Sustainable Transfer of Tomato Landraces to Modern Cropping Systems: The Effects of Environmental Conditions and Management Practices on Long-Shelf-Life Tomatoes


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
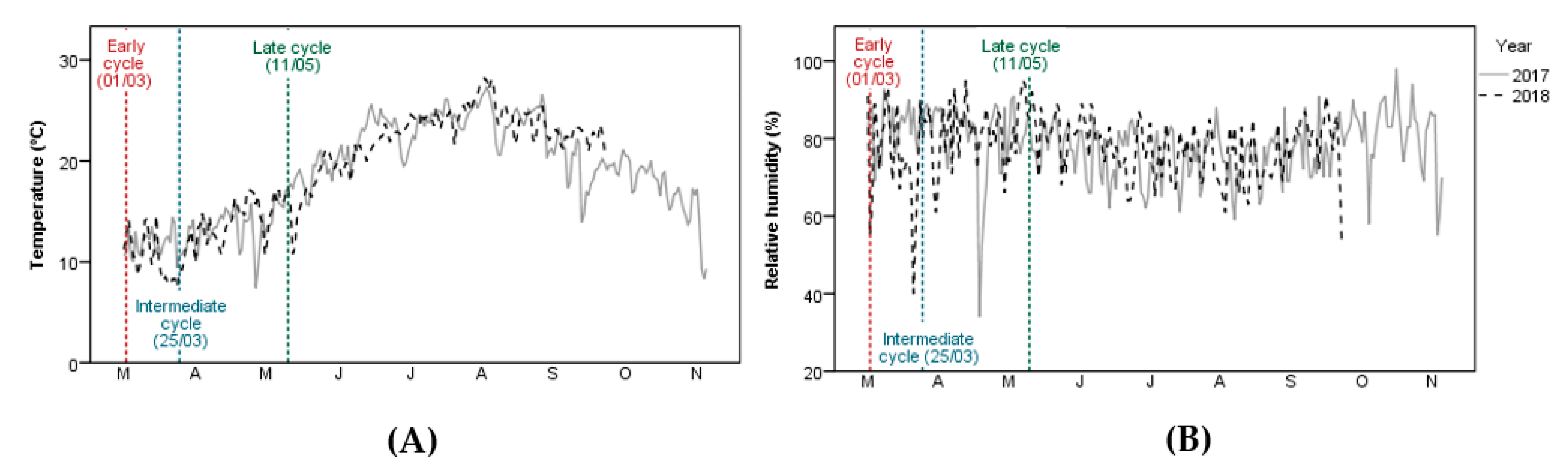
2.2. Experimental Conditions and Crop Management Practices
2.3. Measurements
2.4. Data Analysis
3. Results
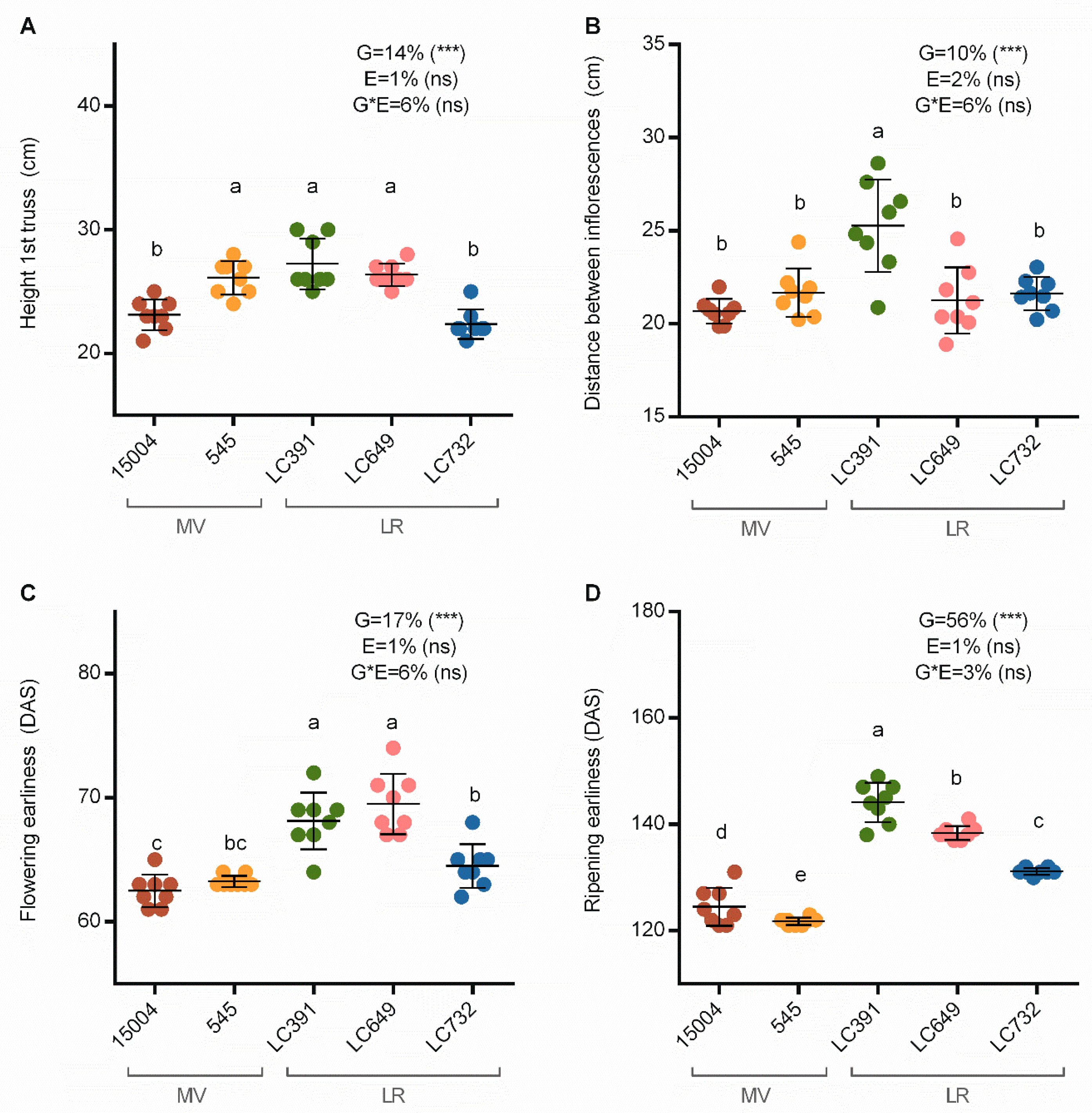
3.1. Earliness and Plant Architecture
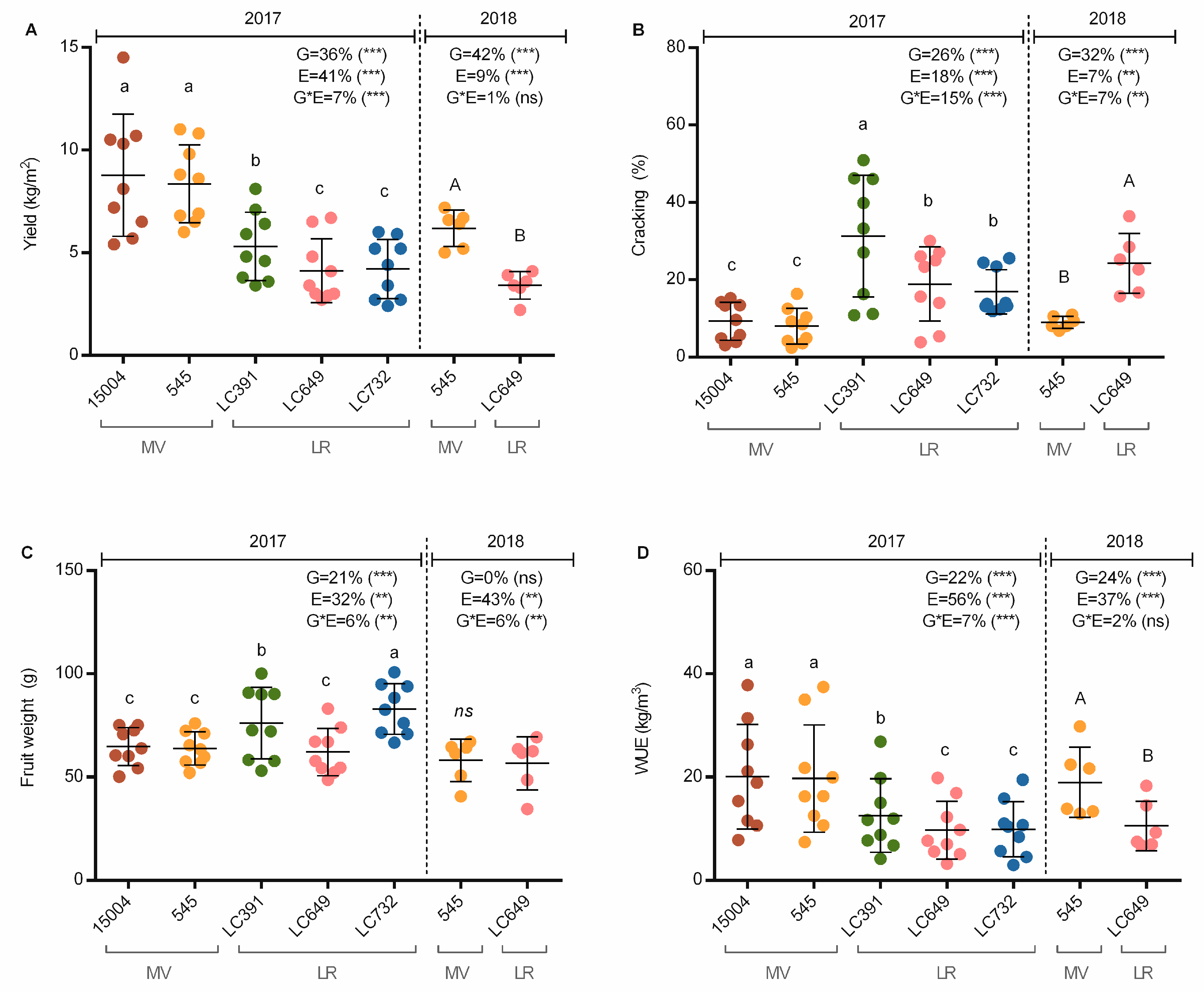
3.2. Agronomic Performance
3.3. Fruit Quality
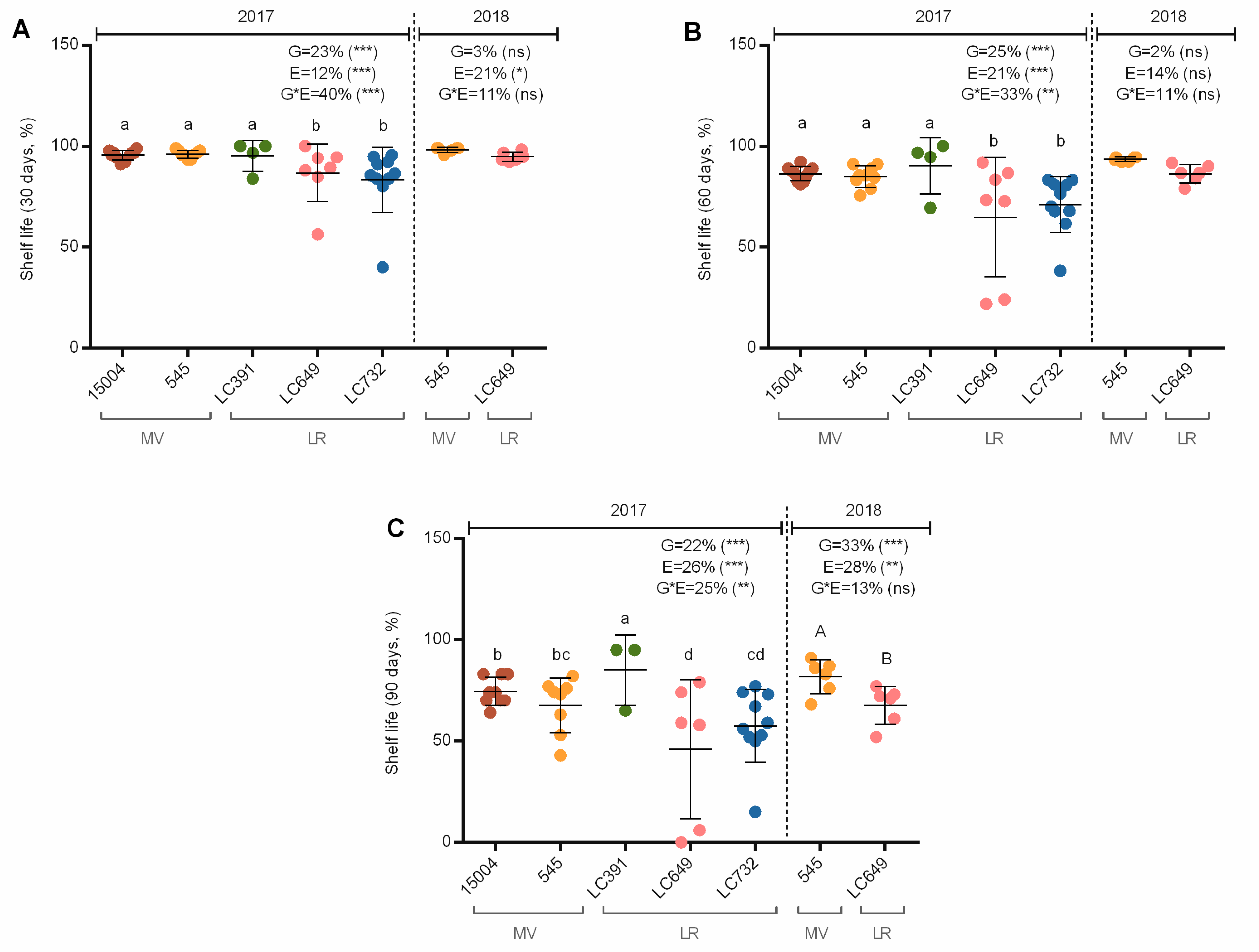
3.4. Shelf-Life
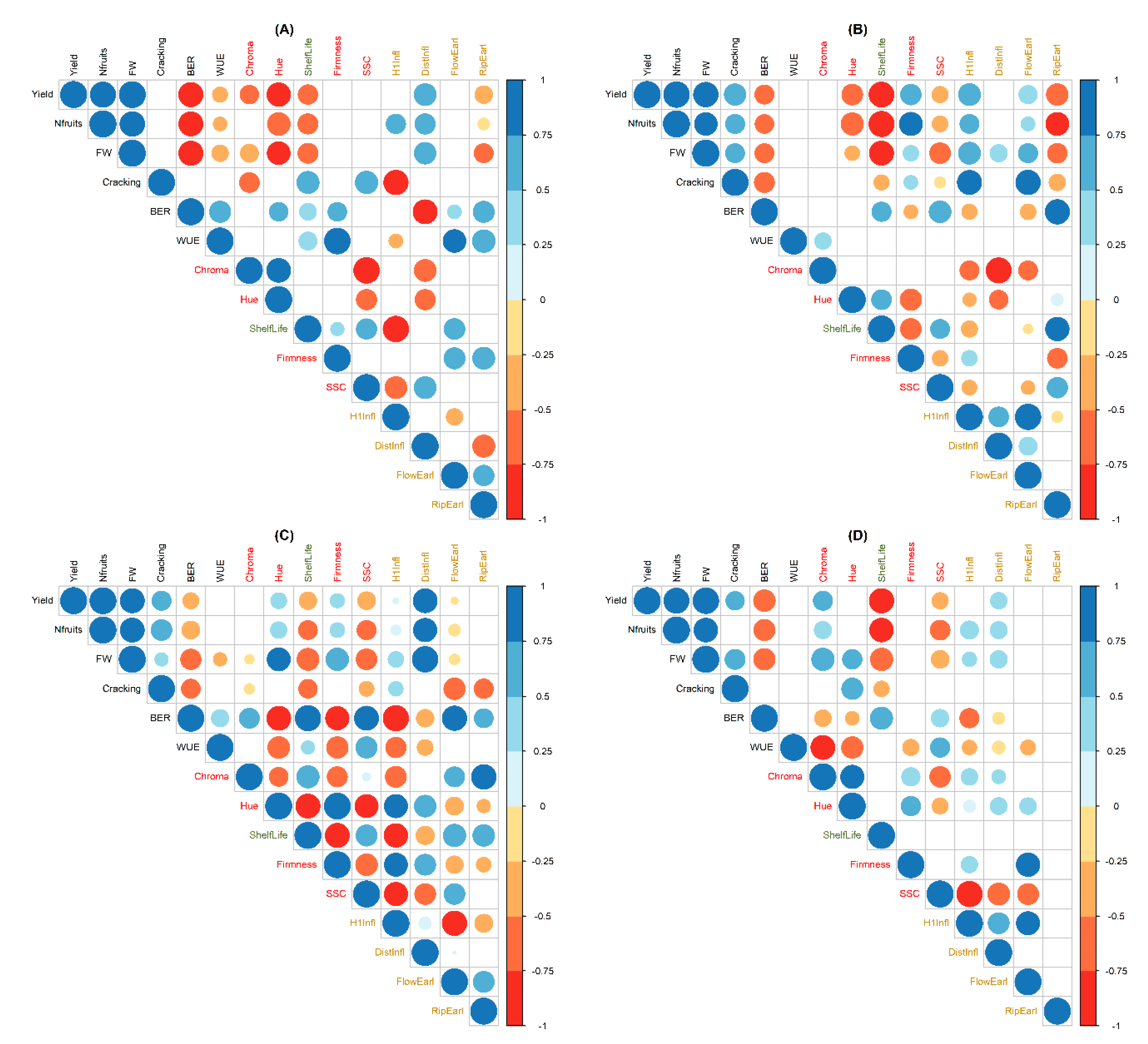
3.5. Trait Correlations
4. Discussion
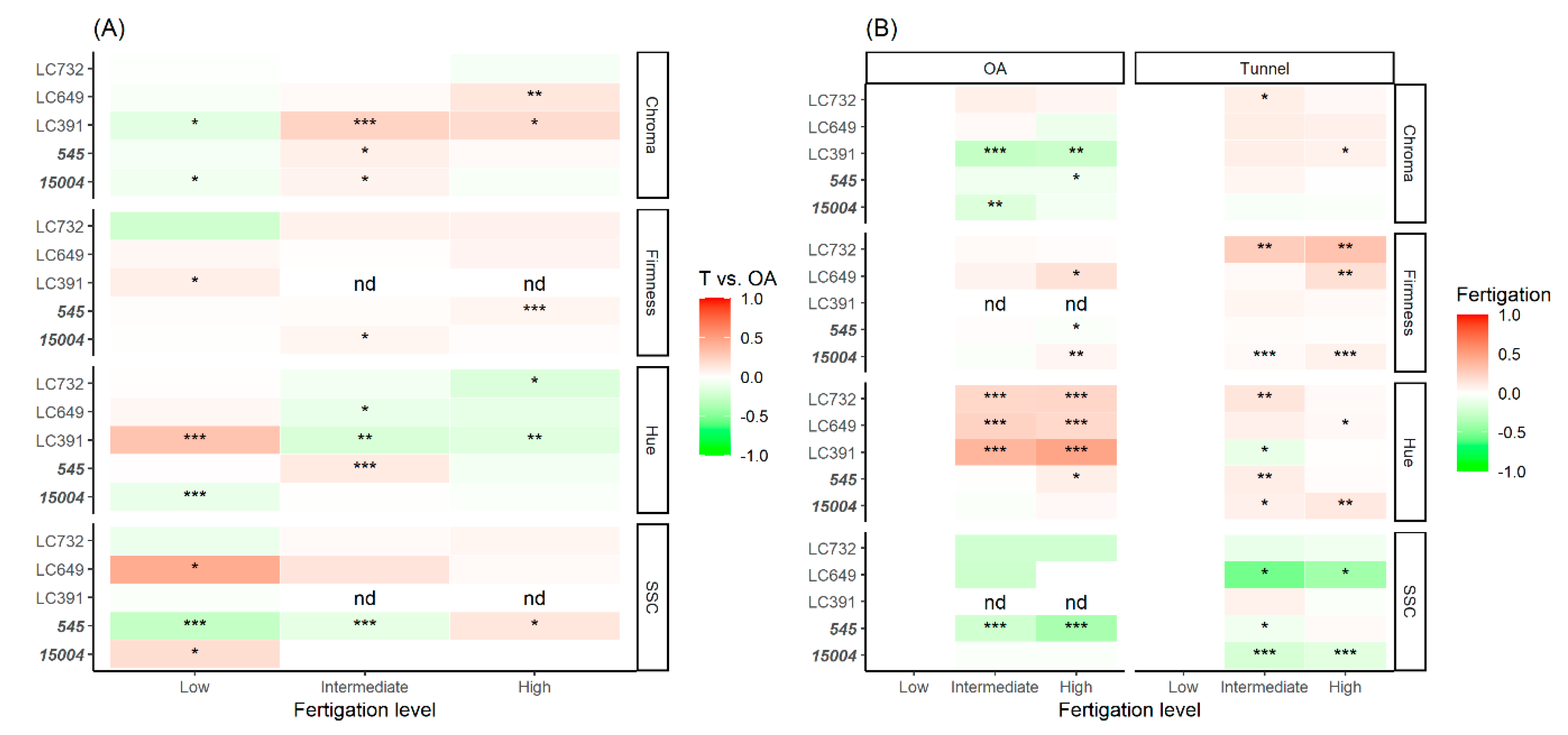
4.1. Differential Contributions of G, E, and GxE Interactions to Agronomic Performance, Plant Architecture, and Fruit-Quality Traits
4.2. Impact of Modern Breeding
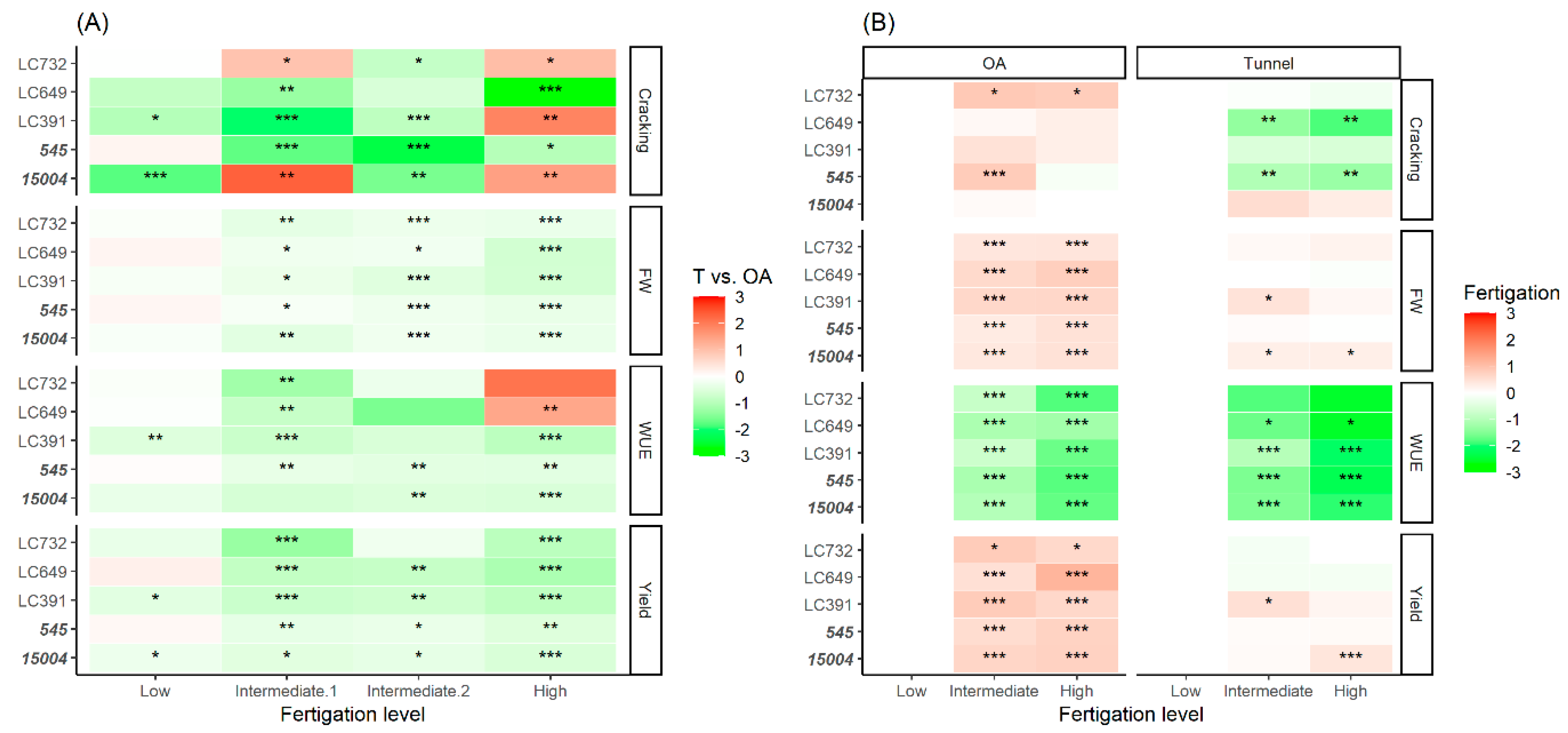
4.3. Effect of Protected Cultivation and Fertigation Dose on Yield, Quality, and Shelf-Life
4.4. Relationships between Plant Architecture and Earliness Components
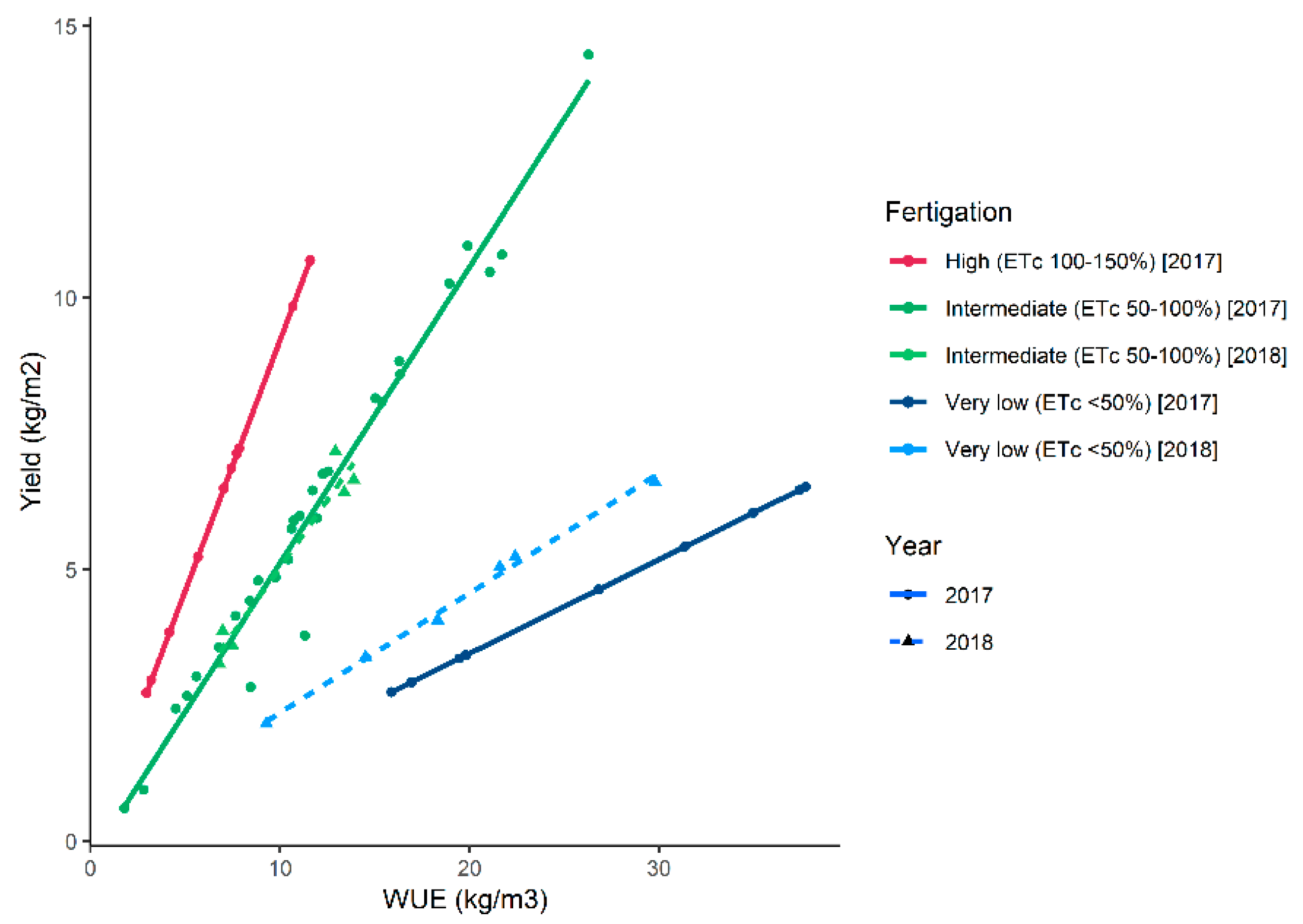
4.5. Effects of Cropping Systems on Water Use Efficiency
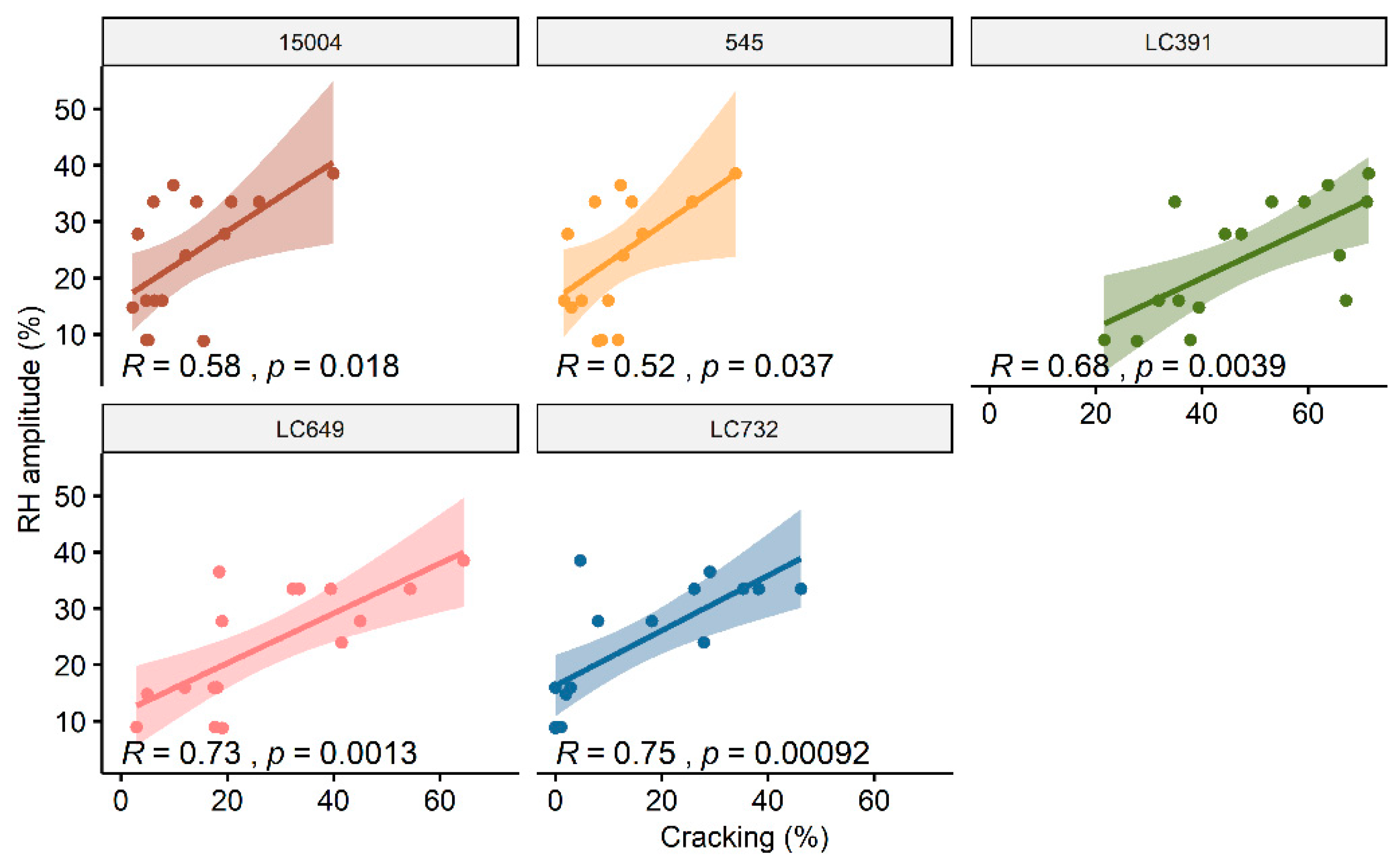
4.6. Genotype and Day-to-Night Variations in Relative Humidity Play an Important Role in the Incidence of Cracking
4.7. SSC and Shelf-Life Are Negatively Correlated with Yield
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- FAO. Food and Agriculture Organisation of the United Nations. Available online: http://faostat.fao.org (accessed on 6 December 2020).
- Causse, M.; Zhao, J.; Diouf, I.; Wang, J.; Lefebvre, V.; Caromel, B.; Génard, M.; Bertin, N. Genomic Designing for Climate-Smart Tomato. In Genomic Designing of Climate-Smart Vegetable Crops; Springer: Vienna, Austria, 2020; pp. 47–159. [Google Scholar]
- Heuvelink, E. (Ed.) Tomatoes; CABI: Wallingford, UK, 2018; ISBN 9781780641935. [Google Scholar]
- Bertin, N.; Génard, M. Tomato quality as influenced by preharvest factors. Sci. Hortic. 2018, 233, 264–276. [Google Scholar] [CrossRef]
- Beckles, D.M. Factors affecting the postharvest soluble solids and sugar content of tomato (Solanum lycopersicum L.) fruit. Postharvest Biol. Technol. 2012, 63, 129–140. [Google Scholar] [CrossRef]
- Dorais, M.; Papadopoulos, A.P.; Gosselin, A. Greenhouse Tomato Fruit Quality. Hortic. Rev. 2010, 26, 239–319. [Google Scholar] [CrossRef]
- Davies, J.N.; Hobson, G.E.; McGlasson, W.B. The constituents of tomato fruit—The influence of environment, nutrition, and genotype. Crit. Rev. Food Sci. Nutr. 1981, 15, 205–280. [Google Scholar] [CrossRef] [PubMed]
- Cuartero, J.; Cubero, J.I. Genotype-environment interaction in tomato. Theor. Appl. Genet. 1982, 61, 273–277. [Google Scholar] [CrossRef]
- Bertin, N.; Martre, P.; Génard, M.; Quilot, B.; Salon, C.; Quilot-Turion, B. Under what circumstances can process-based simulation models link genotype to phenotype for complex traits? Case-study of fruit and grain quality traits. J. Exp. Bot. 2009, 61, 955–967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, N.; Rivero, R.M.; Shulaev, V.; Blumwald, E.; Mittler, R. Abiotic and biotic stress combinations. N. Phytol. 2014, 203, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Zotarelli, L.; Dukes, M.; Scholberg, J.; Muñoz-Carpena, R.; Icerman, J. Tomato nitrogen accumulation and fertilizer use efficiency on a sandy soil, as affected by nitrogen rate and irrigation scheduling. Agric. Water Manag. 2009, 96, 1247–1258. [Google Scholar] [CrossRef]
- Wang, X.; Xing, Y. Evaluation of the effects of irrigation and fertilization on tomato fruit yield and quality: A principal component analysis. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Van Berloo, R.; Zhu, A.; Ursem, R.; Verbakel, H.; Gort, G.; Van Eeuwijk, F.A. Diversity and linkage disequilibrium analysis within a selected set of cultivated tomatoes. Theor. Appl. Genet. 2008, 117, 89–101. [Google Scholar] [CrossRef] [Green Version]
- Sim, S.-C.; Robbins, M.D.; Van Deynze, A.; Michel, A.P.; Francis, D.M. Population structure and genetic differentiation associated with breeding history and selection in tomato (Solanum lycopersicum L.). Heredity 2010, 106, 927–935. [Google Scholar] [CrossRef] [Green Version]
- Casals, J.; Rivera, A.; Sabaté, J.; Del Castillo, R.R.; Simó, J. Cherry and Fresh Market Tomatoes: Differences in Chemical, Morphological, and Sensory Traits and Their Implications for Consumer Acceptance. Agronomy 2018, 9, 9. [Google Scholar] [CrossRef] [Green Version]
- Grandillo, S.; Zamir, D.; Tanksley, S.D. Genetic improvement of processing tomatoes: A 20 years perspective. Euphytica 1999, 110, 85–97. [Google Scholar] [CrossRef]
- Caro, M.; Cruz, V.; Cuartero, J.; Estan, M.T.; Bolarin, M.C. Salinity tolerance of normal-fruited and cherry tomato cultivars. Plant Soil 1991, 136, 249–255. [Google Scholar] [CrossRef]
- Albert, E.; Segura, V.; Gricourt, J.; Bonnefoi, J.; Derivot, L.; Causse, M. Association mapping reveals the genetic architecture of tomato response to water deficit: Focus on major fruit quality traits. J. Exp. Bot. 2016, 67, 6413–6430. [Google Scholar] [CrossRef] [PubMed]
- Ho, L. The mechanism of assimilate partitioning and carbohydrate compartmentation in fruit in relation to the quality and yield of tomato. J. Exp. Bot. 1996, 47, 1239–1243. [Google Scholar] [CrossRef] [PubMed]
- Soyk, S.; Müller, N.A.; Park, S.J.; Schmalenbach, I.; Jiang, K.; Hayama, R.; Zhang, L.; Van Eck, J.; Jiménez-Gómez, J.M.; Lippman, Z.B. Variation in the flowering gene SELF PRUNING 5G promotes day-neutrality and early yield in tomato. Nat. Genet. 2016, 49, 162–168. [Google Scholar] [CrossRef]
- Conesa, M.À.; Fullana-Pericàs, M.; Granell, A.; Galmés, J. Mediterranean Long Shelf-Life Landraces: An Untapped Genetic Resource for Tomato Improvement. Front. Plant Sci. 2020, 10, 1651. [Google Scholar] [CrossRef]
- Casals, J.; Cebolla-Cornejo, J.; Roselló, S.; Beltran, J.; Casañas, F.; Nuez, F. Long-term postharvest aroma evolution of tomatoes with the alcobaça (alc) mutation. Eur. Food Res. Technol. 2011, 233, 331–342. [Google Scholar] [CrossRef] [Green Version]
- Bota, J.; Conesa, M.À.; Ochogavia, J.M.; Medrano, H.; Francis, D.M.; Cifre, J. Characterization of a landrace collection for Tomàtiga de Ramellet (Solanum lycopersicum L.) from the Balearic Islands. Genet. Resour. Crop. Evol. 2014, 61, 1131–1146. [Google Scholar] [CrossRef]
- Tranchida-Lombardo, V.; Cigliano, R.A.; Anzar, I.; Landi, S.; Palombieri, S.; Colantuono, C.; Bostan, H.; Termolino, P.; Aversano, R.; Batelli, G.; et al. Whole-genome re-sequencing of two Italian tomato landraces reveals sequence variations in genes associated with stress tolerance, fruit quality and long shelf-life traits. DNA Res. 2018, 25, 149–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casals, J.; Pascual, L.; Cañizares, J.; Cebolla-Cornejo, J.; Casañas, F.; Nuez, F. Genetic basis of long shelf life and variability into Penjar tomato. Genet. Resour. Crop. Evol. 2012, 59, 219–229. [Google Scholar] [CrossRef]
- Kopeliovitch, E.; Rabinowitch, H.D.; Mizrahi, Y.; Kedar, N. Mode of inheritance of Alcobaca, a tomato fruit-ripening mutant. Euphytica 1981, 30, 223–225. [Google Scholar] [CrossRef]
- Kumar, R.; Tamboli, V.; Sharma, R.; Sreelakshmi, Y. NAC-NOR mutations in tomato Penjar accessions attenuate multiple metabolic processes and prolong the fruit shelf life. Food Chem. 2018, 259, 234–244. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Wei, W.; Fan, Z.; Zhao, X.; Zhang, Y.; Jing, Y.; Zhu, B.; Zhu, H.; Shan, W.; Chen, J.; et al. Reevaluation of the nor mutation and the role of the NAC-NOR transcription factor in tomato fruit ripening. J. Exp. Bot. 2020, 71, 3560–3574. [Google Scholar] [CrossRef]
- Mercati, F.; Longo, C.; Poma, D.; Araniti, F.; Lupini, A.; Mammano, M.M.; Fiore, M.C.; Abenavoli, M.R.; Sunseri, F. Genetic variation of an Italian long shelf-life tomato (Solanum lycopersicon L.) collection by using SSR and morphological fruit traits. Genet. Resour. Crop. Evol. 2014, 62, 721–732. [Google Scholar] [CrossRef]
- Fullana-Pericàs, M.; Ponce, J.; Conesa, M.À.; Juan, A.; Ribas-Carbó, M.; Galmés, J. Changes in yield, growth and photosynthesis in a drought-adapted Mediterranean tomato landrace (Solanum lycopersicum ‘Ramellet’) when grafted onto commercial rootstocks and Solanum pimpinellifolium. Sci. Hortic. 2018, 233, 70–77. [Google Scholar] [CrossRef]
- Missio, J.C.; Renau, R.M.; Artigas, F.C.; Cornejo, J.C. Sugar and acid profile of Penjar tomatoes and its evolution during storage. Sci. Agric. 2015, 72, 314–321. [Google Scholar] [CrossRef]
- Conesa, M.À.; Galmés, J.; Ochogavía, J.M.; March, J.; Jaume, J.; Martorell, A.; Francis, D.M.; Medrano, H.; Rose, J.K.; Cifre, J. The postharvest tomato fruit quality of long shelf-life Mediterranean landraces is substantially influenced by irrigation regimes. Postharvest Biol. Technol. 2014, 93, 114–121. [Google Scholar] [CrossRef]
- Galmés, J.; Conesa, M.A.; Ochogavía, J.M.; Perdomo, J.A.; Francis, D.M.; Ribas-Carbó, M.; Savé, R.; Flexas, J.; Medrano, H.; Cifre, J. Physiological and morphological adaptations in relation to water use efficiency in Mediterranean accessions of Solanum lycopersicum. Plant Cell Environ. 2010, 34, 245–260. [Google Scholar] [CrossRef]
- Figàs, M.R.; Prohens, J.; Raigón, M.D.; Pereira-Dias, L.; Casanova, C.; García-Martínez, M.D.; Rosa, E.; Soler, E.; Plazas, M.; Soler, S. Insights into the Adaptation to Greenhouse Cultivation of the Traditional Mediterranean Long Shelf-Life Tomato Carrying the alc Mutation: A Multi-Trait Comparison of Landraces, Selections, and Hybrids in Open Field and Greenhouse. Front. Plant Sci. 2018, 9, 1774. [Google Scholar] [CrossRef]
- Casals, J.; Casañas, F.; Simó, J. Is it Still Necessary to Continue to Collect Crop Genetic Resources in the Mediterranean Area? A Case Study in Catalonia. Econ. Bot. 2017, 71, 330–341. [Google Scholar] [CrossRef]
- Poore, J.; Nemecek, T. Reducing food’s environmental impacts through producers and consumers. Science 2018, 360, 987–992. [Google Scholar] [CrossRef] [Green Version]
- Peet, M.M. Irrigation and fertilization. In Tomatoes; Heuvelink, E., Ed.; CABI: Wallingford, UK, 2005; pp. 171–198. [Google Scholar]
- De Pascale, S.; Rouphael, Y.; Gallardo, M.; Thompson, R. Water and fertilization management of vegetables: State of art and future challenges. Eur. J. Hortic. Sci. 2018, 83, 306–318. [Google Scholar] [CrossRef]
- Albornoz, F. Crop responses to nitrogen overfertilization: A review. Sci. Hortic. 2016, 205, 79–83. [Google Scholar] [CrossRef]
- Good, A.G.; Beatty, P.H. Fertilizing Nature: A Tragedy of Excess in the Commons. PLoS Biol. 2011, 9, e1001124. [Google Scholar] [CrossRef] [PubMed]
- Lemaire, G.; Tang, L.; Bélanger, G.; Zhu, Y.; Jeuffroy, M.-H. Forward new paradigms for crop mineral nutrition and fertilization towards sustainable agriculture. Eur. J. Agron. 2021, 125, 126248. [Google Scholar] [CrossRef]
- Peet, M. Fruit Cracking in Tomato. HortTechnology 1992, 2, 216–223. [Google Scholar] [CrossRef] [Green Version]
- Thole, V.; Vain, P.; Yang, R.; Da Silva, J.A.B.; Enfissi, E.M.A.; Nogueira, M.; Price, E.J.; Alseekh, S.; Fernie, A.R.; Fraser, P.D.; et al. Analysis of Tomato Post-Harvest Properties: Fruit Color, Shelf Life, and Fungal Susceptibility. Curr. Protoc. Plant Biol. 2020, 5, e20108. [Google Scholar] [CrossRef]
- Panthee, D.R.; Cao, C.; Debenport, S.J.; Rodríguez, G.R.; Labate, J.A.; Robertson, L.D.; Breksa, A.P.; Van Der Knaap, E.; Gardener, B.B.M. Magnitude of Genotype × Environment Interactions Affecting Tomato Fruit Quality. HortScience 2012, 47, 721–726. [Google Scholar] [CrossRef] [Green Version]
- Causse, M.; Damidaux, R.; Rousselle, P. Traditional and Enhanced Breeding for Quality Traits in Tomato. In Genetic Improvement of Solanaceous Crops; CRC Press: Boca Raton, FL, USA, 2006; Volume 2, pp. 153–192. [Google Scholar]
- Ortiz, R.; Crossa, J.; Vargas, M.; Izquierdo, J. Studying the effect of environmental variables on the genotype × environment interaction of tomato. Euphytica 2006, 153, 119–134. [Google Scholar] [CrossRef]
- Figàs, M.R.; Prohens, J.; Raigón, M.D.; Fita, A.; García-Martínez, M.D.; Casanova, C.; Borràs, D.; Plazas, M.; Andújar, I.; Soler, S. Characterization of composition traits related to organoleptic and functional quality for the differentiation, selection and enhancement of local varieties of tomato from different cultivar groups. Food Chem. 2015, 187, 517–524. [Google Scholar] [CrossRef] [Green Version]
- Casañas, F.; Simó, J.; Casals, J.; Prohens, J. Toward an Evolved Concept of Landrace. Front. Plant Sci. 2017, 8, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeven, A. Landraces: A review of definitions and classifications. Euphytica 1998, 104, 127–139. [Google Scholar] [CrossRef]
- De Gelder, A.; Dieleman, J.A.; Bot, G.; Marcelis, L.F.M. An overview of climate and crop yield in closed greenhouses. J. Hortic. Sci. Biotechnol. 2012, 87, 193–202. [Google Scholar] [CrossRef]
- O’Connell, S.; Rivard, C.; Peet, M.M.; Harlow, C.; Louws, F. High Tunnel and Field Production of Organic Heirloom Tomatoes: Yield, Fruit Quality, Disease, and Microclimate. HortScience 2012, 47, 1283–1290. [Google Scholar] [CrossRef] [Green Version]
- Erhioui, B.M.; Gosselin, A.; Hao, X.; Papadopoulos, A.P.; Dorais, M. Greenhouse Covering Materials and Supplemental Lighting Affect Growth, Yield, Photosynthesis, and Leaf Carbohydrate Synthesis of Tomato Plants. J. Am. Soc. Hortic. Sci. 2002, 127, 819–824. [Google Scholar] [CrossRef]
- Van Eeuwijk, F.A.; Bustos-Korts, D.; Millet, E.J.; Boer, M.P.; Kruijer, W.; Thompson, A.; Malosetti, M.; Iwata, H.; Quiroz, R.; Kuppe, C.; et al. Modelling strategies for assessing and increasing the effectiveness of new phenotyping techniques in plant breeding. Plant Sci. 2019, 282, 23–39. [Google Scholar] [CrossRef]
- Doganlar, S.; Tanksley, S.D.; Mutschler, M.A. Identification and molecular mapping of loci controlling fruit ripening time in tomato. Theor. Appl. Genet. 2000, 100, 249–255. [Google Scholar] [CrossRef]
- Powers, L.; Lyon, C.B. Inheritance studies on duration of developmental stages in crosses within the genus Lycopersicon. Agric. Res. 1941, 63, 129–148. [Google Scholar]
- Monforte, A.J.; Asíns, M.J.; Carbonell, E.A. Salt tolerance in Lycopersicon spp. VII. Pleiotropic action of genes controlling earliness on fruit yield. Theor. Appl. Genet. 1999, 98, 593–601. [Google Scholar] [CrossRef]
- Atherton, J.G.; Harris, G.P. Flowering. In The Tomato Crop: A Scientific Basis for Improvement; Atherton, J.G., Rudich, J., Eds.; Springer: Dordrecht, The Netherlands, 1986; pp. 167–200. ISBN 978-94-009-3137-4. [Google Scholar]
- Pnueli, L.; Carmel-Goren, L.; Hareven, D.; Gutfinger, T.; Alvarez, J.; Ganal, M.; Zamir, D.; Lifschitz, E. The SELF-PRUNING gene of tomato regulates vegetative to reproductive switching of sympodial meristems and is the ortholog of CEN and TFL1. Development 1998, 125, 1979–1989. [Google Scholar]
- Jones, C.M.; Rick, C.M.; Adams, D.; Jernstedt, J.; Chetelat, R.T. Genealogy and fine mapping of obscuravenosa, a gene affecting the distribution of chloroplasts in leaf veins, and evidence of selection during breeding of tomatoes (Lycopersicon esculentum; Solanaceae). Am. J. Bot. 2007, 94, 935–947. [Google Scholar] [CrossRef] [PubMed]
- Samach, A.; Lotan, H. The transition to flowering in tomato. Plant Biotechnol. 2007, 24, 71–82. [Google Scholar] [CrossRef] [Green Version]
- Heuvelink, E. Growth, development and yield of a tomato crop: Periodic destructive measurements in a greenhouse. Sci. Hortic. 1995, 61, 77–99. [Google Scholar] [CrossRef]
- Uzun, S. The quantitative effects of temperature and light on the number of leaves preceding the first fruiting inflorescence on the stem of tomato (Lycopersicon esculentum, Mill.) and aubergine (Solanum melongena L.). Sci. Hortic. 2006, 109, 142–146. [Google Scholar] [CrossRef]
- Dieleman, J.A.; Heuvelink, E. Factors affecting the number of leaves preceding the first inflorescence in the tomato. J. Hortic. Sci. 1992, 67, 1–10. [Google Scholar] [CrossRef]
- Pretty, J. Intensification for redesigned and sustainable agricultural systems. Science 2018, 362. [Google Scholar] [CrossRef] [Green Version]
- Cui, J.; Shao, G.; Lu, J.; Keabetswe, L.; Hoogenboom, G. Yield, quality and drought sensitivity of tomato to water deficit during different growth stages. Sci. Agric. 2020, 77. [Google Scholar] [CrossRef] [Green Version]
- Cantero-Navarro, E.; Romero-Aranda, R.; Fernández-Muñoz, R.; Martínez-Andújar, C.; Pérez-Alfocea, F.; Albacete, A. Improving agronomic water use efficiency in tomato by rootstock-mediated hormonal regulation of leaf biomass. Plant Sci. 2016, 251, 90–100. [Google Scholar] [CrossRef]
- Hooshmand, M.; Albaji, M.; Nasab, S.B.; Ansari, N.A.Z. The effect of deficit irrigation on yield and yield components of greenhouse tomato (Solanum lycopersicum) in hydroponic culture in Ahvaz region, Iran. Sci. Hortic. 2019, 254, 84–90. [Google Scholar] [CrossRef]
- Patanè, C.; Tringali, S.; Sortino, O. Effects of deficit irrigation on biomass, yield, water productivity and fruit quality of processing tomato under semi-arid Mediterranean climate conditions. Sci. Hortic. 2011, 129, 590–596. [Google Scholar] [CrossRef]
- Valcárcel, M.; Lahoz, I.; Campillo, C.; Martí, R.; Leiva-Brondo, M.; Roselló, S.; Cebolla-Cornejo, J. Controlled deficit irrigation as a water-saving strategy for processing tomato. Sci. Hortic. 2020, 261. [Google Scholar] [CrossRef]
- Chen, J.; Kang, S.; Du, T.; Qiu, R.; Guo, P.; Chen, R. Quantitative response of greenhouse tomato yield and quality to water deficit at different growth stages. Agric. Water Manag. 2013, 129, 152–162. [Google Scholar] [CrossRef]
- Fereres, E.; Soriano, M.A. Deficit irrigation for reducing agricultural water use. J. Exp. Bot. 2007, 58, 147–159. [Google Scholar] [CrossRef] [Green Version]
- Abbott, J.; Peet, M.; Willits, D.; Sanders, D.; Gough, R. Effects of irrigation frequency and scheduling on fruit production and radial fruit cracking in greenhouse tomatoes in soil beds and in a soil-less medium in bags. Sci. Hortic. 1986, 28, 209–217. [Google Scholar] [CrossRef]
- Lichter, A.; Dvir, O.; Fallik, E.; Cohen, S.; Golan, R.; Shemer, Z.; Sagi, M. Cracking of cherry tomatoes in solution. Postharvest Biol. Technol. 2002, 26, 305–312. [Google Scholar] [CrossRef]
- Matas, A.J.; López-Casado, G.; Cuartero, J.; Heredia, A. Relative humidity and temperature modify the mechanical properties of isolated tomato fruit cuticles. Am. J. Bot. 2005, 92, 462–468. [Google Scholar] [CrossRef]
- Correia, S.; Schouten, R.; Silva, A.P.; Gonçalves, B. Sweet cherry fruit cracking mechanisms and prevention strategies: A review. Sci. Hortic. 2018, 240, 369–377. [Google Scholar] [CrossRef]
- Opara, L.U.; Hodson, A.D.; Studman, S.P. Stem end splitting and internal ring-cracking of ‘Gala’ apples as influenced by orchard management practices. J. Hortic. Sci. Biotechnol. 2000, 75, 465–469. [Google Scholar] [CrossRef]
- Khadivi-Khub, A. Physiological and genetic factors influencing fruit cracking. Acta Physiol. Plant. 2015, 37, 1–14. [Google Scholar] [CrossRef]
- Domínguez, E.; Fernández, M.D.; Hernández, J.C.L.; Parra, J.P.; España, L.; Heredia, A.; Cuartero, J. Tomato fruit continues growing while ripening, affecting cuticle properties and cracking. Physiol. Plant. 2012, 146, 473–486. [Google Scholar] [CrossRef] [PubMed]
- Aloni, B.; Karni, L.; Rylski, I.; Cohen, Y.; Lee, Y.; Fuchs, M.; Moreshet, S.; Yao, C. Cuticular cracking in pepper fruit. I. Effects of night temperature and humidity. J. Hortic. Sci. Biotechnol. 1998, 73, 743–749. [Google Scholar] [CrossRef]
- Costan, A.; Stamatakis, A.; Chrysargyris, A.; Petropoulos, S.A.; Tzortzakis, N. Interactive effects of salinity and silicon application on Solanum lycopersicum growth, physiology and shelf-life of fruit produced hydroponically. J. Sci. Food Agric. 2020, 100, 732–743. [Google Scholar] [CrossRef]
- Arah, I.K.; Amaglo, H.; Kumah, E.K.; Ofori, H. Preharvest and Postharvest Factors Affecting the Quality and Shelf Life of Harvested Tomatoes: A Mini Review. Int. J. Agron. 2015, 2015, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Janse, J. Quality research of tomatoes. Anual Rep. Glas. Crop. Res. Stn. Naaldwijk 1988, 24–26. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trial | Year | Locality | Environment | Cycle | Cycle Length (DAS) | Water Requirements (ETc-Pr) (mm) (A) | Irrigation (mm) (B) | % Water Requirement (=B/A) | Nitrogen (N-N03) (kg/ha) | Potassium (K2O) (kg/ha) | Datasets Measured 1 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Exp1 | 2018 | Sta. Susanna | Tunnel | Early | 208 | 833 | 222 | 27 | 168 | 292 | B-G |
| Exp2 | 2018 | Sta. Susanna | Tunnel | Early | 208 | 833 | 555 | 67 | 408 | 713 | B-G |
| Exp3 | 2018 | Sta. Susanna | Tunnel | Intermediate | 184 | 729 | 479 | 66 | 339 | 580 | B-G |
| Exp4 | 2018 | Sta. Susanna | Tunnel | Intermediate | 184 | 729 | 233 | 32 | 152 | 289 | B-G |
| Exp5 | 2018 | Sta. Susanna | Open air | Intermediate | 184 | 636 | 233 | 37 | 152 | 289 | B-G |
| Exp6 | 2018 | Sta. Susanna | Open air | Intermediate | 184 | 636 | 479 | 75 | 339 | 580 | B-G |
| Exp7 | 2017 | Sta. Susanna | Open air | Intermediate | 186 | 648 | 922 | 142 | 868 | 1913 | A-G |
| Exp8 | 2017 | Sta. Susanna | Open air | Intermediate | 186 | 648 | 542 | 84 | 581 | 1366 | A-G |
| Exp9 | 2017 | Sta. Susanna | Open air | Intermediate | 186 | 648 | 173 | 27 | 157 | 342 | A-G |
| Exp10 | 2017 | Sta. Susanna | Tunnel | Intermediate | 176 | 776 | 922 | 119 | 868 | 1913 | A-G |
| Exp11 | 2017 | Sta. Susanna | Tunnel | Intermediate | 176 | 776 | 542 | 70 | 581 | 1366 | A-G |
| Exp12 | 2017 | Sta. Susanna | Tunnel | Intermediate | 176 | 776 | 1727 | 22 | 157 | 342 | A-G |
| Exp13 | 2017 | Palafolls | Tunnel | Early | 223 | 593 | 5502 | 93 | 201 | 456 | A-G |
| Exp14 | 2017 | Palafolls | Tunnel | Intermediate | 208 | 944 | 5259 | 56 | 329 | 546 | A-G |
| Exp15 | 2017 | Palafolls | Tunnel | Late | 181 | 528 | 3346 | 63 | 227 | 420 | A-G |
| Exp16 | 2017 | Palafolls | Open air | Intermediate | 172 | 746 | 4968 | 67 | 258 | 457 | A-G |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Casals, J.; Martí, M.; Rull, A.; Pons, C. Sustainable Transfer of Tomato Landraces to Modern Cropping Systems: The Effects of Environmental Conditions and Management Practices on Long-Shelf-Life Tomatoes. Agronomy 2021, 11, 533. https://doi.org/10.3390/agronomy11030533
Casals J, Martí M, Rull A, Pons C. Sustainable Transfer of Tomato Landraces to Modern Cropping Systems: The Effects of Environmental Conditions and Management Practices on Long-Shelf-Life Tomatoes. Agronomy. 2021; 11(3):533. https://doi.org/10.3390/agronomy11030533
Chicago/Turabian StyleCasals, Joan, Montserrat Martí, Aurora Rull, and Clara Pons. 2021. "Sustainable Transfer of Tomato Landraces to Modern Cropping Systems: The Effects of Environmental Conditions and Management Practices on Long-Shelf-Life Tomatoes" Agronomy 11, no. 3: 533. https://doi.org/10.3390/agronomy11030533