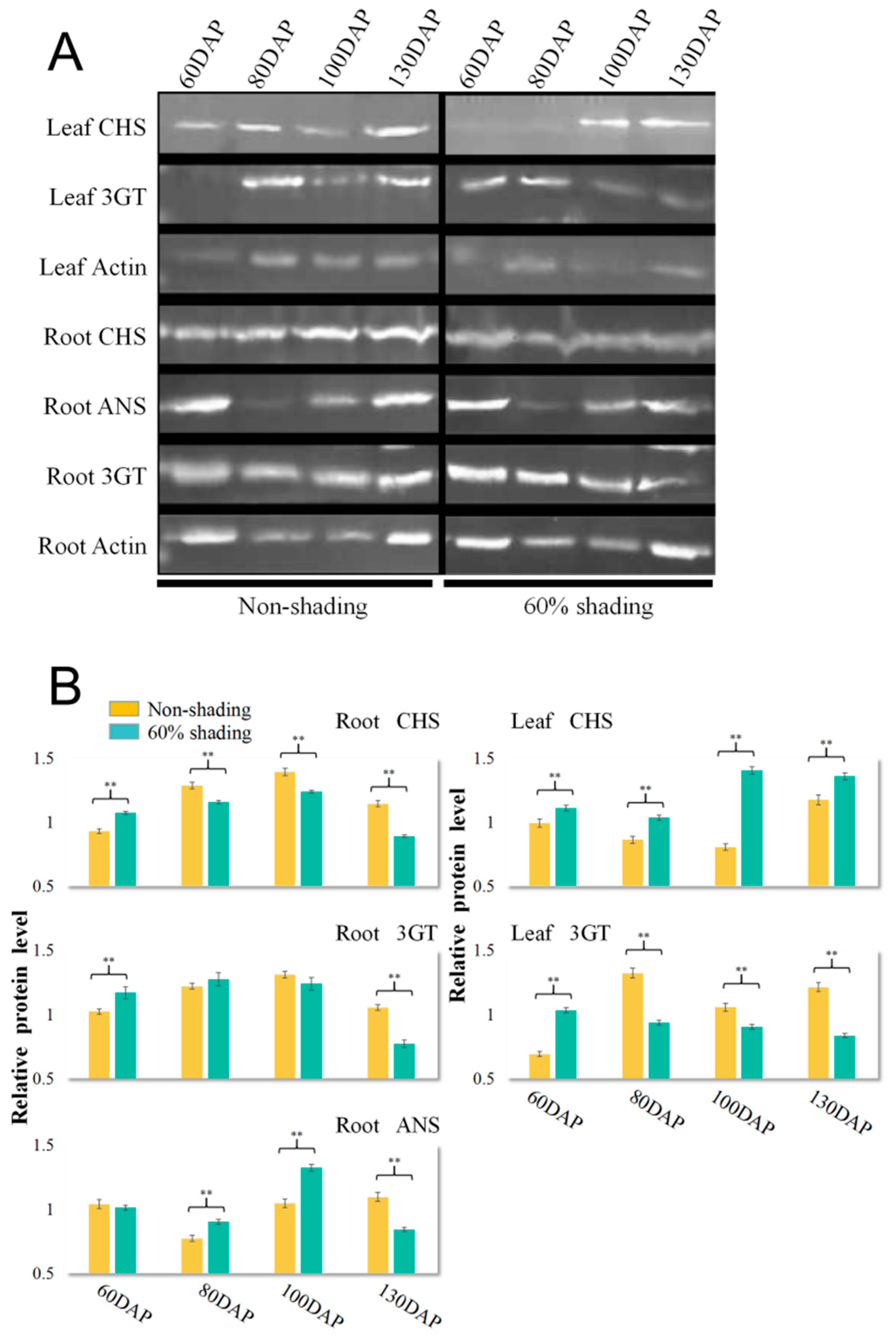
2.6. Measurement of CHS, ANS and 3GT Protein Expression by Western Blot Assay
2.7. qPCR Verification of CHS, ANS, and UFGT Gene Expression
2.8. Data Analysis
3. Results
3.1. Effects of Shading on Storage Root Yield and Anthocyanin Content in PSP
3.2. Tissue-Specific and Cultivar-Specific Effects on Soluble Sugar Contents and Starch Accumulation by Shading Treatment
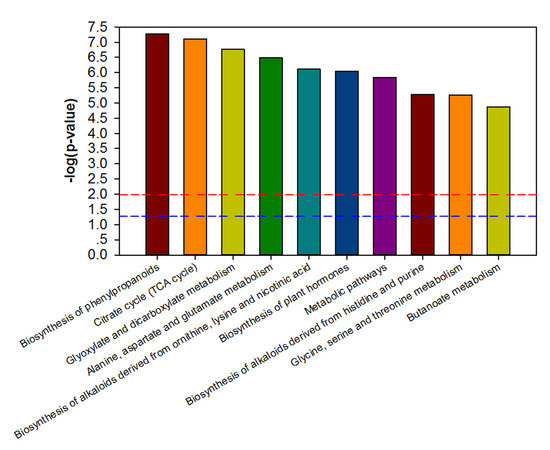
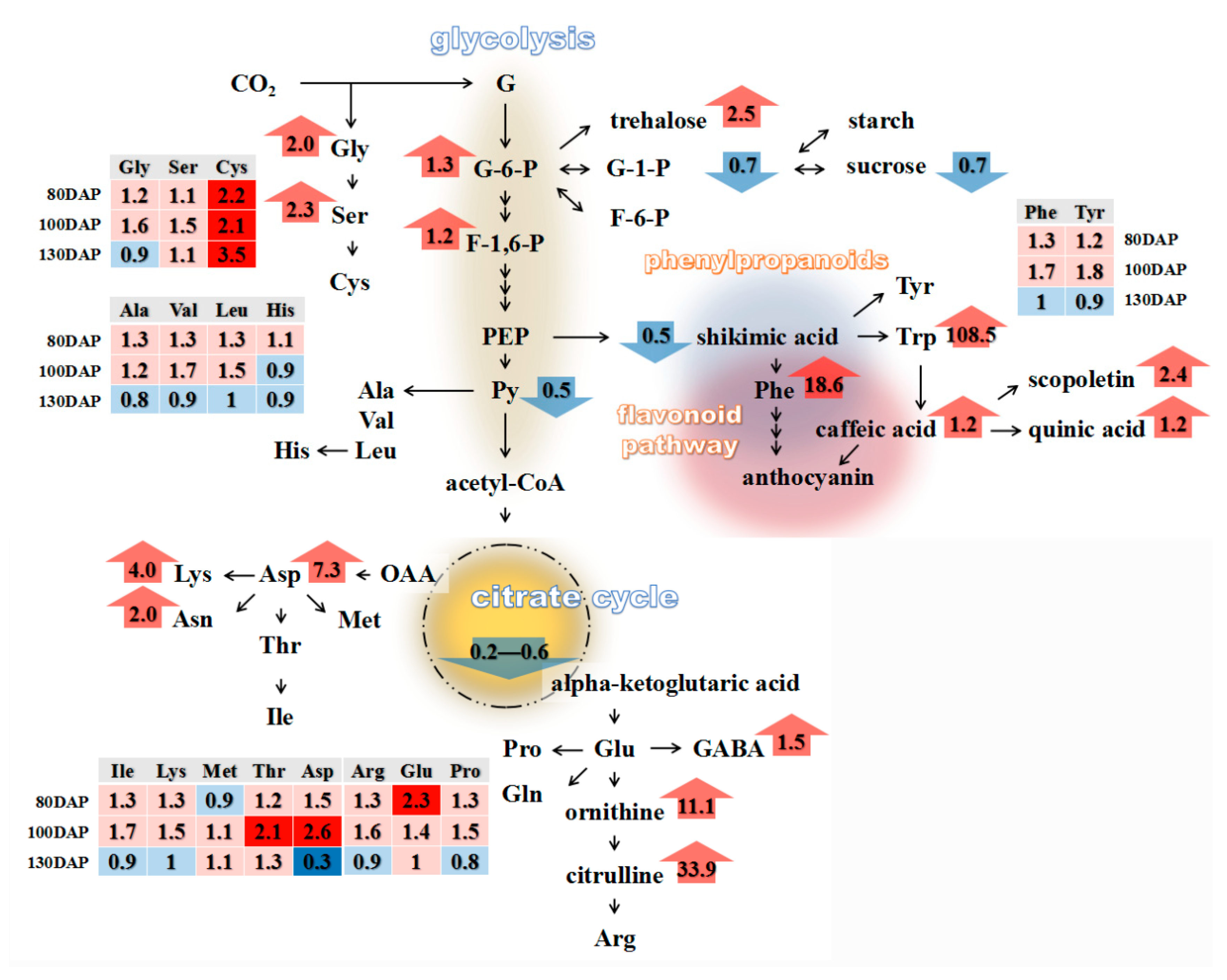
3.3. Metabolic Characterization of Ripe PSP Storage Roots under Shading
3.4. Changes in the Amino Acid Contents of PSP Storage Roots in Response to Shading
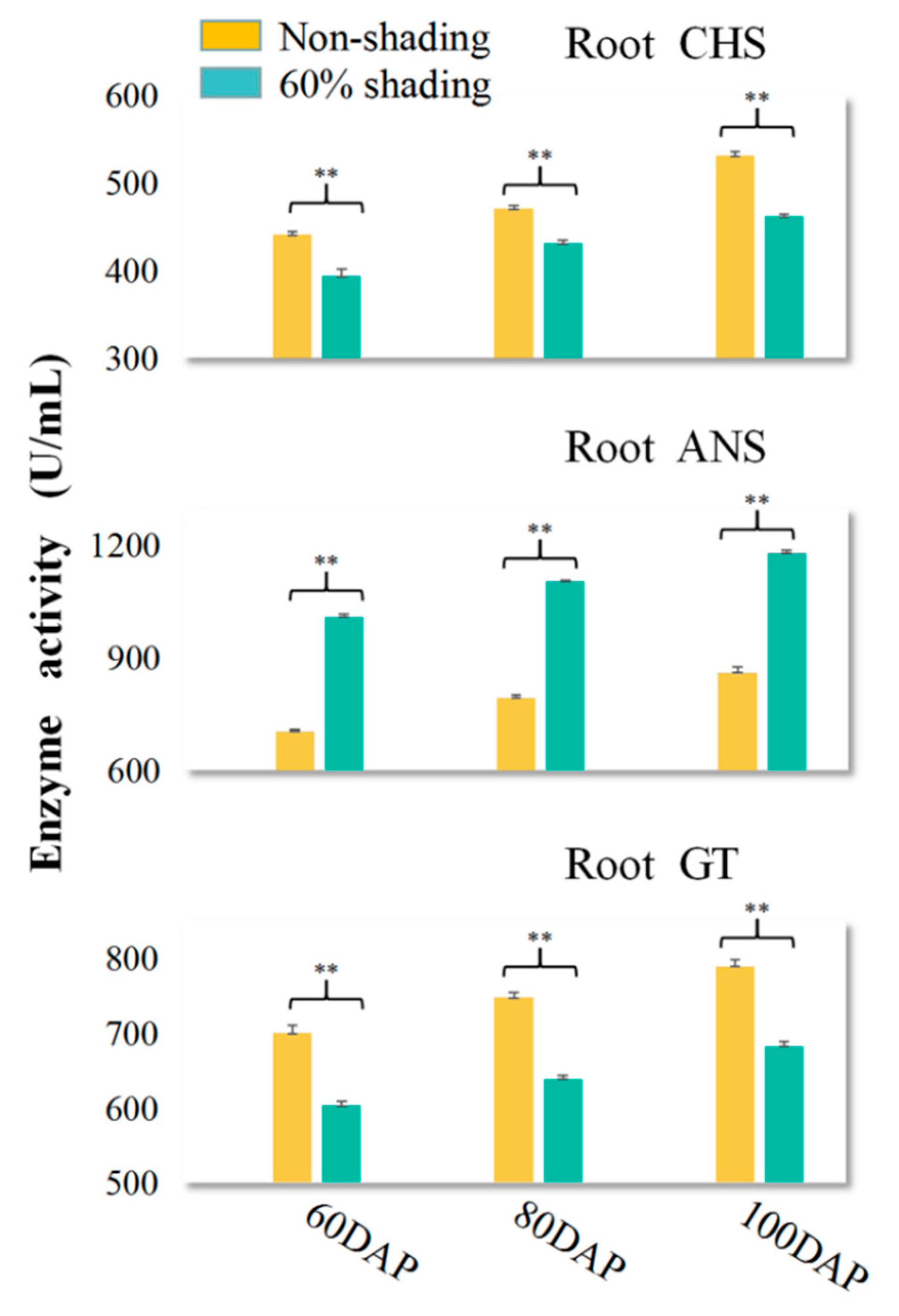
3.5. Effects of Shading on Three Key Anthocyanin Metabolism Enzymes, CHS, ANS, and 3GT, in Treated PSP Storage Roots
4. Discussion
4.1. Effect of Shading Treatment on Storage Root Development in Sweet Potato and Species-Specific Strategy
4.2. Effect on Key Enzyme Expression of Flavonoid Pathways to Promote PSP Storage Root Anthocyanin Biosynthesis in Response to Shading Treatment
4.3. Nitrogen/Carbon Balance and Sugar/Hormone Signalling to Regulate Storage Root Flavonoid Pathways under Shading
5. Conclusions
6. Patents
Author Contributions
Funding
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Accession | Primer Sequence (5′-3′) |
|---|---|---|
| ANS | FJ478179 | GCTGACACGCCCAAACCT CCATAATAATACCAAACGGACT |
| CHS | AB037680 | GCACTTGGATAGCCTGGTC GAGTAAATGGAACGTGAGCC |
| 3GT | JN258961 | GACGGATTCGTTTCTGTGG GTTTGCGTTTGGAGTGGC |
| ACTIN | EU250003 | GAGACCTTCAACACCCCTGCTA ATCACCAGAGTCCAACACATTACCT |




References
- Jaakola, L.; Määttä-Riihinen, K.; Kärenlampi, S.; Hohtola, A. Activation of flavonoid biosynthesis by solar radiation in bilberry (Vaccinium myrtillus L.) leaves. Planta 2004, 218, 721–728. [Google Scholar]
- Deluc, L.; Bogs, J.; Walker, A.R.; Ferrier, T.; Decendit, A.; Merillon, J.-M.; Robinson, S.P.; Barrieu, F. The transcription factor VvMYB5b contributes to the regulation of anthocyanin and proanthocyanidin biosynthesis in developing grape berries. Plant Physiol. 2008, 147, 2041–2053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, S.; Ishimaru, M.; Hiraoka, K.; Honda, C. Myb-related genes of the Kyoho grape (Vitis labruscana) regulate anthocyanin biosynthesis. Planta 2002, 215, 924–933. [Google Scholar]
- Kobayashi, S.; Goto-Yamamoto, N.; Hirochika, H. Retrotransposon-induced mutations in grape skin color. Science 2004, 304, 982. [Google Scholar] [CrossRef] [PubMed]
- Ogo, Y.; Mori, T.; Nakabayashi, R.; Saito, K.; Takaiwa, F. Transgenic rice seed expressing flavonoid biosynthetic genes accumulate glycosylated and/or acylated flavonoids in protein bodies. J. Exp. Bot. 2016, 67, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Wang, F.; Lian, Y.; Xiao, H.; Zheng, J. Biosynthesis of citrus flavonoids and their health effects. Crit. Rev. Food Sci. Nutr. 2020, 60, 566–583. [Google Scholar] [CrossRef] [PubMed]
- Tohge, T.; de Souza, L.P.; Fernie, A.R. Current understanding of the pathways of flavonoid biosynthesis in model and crop plants. J. Exp. Bot. 2017, 68, 4013–4028. [Google Scholar] [CrossRef]
- Lepiniec, L.; Debeaujon, I.; Routaboul, J.-M.; Baudry, A.; Pourcel, L.; Nesi, N.; Caboche, M. Genetics and biochemistry of seed flavonoids. Annu. Rev. Plant Biol. 2006, 57, 405–430. [Google Scholar] [CrossRef]
- Fuglevand, G.; Jackson, J.A.; Jenkins, G.I. UV-B, UV-A, and blue light signal transduction pathways interact synergistically to regulate chalcone synthase gene expression in Arabidopsis. Plant Cell 1996, 8, 2347–2357. [Google Scholar]
- Lloyd, A.; Brockman, A.; Aguirre, L.; Campbell, A.; Bean, A.; Cantero, A.; Gonzalez, A. Advances in the MYB–bHLH–WD repeat (MBW) pigment regulatory model: Addition of a WRKY factor and co-option of an anthocyanin MYB for betalain regulation. Plant Cell Physiol. 2017, 58, 1431–1441. [Google Scholar] [CrossRef] [Green Version]
- Henry-Kirk, R.A.; Plunkett, B.; Hall, M.; McGhie, T.; Allan, A.C.; Wargent, J.J.; Espley, R.V. Solar UV light regulates flavonoid metabolism in apple (Malus × domestica). Plant Cell Environ. 2018, 41, 675–688. [Google Scholar] [CrossRef]
- Cohen, S.D.; Tarara, J.M.; Gambetta, G.A.; Matthews, M.A.; Kennedy, J.A. Impact of diurnal temperature variation on grape berry development, proanthocyanidin accumulation, and the expression of flavonoid pathway genes. J. Exp. Bot. 2012, 63, 2655–2665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koyama, K.; Ikeda, H.; Poudel, P.R.; Goto-Yamamoto, N. Light quality affects flavonoid biosynthesis in young berries of Cabernet Sauvignon grape. Phytochemistry 2012, 78, 54–64. [Google Scholar] [CrossRef] [PubMed]
- Goh, H.H.; Khairudin, K.; Sukiran, N.A.; Normah, M.; Baharum, S. Metabolite profiling reveals temperature effects on the VOC s and flavonoids of different plant populations. Plant Biol. 2016, 18, 130–139. [Google Scholar] [CrossRef]
- Jia, H.; Jiu, S.; Zhang, C.; Wang, C.; Tariq, P.; Liu, Z.; Wang, B.; Cui, L.; Fang, J. Abscisic acid and sucrose regulate tomato and strawberry fruit ripening through the abscisic acid-stress-ripening transcription factor. Plant Biotechnol. J. 2016, 14, 2045–2065. [Google Scholar] [CrossRef] [PubMed]
- Gambetta, G.A.; Matthews, M.A.; Shaghasi, T.H.; McElrone, A.J.; Castellarin, S.D. Sugar and abscisic acid signaling orthologs are activated at the onset of ripening in grape. Planta 2010, 232, 219–234. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Tian, L.; Liu, H.; Pan, Q.; Zhan, J.; Huang, W. Sugars induce anthocyanin accumulation and flavanone 3-hydroxylase expression in grape berries. Plant Growth Regul. 2009, 58, 251–260. [Google Scholar] [CrossRef]
- Jia, H.; Xie, Z.; Wang, C.; Shangguan, L.; Qian, N.; Cui, M.; Liu, Z.; Zheng, T.; Wang, M.; Fang, J. Abscisic acid, sucrose, and auxin coordinately regulate berry ripening process of the Fujiminori grape. Funct. Integr. Genomics 2017, 17, 441–457. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Li, H.; Ma, X.; Dong, H.; Ma, D.; Zeng, H. Proteome alterations of reverse photoperiod-sensitive genic male sterile rice (Oryza sativa L.) at fertility transformation stage. Genes Genomics 2014, 36, 711–726. [Google Scholar] [CrossRef]
- Acero, N.; Gradillas, A.; Beltran, M.; García, A.; Mingarro, D.M. Comparison of phenolic compounds profile and antioxidant properties of different sweet cherry (Prunus avium L.) varieties. Food Chem. 2019, 279, 260–271. [Google Scholar] [CrossRef]
- Bai, S.; Tao, R.; Tang, Y.; Yin, L.; Ma, Y.; Ni, J.; Yan, X.; Yang, Q.; Wu, Z.; Zeng, Y. BBX16, a B-box protein, positively regulates light-induced anthocyanin accumulation by activating MYB10 in red pear. Plant Biotechnol. J. 2019, 17, 1985–1997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mei, X.; Zhou, C.; Zhang, W.; Rothenberg, D.O.N.; Wan, S.; Zhang, L. Comprehensive analysis of putative dihydroflavonol 4-reductase gene family in tea plant. PLoS ONE 2019, 14, e0227225. [Google Scholar] [CrossRef] [PubMed]
- Barbieri, P.; Pellerin, S.; Nesme, T. Comparing crop rotations between organic and conventional farming. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Hoffland, E.; Kuyper, T.W.; Yu, Y.; Zhang, C.; Li, H.; Zhang, F.; van der Werf, W. Syndromes of production in intercropping impact yield gains. Nat. Plants 2020, 6, 653–660. [Google Scholar] [CrossRef] [PubMed]
- Voisin, A.-S.; Guéguen, J.; Huyghe, C.; Jeuffroy, M.-H.; Magrini, M.-B.; Meynard, J.-M.; Mougel, C.; Pellerin, S.; Pelzer, E. Legumes for feed, food, biomaterials and bioenergy in Europe: A review. Agron. Sustain. Dev. 2014, 34, 361–380. [Google Scholar] [CrossRef]
- Midmore, D.J. Scientific basis and scope for further improvement of intercropping with potato in the tropics. Field Crop. Res. 1990, 25, 3–24. [Google Scholar] [CrossRef]
- Mushagalusa, G.N.; Ledent, J.-F.; Draye, X. Shoot and root competition in potato/maize intercropping: Effects on growth and yield. Environ. Exp. Bot. 2008, 64, 180–188. [Google Scholar] [CrossRef]
- Brooker, R.W.; Bennett, A.E.; Cong, W.F.; Daniell, T.J.; George, T.S.; Hallett, P.D.; Hawes, C.; Iannetta, P.P.; Jones, H.G.; Karley, A.J. Improving intercropping: A synthesis of research in agronomy, plant physiology and ecology. New Phytol. 2015, 206, 107–117. [Google Scholar] [CrossRef]
- Krieger-Liszkay, A.; Krupinska, K.; Shimakawa, G. The impact of photosynthesis on initiation of leaf senescence. Physiol. Plant. 2019, 166, 148–164. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Zhang, Y.; Dai, J.; Dong, H.; Kong, X. High plant density inhibits vegetative branching in cotton by altering hormone contents and photosynthetic production. Field Crop. Res. 2019, 230, 121–131. [Google Scholar] [CrossRef]
- Wang, Q.; Hou, F.; Dong, S.; Xie, B.; Li, A.; Zhang, H.; Zhang, L. Effects of shading on the photosynthetic capacity, endogenous hormones and root yield in purple-fleshed sweetpotato (Ipomoea batatas (L.) Lam). Plant Growth Regul. 2014, 72, 113–122. [Google Scholar] [CrossRef]
- Lai, J.-l.; Liu, Z.-w.; Luo, X.-g. A metabolomic, transcriptomic profiling, and mineral nutrient metabolism study of the phytotoxicity mechanism of uranium. J. Hazard. Mater. 2020, 386, 121437. [Google Scholar] [CrossRef] [PubMed]
- Scandiani, M.M.; Luque, A.G.; Razori, M.V.; Ciancio Casalini, L.; Aoki, T.; O’Donnell, K.; Cervigni, G.D.; Spampinato, C.P. Metabolic profiles of soybean roots during early stages of Fusarium tucumaniae infection. J. Exp. Bot. 2015, 66, 391–402. [Google Scholar] [CrossRef] [Green Version]
- Laurentin, A.; Edwards, C.A. A microtiter modification of the anthrone-sulfuric acid colorimetric assay for glucose-based carbohydrates. Anal. Biochem. 2003, 315, 143–145. [Google Scholar] [CrossRef]
- Jing, P.; Zhao, S.-J.; Ruan, S.-Y.; Xie, Z.-H.; Dong, Y.; Yu, L.L. Anthocyanin and glucosinolate occurrences in the roots of Chinese red radish (Raphanus sativus L.), and their stability to heat and pH. Food Chem. 2012, 133, 1569–1576. [Google Scholar] [CrossRef]
- Aredes, R.S.; Peixoto, F.C.; Sphaier, L.A.; de Carvalho Marques, F.F. Evaluation of craft beers through the direct determination of amino acids by capillary electrophoresis and principal component analysis. Food Chem. 2021, 344, 128572. [Google Scholar] [CrossRef] [PubMed]
- Qiao, X.; He, Y.; Wang, Z.; Li, X.; Zhang, K.; Zeng, H. Effect of foliar spray of zinc on chloroplast β-carbonic anhydrase expression and enzyme activity in rice (Oryza sativa L.) leaves. Acta Physiol. Plant 2014, 36, 263–272. [Google Scholar] [CrossRef]
- Zhu, L.; Chen, Z.; Li, H.; Sun, Y.; Wang, L.; Zeng, H.; He, Y. Lipid metabolism is involved in male fertility regulation of the photoperiod-and thermo sensitive genic male sterile rice line Peiai 64S. Plant Sci. 2020, 299, 110581. [Google Scholar] [CrossRef]
- He, W.; Zeng, M.; Chen, J.; Jiao, Y.; Niu, F.; Tao, G.; Zhang, S.; Qin, F.; He, Z. Identification and quantitation of anthocyanins in purple-fleshed sweet potatoes cultivated in China by UPLC-PDA and UPLC-QTOF-MS/MS. J. Agric. Food Chem. 2016, 64, 171–177. [Google Scholar] [CrossRef]
- Li, J.; Li, X.-d.; Zhang, Y.; Zheng, Z.-d.; Qu, Z.-y.; Liu, M.; Zhu, S.-h.; Liu, S.; Wang, M.; Qu, L. Identification and thermal stability of purple-fleshed sweet potato anthocyanins in aqueous solutions with various pH values and fruit juices. Food Chem. 2013, 136, 1429–1434. [Google Scholar] [CrossRef]
- Tanner, G.J.; Francki, K.T.; Abrahams, S.; Watson, J.M.; Larkin, P.J.; Ashton, A.R. Proanthocyanidin biosynthesis in plants: Purification of legume leucoanthocyanidin reductase and molecular cloning of its cDNA. J. Biol. Chem. 2003, 278, 31647–31656. [Google Scholar] [CrossRef] [Green Version]
- Gommers, C.M.; Visser, E.J.; St Onge, K.R.; Voesenek, L.A.; Pierik, R. Shade tolerance: When growing tall is not an option. Trends Plant Sci. 2013, 18, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Gong, W.; Jiang, C.; Wu, Y.; Chen, H.; Liu, W.; Yang, W. Tolerance vs. avoidance: Two strategies of soybean (Glycine max) seedlings in response to shade in intercropping. Photosynthetica 2015, 53, 259–268. [Google Scholar] [CrossRef]
- Yang, F.; Liao, D.; Wu, X.; Gao, R.; Fan, Y.; Raza, M.A.; Wang, X.; Yong, T.; Liu, W.; Liu, J. Effect of aboveground and belowground interactions on the intercrop yields in maize-soybean relay intercropping systems. Field Crop. Res. 2017, 203, 16–23. [Google Scholar] [CrossRef]
- Shafiq, I.; Hussain, S.; Raza, M.A.; Iqbal, N.; Asghar, M.A.; Ali, R.; Fan, Y.-f.; Mumtaz, M.; Shoaib, M.; Ansar, M. Crop photosynthetic response to light quality and light intensity. J. Integr. Agric. 2021, 20, 4–23. [Google Scholar] [CrossRef]
- Wu, Y.; Gong, W.; Yang, W. Shade inhibits leaf size by controlling cell proliferation and enlargement in soybean. Sci. Rep. 2017, 7, 9295. [Google Scholar] [CrossRef]
- Song, D.; Tariq, A.; Pan, K.; Khan, S.U.; Saleh, T.A.; Gong, S.; Zhang, A.; Wu, X. Influence of planting distance and density on the yield and photosynthetic traits of sweet potato (Ipomoea balatas L.) under an intercropping system with walnut (Juglans regia) saplings. Soil Tillage Res. 2020, 196, 104484. [Google Scholar] [CrossRef]
- Fogelman, E.; Tanami, S.; Ginzberg, I. Anthocyanin synthesis in native and wound periderms of potato. Physiol. Plant. 2015, 153, 616–626. [Google Scholar] [CrossRef] [PubMed]
- Loreti, E.; Povero, G.; Novi, G.; Solfanelli, C.; Alpi, A.; Perata, P. Gibberellins, jasmonate and abscisic acid modulate the sucrose-induced expression of anthocyanin biosynthetic genes in Arabidopsis. New Phytol. 2008, 179, 1004–1016. [Google Scholar] [CrossRef]
- Wingler, A. Transitioning to the next phase: The role of sugar signaling throughout the plant life cycle. Plant Physiol. 2018, 176, 1075–1084. [Google Scholar] [CrossRef] [Green Version]
- Cominelli, E.; Gusmaroli, G.; Allegra, D.; Galbiati, M.; Wade, H.K.; Jenkins, G.I.; Tonelli, C. Expression analysis of anthocyanin regulatory genes in response to different light qualities in Arabidopsis thaliana. J. Plant Physiol. 2008, 165, 886–894. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Tang, X.; Huang, H.; Zhang, Y.; Dai, S. Transcriptomic analyses reveal species-specific light-induced anthocyanin biosynthesis in chrysanthemum. BMC Genom. 2015, 16, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, F.; Liu, J.; Ma, Y.; Chen, J.; Keyoumu· Abudurezikekey, A.; Wang, Y.; Chen, B.; Meng, Y.; Zhou, Z. Effect of shading on cotton yield and quality on different fruiting branches. Crop Sci. 2013, 53, 2670–2678. [Google Scholar] [CrossRef]
- Solfanelli, C.; Poggi, A.; Loreti, E.; Alpi, A.; Perata, P. Sucrose-specific induction of the anthocyanin biosynthetic pathway in Arabidopsis. Plant Physiol. 2006, 140, 637–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.; Iqbal, N.; Hu, B.; Zhang, Q.; Wu, H.; Liu, X.; Zhang, J.; Liu, W.; Yang, W.; Liu, J. Targeted metabonomics analysis of fatty acid in soybean seed by GC-MS reveal the metabolic manipulation of shading in intercropping system. Anal. Methods 2017, 3, 1–27. [Google Scholar]
- Feng, L.; Raza, M.A.; Li, Z.; Chen, Y.; Khalid, M.H.B.; Du, J.; Liu, W.; Wu, X.; Song, C.; Yu, L.; et al. The influence of light intensity and leaf movement on photosynthesis characteristics and carbon balance of soybean. Front. Plant Sci. 2019, 9, 1952. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Zou, R.; Pu, D.; Lan, Z.; Zhao, B. Non-targeted and targeted metabolomics profiling of tea plants (Camellia sinensis) in response to its intercropping with Chinese chestnut. BMC Plant Biol. 2021, 21, 55. [Google Scholar] [CrossRef]
- Wu, Y.; Shi, L.; Li, L.; Fu, L.; Liu, Y.; Xiong, Y.; Sheen, J. Integration of nutrient, energy, light, and hormone signalling via TOR in plants. J. Exp. Bot. 2019, 70, 2227–2238. [Google Scholar] [CrossRef]
- Liu, L.; Li, Y.; She, G.; Zhang, X.; Jordan, B.; Chen, Q.; Zhao, J.; Wan, X. Metabolite profiling and transcriptomic analyses reveal an essential role of UVR8-mediated signal transduction pathway in regulating flavonoid biosynthesis in tea plants (Camellia sinensis) in response to shading. BMC Plant Biol. 2018, 18, 233. [Google Scholar] [CrossRef]
- Lloyd, J.C.; Zakhleniuk, O.V. Responses of primary and secondary metabolism to sugar accumulation revealed by microarray expression analysis of the Arabidopsis mutant, pho3. J. Exp. Bot. 2004, 55, 1221–1230. [Google Scholar] [CrossRef]
- Fernie, A.R.; Bachem, C.W.; Helariutta, Y.; Neuhaus, H.E.; Prat, S.; Ruan, Y.-L.; Stitt, M.; Sweetlove, L.J.; Tegeder, M.; Wahl, V. Synchronization of developmental, molecular and metabolic aspects of source–sink interactions. Nat. Plants 2020, 6, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Wingler, A.; Delatte, T.L.; O’Hara, L.E.; Primavesi, L.F.; Jhurreea, D.; Paul, M.J.; Schluepmann, H. Trehalose 6-phosphate is required for the onset of leaf senescence associated with high carbon availability. Plant Physiol. 2012, 158, 1241–1251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, T.; Obata, T.; Feil, R.; Lunn, J.E.; Fujita, Y.; Yamaguchi-Shinozaki, K.; Fernie, A.R. The role of abscisic acid signaling in maintaining the metabolic balance required for Arabidopsis growth under nonstress conditions. Plant Cell 2019, 31, 84–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]









| Group | Storage Root FW (g) | Storage Root Num. | Outputs Per Plant (kg) | Anthocyanin Content Per Plant (mg) | Storage Root Yield (t/hm2) | Anthocyanin Outputs (g/hm2) |
|---|---|---|---|---|---|---|
| Non shading | 82.63 ± 3.07 | 7.0 | 0.58 ± 0.06 | 73.95 ± 7.65 | 47.22 | 6020.19 |
| 30% shading | 82.14 ± 6.57 | 5.2 | 0.43 ± 0.03 ** | 93.01 ± 6.49 ** | 34.87 | 7541.86 |
| 60% shading | 75.05 ± 9.53 * | 4.6 | 0.35 ± 0.10 ** | 91.70 ± 26.21 * | 28.18 | 7383.69 |
| Differential Metabolites | VIP 2 | FC 3 | Differential Metabolites | VIP 2 | FC 3 | Differential Metabolites | VIP 2 | FC 3 |
|---|---|---|---|---|---|---|---|---|
| Carbohydrates and carbohydrate conjugates 1 | Carbohydrates and carbohydrate conjugates 1 | Amino acids, peptides and analogues 1 | ||||||
| L-Threose | 1.37 | 0.25 | P-benzoquinone | 1.14 | 0.77 | β-Alanine | 1.21 | 0.20 |
| Glycerol | 1.51 | 1.00 | Allo-inositol | / | 0.47 | Valine | 1.60 | 1.73 |
| D-Glyceric acid | 1.31 | 0.86 | Myo-inositol | / | 0.57 | D-alanyl-D-alanine | 1.28 | 0.76 |
| Lyxose | 1.54 | 0.13 | Quinic acid | / | 1.16 | Norleucine | 1.22 | 1.35 |
| Dihydroxyacetone | 1.49 | 0.43 | Shikimic acid | / | 0.47 | L-Isoleucine | 1.47 | 0.92 |
| Erythrose | 1.66 | 0.07 | Chlorogenic acid | / | 2.29 | Glycine | 1.21 | 2.07 |
| Threitol | / | 0.18 | Serine | 1.49 | 2.32 | |||
| Threonic acid | / | 0.12 | Organic acids and derivatives 1 | 3-Cyanoalanine | 1.38 | 1.16 | ||
| Glucose-1-phosphate | / | 0.70 | Urea | 1.63 | 1.15 | Threonine | 1.41 | 1.17 |
| Tartaric acid | / | 0.00 | Aconitic acid | / | 0.20 | Gly-Pro | 1.29 | 2.57 |
| Sorbitol | / | 0.07 | Citric acid | / | 0.35 | L-dopa | 1.15 | 1.69 |
| Ribose | / | 0.68 | Isocitric acid | / | 0.28 | Glycocyamine | 1.11 | 0.43 |
| Xylose | / | 0.43 | 2-Ketobutyric acid | 1.04 | 0.97 | L-Allothreonine | 1.36 | 0.25 |
| Galactinol | / | 0.82 | 2-Ketovaleric acid | 1.01 | 1.06 | N-Acetyl-L-phenylalanine | 1.11 | 0.33 |
| Xylitol | / | 0.67 | α-Ketoisocaproic acid | 1.41 | 0.00 | Methionine | 1.56 | 1.33 |
| Sedoheptulose | / | 0.66 | Methyl phosphate | 1.56 | 0.81 | Oxoproline | 1.25 | 1.20 |
| D-Arabitol | / | 0.45 | O-Phosphorylethanolamine | / | 3.51 | Aspartic acid | 1.29 | 7.30 |
| Fucose | / | 0.81 | Galactonic acid | / | 0.29 | 3-hydroxy-L-proline | / | 0.52 |
| Gluconic acid | / | 0.30 | α-Ketoglutaric acid | / | 0.56 | 4-aminobutyric acid | / | 1.51 |
| Mucic acid | / | 0.26 | Oxalic acid | 1.59 | 2.44 | L-glutamic acid | / | 1.02 |
| Saccharic acid | / | 0.37 | Succinic acid | 1.25 | 0.23 | Ornithine | / | 11.10 |
| 1-Kestose | / | 0.44 | Fumaric acid | 1.33 | 0.23 | Glutamic acid | / | 1.86 |
| 1,5-Anhydroglucitol | / | 1.20 | Malonic acid | / | 15.82 | Phenylalanine | / | 18.64 |
| Tagatose | / | 0.32 | Methylmalonic acid | / | 0.00 | Sarcosine | / | 3.41 |
| Glucuronic acid | / | 0.36 | β-Hydroxypyruvate | 1.23 | 0.99 | N-Carbamylglutamate | / | 1.05 |
| Mannitol | / | 1.01 | 3-Hydroxypropionic acid | 1.24 | 0.40 | Asparagine | / | 1.97 |
| Glucose-6-phosphate | / | 1.30 | L-Malic acid | 1.58 | 0.26 | α-Aminoadipic acid | / | 2.86 |
| Digalacturonic acid | / | 1.89 | Pyruvic acid | 1.51 | 0.47 | Tyrosine | / | 0.94 |
| D-Fructose 1,6-bisphosphate | / | 1.16 | Lactic acid | 1.61 | 0.12 | Glutamine | / | 0.80 |
| Maltotriose | / | 0.58 | Glycolic acid | 1.43 | 0.69 | Citrulline | / | 33.91 |
| Arbutin | / | 177,050.00 | Allantoic acid | / | 0.81 | |||
| Sucrose | / | 0.74 | Amino acids, peptides, and analogues 1 | N-Methyl-L-glutamic acid | / | 12.62 | ||
| Isomaltose | / | 0.46 | 3-Aminoisobutyric acid | 1.38 | 0.30 | Lysine | / | 3.98 |
| Maltose | / | 0.77 | Nicotinoylglycine | 1.38 | 0.64 | Iminodiacetic acid | / | 3.45 |
| Ribitol | / | 0.15 | Alanine | 1.00 | 0.83 | Kyotorphin | / | 0.86 |
| Melezitose | / | 5.40 | N-Ethylglycine | 1.57 | 0.67 | |||
| Trehalose | / | 2.47 | Proline | 1.37 | 1.19 | |||
| Gluconic lactone | / | 4.80 | Pipecolinic acid | 1.42 | 0.58 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, Y.; Zhu, D.; Sun, Y.; Wang, Q.; Zhu, L.; Zeng, H. Metabonomic Profiling Analyses Reveal ANS Upregulation to Enhance the Flavonoid Pathway of Purple-Fleshed Sweet Potato Storage Root in Response to Deep Shading. Agronomy 2021, 11, 737. https://doi.org/10.3390/agronomy11040737
He Y, Zhu D, Sun Y, Wang Q, Zhu L, Zeng H. Metabonomic Profiling Analyses Reveal ANS Upregulation to Enhance the Flavonoid Pathway of Purple-Fleshed Sweet Potato Storage Root in Response to Deep Shading. Agronomy. 2021; 11(4):737. https://doi.org/10.3390/agronomy11040737
Chicago/Turabian StyleHe, Ying, Dan Zhu, Yujun Sun, Qian Wang, Lan Zhu, and Hanlai Zeng. 2021. "Metabonomic Profiling Analyses Reveal ANS Upregulation to Enhance the Flavonoid Pathway of Purple-Fleshed Sweet Potato Storage Root in Response to Deep Shading" Agronomy 11, no. 4: 737. https://doi.org/10.3390/agronomy11040737