
Genetic Mapping of Quantitative Trait Loci for End-Use Quality and Grain Minerals in Hard Red Winter Wheat

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Germplasms and Field Trials
2.2. End-Use Quality Evaluations
2.3. Genotyping
2.4. Statistical and QTL analyses
3. Results
3.1. Analysis of Variance, Heritability, and Correlations
3.2. Consistent QTLs for End-Use Quality Traits
3.2.1. QTL for Kernel Characteristics and Flour Protein Content
3.2.2. QTLs Linked to Mixograph Parameters
3.2.3. QTLs for Grain Mineral Element Concentrations
3.3. Pleiotropic QTL
3.4. Interactions of Epistasis and Epistasis-by-Environment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ANOVA | analysis of variance |
| CTAB | cetyltrimethylammonium bromide |
| FPC | flour protein concentration |
| GCD | grain Cd concentration |
| GCO | grain Co concentration |
| GMO | grain Mo concentration |
| HDI | hardness index |
| HMW-GS | high molecular weight glutenin sub-unit |
| ICIM | inclusive composite interval mapping |
| KASP | Kompetitive Allele Specific PCR |
| KD | kernel diameter |
| MLI | midline left integral |
| MLS | midline left slope |
| MLT | midline left time |
| MLV | midline left value |
| MLW | midline left width |
| MMST | midline mixing stability or tolerance |
| MPI | midline peak integral |
| MPT | midline peak time |
| MPV | midline peak value |
| MPW | midline peak width |
| MRI | midline right integral |
| MRS | midline right slope |
| MRT | midline right time |
| MRV | midline right value |
| MRW | midline right width |
| MTI | midline tail integral |
| MTS | midline tail slope |
| MTV | midline tail value |
| MTW | midline tail width |
| MTX | midline time_X integral |
| MTXS | midline time_X slope |
| MTXV | midline time _X value |
| MTXW | midline time _X width |
| QTL | quantitative trait loci |
| RIL | recombinant inbred line |
| SKW | single kernel weight |
| WAB | water absorption |
References
- Braun, H.J.; Atlin, G.; Payne, T. Multi-location testing as a tool to identify plant response to global climate change. In Climate Change and Crop Production; Reynolds, M.P., Ed.; CAB International: Wallingford, UK, 2010; pp. 115–138. [Google Scholar]
- Dhakal, S.; Liu, X.; Girard, A.; Chu, C.; Yang, Y.; Wang, S.; Xue, Q.; Rudd, J.C.; Ibrahim, A.M.H.; Awika, J.M.; et al. Genetic dissection of end-use quality traits in two widely adapted wheat cultivars ‘TAM 111’ and ‘TAM 112’. Crop Sci. 2021, 61, 1944–1959. [Google Scholar] [CrossRef]
- AACC International. Approved methods of analysis, 11th ed.; American Association of Cereal Chemists (AACC) International: St. Paul, MN, USA, 2010. [Google Scholar]
- Morris, C.F.; Beecher, B.S. The distal portion of the short arm of wheat (Triticum aestivum L.) chromosome 5D controls endosperm vitreosity and grain hardness. Theor. Appl. Genet. 2012, 125, 247–254. [Google Scholar] [CrossRef]
- Goesaert, H.; Brijs, K.; Veraverbeke, W.; Courtin, C.; Gebruers, K.; Delcour, J. Wheat flour constituents: How they impact bread quality, and how to impact their functionality. Trends Food Sci. Technol. 2005, 16, 12–30. [Google Scholar] [CrossRef]
- Payne, P.I.; Nightingale, M.A.; Krattiger, A.F.; Holt, L.M. The relationship between HMW glutenin subunit composition and the bread-making quality of British-grown wheat varieties. J. Sci. Food Agric. 1987, 40, 51–65. [Google Scholar] [CrossRef]
- Shewry, P.R.; Halford, N.G.; Lafiandra, D. Genetics of wheat gluten proteins. Adv. Genet. 2003, 49, 111–184. [Google Scholar] [CrossRef]
- Wan, Y.; Shewry, P.R.; Hawkesford, M. A novel family of γ-gliadin genes are highly regulated by nitrogen supply in developing wheat grain. J. Exp. Bot. 2012, 64, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Zhang, K.; Dong, L.; Dong, Z.; Li, Y.; Hussain, A.; Zhai, H. Molecular genetic and genomic analysis of wheat milling and end-use traits in China: Progress and perspectives. Crop J. 2018, 6, 68–81. [Google Scholar] [CrossRef]
- Wang, W.; Guo, H.; Wu, C.; Yu, H.; Li, X.; Chen, G.; Tian, J.; Deng, Z. Identification of novel genomic regions associated with nine mineral elements in Chinese winter wheat grain. BMC Plant Biol. 2021, 21, 1–16. [Google Scholar] [CrossRef]
- Nawrot, T.; Plusquin, M.; Hogervorst, J.; A Roels, H.; Celis, H.; Thijs, L.; Vangronsveld, J.; Van Hecke, E.; Staessen, J.A. Environmental exposure to cadmium and risk of cancer: A prospective population-based study. Lancet Oncol. 2006, 7, 119–126. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, A.; Yamaji, N.; Yokosho, K.; Ma, J.F. Nramp5 is a major transporter responsible for manganese and cadmium uptake in rice. Plant Cell 2012, 24, 2155–2167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, W.-Y.; Mendoza-Cózatl, D.G.; Lee, Y.; Schroeder, J.I.; Ahn, S.-N.; Lee, H.-S.; Wicker, T.; Martinoia, E. Phytochelatin-metal(loid) transport into vacuoles shows different substrate preferences in barley and Arabidopsis. Plant Cell Environ. 2013, 37, 1192–1201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Feki, W.M.; Byrne, P.F.; Reid, S.D.; Lapitan, N.L.; Haley, S.D. Quantitative trait locus mapping for end-use quality traits in hard winter wheat under contrasting soil moisture levels. Crop Sci. 2013, 53, 1953–1967. [Google Scholar] [CrossRef]
- Echeverry-Solarte, M.; Kumar, A.; Kianian, S.; Simsek, S.; Alamri, M.S.; Mantovani, E.E.; McClean, P.E.; Deckard, E.L.; Elias, E.; Schatz, B.; et al. New QTL alleles for quality-related traits in spring wheat revealed by RIL population derived from supernumerary × non-supernumerary spikelet genotypes. Theor. Appl. Genet. 2015, 128, 893–912. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wu, B.; Singh, R.P.; Velu, G. QTL mapping for micronutrients concentration and yield component traits in a hexaploid wheat mapping population. J. Cereal Sci. 2019, 88, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Guttieri, M.J.; Baenziger, P.S.; Frels, K.; Carver, B.; Arnall, B.; Wang, S.; Akhunov, E.; Waters, B.M. Prospects for selecting wheat with increased zinc and decreased cadmium concentration in grain. Crop Sci. 2015, 55, 1712–1728. [Google Scholar] [CrossRef]
- Meng, L.; Li, H.; Zhang, L.; Wang, J. QTL IciMapping: Integrated software for genetic linkage map construction and quantitative trait locus mapping in biparental populations. Crop J. 2015, 3, 269–283. [Google Scholar] [CrossRef] [Green Version]
- Haley, S.; Martin, T.; Quick, J.; Seifers, D.; Stromberger, J.; Clayshulte, S.; Clifford, B.; Peairs, F.; Rudolph, J.; Johnson, J.; et al. Registration of CO960293-2 Wheat Germplasm Resistant to Wheat streak mosaic virus and Russian Wheat Aphid. Crop Sci. 2002, 42, 1381–1382. [Google Scholar] [CrossRef]
- Assanga, S.; Zhang, G.; Tan, C.; Rudd, J.C.; Ibrahim, A.; Xue, Q.; Chao, S.; Fuentealba, M.P.; Liu, S. Saturated genetic mapping of wheat streak mosaic virus resistance gene Wsm2 in wheat. Crop Sci. 2017, 57, 332–339. [Google Scholar] [CrossRef]
- Tan, C.; Assanga, S.; Zhang, G.; Rudd, J.C.; Haley, S.D.; Xue, Q.; Ibrahim, A.; Bai, G.; Zhang, X.; Byrne, P.; et al. Development and validation of KASP markers for wheat streak mosaic virus resistance gene Wsm2. Crop. Sci. 2017, 57, 340–349. [Google Scholar] [CrossRef] [Green Version]
- Lazar, M.; Worrall, W.; Peterson, G.; Fritz, A.; Marshall, D.; Nelson, L.; Rooney, L. Registration of ‘TAM 111’ wheat. Crop Sci. 2004, 44, 355–356. [Google Scholar] [CrossRef]
- Jondiko, T.O.; Yang, L.; Hays, D.B.; Ibrahim, A.M.; Tilley, M.; Awika, J.M. Prediction of wheat tortilla quality using multivariate modeling of kernel, flour, and dough properties. Innov. Food Sci. Emerg. Technol. 2016, 34, 9–15. [Google Scholar] [CrossRef]
- Guttieri, M.J.; Baenziger, P.S.; Frels, K.; Carver, B.; Arnall, B.; Waters, B.M. Variation for grain mineral concentration in a diversity panel of current and historical Great Plains hard winter wheat germplasm. Crop Sci. 2015, 55, 1035–1052. [Google Scholar] [CrossRef]
- Assanga, S.O.; Fuentealba, M.; Zhang, G.; Tan, C.; Dhakal, S.; Rudd, J.C.; Ibrahim, A.M.H.; Xue, Q.; Haley, S.; Chen, J.; et al. Mapping of quantitative trait loci for grain yield and its components in a US popular winter wheat TAM 111 using 90K SNPs. PLoS ONE 2017, 12, e0189669. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Christopher, M.D.; Griffey, C.A.; Hall, M.D.; Gundrum, P.G.; Brooks, W.S. Molecular characterization of resistance to Fusarium head blight in U.S. soft red winter wheat breeding line VA00W-38. Crop Sci. 2012, 52, 2283–2292. [Google Scholar] [CrossRef]
- Van Ooijen, J. JoinMap® 4, Software For The Calculation Of Genetic Linkage Maps In Experimental Populations; Kyazma BV: Wageningen, The Netherlands, 2006. [Google Scholar]
- Liu, S.; Assanga, S.O.; Dhakal, S.; Gu, X.; Tan, C.-T.; Yang, Y.; Rudd, J.; Hays, D.; Ibrahim, A.; Xue, Q.; et al. Validation of chromosomal locations of 90K array single nucleotide polymorphisms in US wheat. Crop Sci. 2016, 56, 364–373. [Google Scholar] [CrossRef] [Green Version]
- Mayer, K.X. The International Wheat Genome Sequencing Consortium (IWGSC). A chromosome-based draft sequence of the hexaploid bread wheat (Triticum aestivum) genome. Science 2014, 345, 1251788. [Google Scholar] [CrossRef]
- SAS Institute Inc. SAS/STAT® 14.1 User’s Guide; SAS Institute Inc.: Cary, NC, USA, 2015. [Google Scholar]
- Fehr, W.R.; Fehr, E.L.; Jessen, H.J. Principles of Cultivar Development: Theory and Technique; Macmillan: New York, NY, USA, 1987; Volume 1. [Google Scholar]
- Yang, Y.; Dhakal, S.; Chu, C.; Wang, S.; Xue, Q.; Rudd, J.C.; Ibrahim, A.M.H.; Jessup, K.; Baker, J.; Fuentealba, M.P.; et al. Genome wide identification of QTL associated with yield and yield components in two popular wheat cultivars TAM 111 and TAM 112. PLoS ONE 2020, 15, e0237293. [Google Scholar] [CrossRef]
- Barakat, M.; Al-Doss, A.; Moustafa, K.; Motawei, M.; Alamri, M.; Mergoum, M.; Sallam, M.; Al-Ashkar, I. QTL analysis of farinograph and mixograph related traits in spring wheat under heat stress conditions. Mol. Biol. Rep. 2020, 47, 5477–5486. [Google Scholar] [CrossRef] [PubMed]
- Tsilo, T.J.; Simsek, S.; Ohm, J.-B.; Hareland, G.A.; Chao, S.; Anderson, J.A. Quantitative trait loci influencing endosperm texture, dough-mixing strength, and bread-making properties of the hard red spring wheat breeding lines. Genome 2011, 54, 460–470. [Google Scholar] [CrossRef]
- Zhao, F.J.; Su, Y.H.; Dunham, S.J.; Rakszegi, M.; Bedo, Z.; McGrath, S.P.; Shewry, P.R. Variation in mineral micronutrient concentrations in grain of wheat lines of diverse origin. J. Cereal Sci. 2009, 49, 290–295. [Google Scholar] [CrossRef]
- Morgounov, A.; Gómez-Becerra, H.F.; Abugalieva, A.; Dzhunusova, M.; Yessimbekova, M.; Muminjanov, H.; Zelenskiy, Y.; Ozturk, L.; Cakmak, I. Iron and zinc grain density in common wheat grown in Central Asia. Euphytica 2006, 155, 193–203. [Google Scholar] [CrossRef] [Green Version]
- Ibba, M.I.; Kiszonas, A.; See, D.R.; Skinner, D.Z.; Morris, C.F. Mapping kernel texture in a soft durum (Triticum turgidum subsp. durum) wheat population. J. Cereal Sci. 2019, 85, 20–26. [Google Scholar] [CrossRef]
- Smith, N.; Souza, E.; Sneller, C.; Sorrells, M.; Griffey, C.; Ohm, H.; Van Sanford, D.; Guttieri, M.J.; Sturbaum, A. Association analysis of soft wheat quality traits in Eastern US soft winter wheat. In Proceedings of the 2008 Joint Annual Meeting of ASA-CSSA-SSSA, Houston, TX, USA, 5–9 October 2008. [Google Scholar]
- Lesage, V.; Rhazi, L.; Aussenac, T.; Meleard, B.; Branlard, G. Effects of HMW- and LMW-glutenins and grain hardness on size of gluten polymers. In Proceedings of the 11th International Gluten Workshop, Beijing, China, 12–15 August 2012; He, Z., Wang, D., Eds.; CIMMYT: El Batán, Mexico, 2012; pp. 200–205. [Google Scholar]
- Arbelbide, M.; Bernardo, R. Mixed-model QTL mapping for kernel hardness and dough strength in bread wheat. Theor. Appl. Genet. 2006, 112, 885–890. [Google Scholar] [CrossRef] [PubMed]
- Semagn, K.; Iqbal, M.; Chen, H.; Perez-Lara, E.; Bemister, D.H.; Xiang, R.; Zou, J.; Asif, M.; Kamran, A.; N’Diaye, A.; et al. Physical mapping of QTL associated with agronomic and end-use quality traits in spring wheat under conventional and organic management systems. Theor. Appl. Genet. 2021, 134, 3699–3719. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Marza, F.; Ma, H.; Carver, B.F.; Bai, G. Mapping quantitative trait loci for quality factors in an inter-class cross of US and Chinese wheat. Theor. Appl. Genet. 2010, 120, 1041–1051. [Google Scholar] [CrossRef]
- Maphosa, L.; Langridge, P.; Taylor, H.; Parent, B.; Emebiri, L.; Kuchel, H.; Reynolds, M.P.; Chalmers, K.J.; Okada, A.; Edwards, J.; et al. Genetic control of grain yield and grain physical characteristics in a bread wheat population grown under a range of environmental conditions. Theor. Appl. Genet. 2014, 127, 1607–1624. [Google Scholar] [CrossRef]
- Huang, X.Q.; Cloutier, S.; Lycar, L.; Radovanovic, N.; Humphreys, D.G.; Noll, J.S.; Somers, D.J.; Brown, P.D. Molecular detection of QTLs for agronomic and quality traits in a doubled haploid population derived from two Canadian wheats (Triticum aestivum L.). Theor. Appl. Genet. 2006, 113, 753–766. [Google Scholar] [CrossRef]
- Alemu, A.; El Baouchi, A.; El Hanafi, S.; Kehel, Z.; Eddakhir, K.; Tadesse, W. Genetic analysis of grain protein content and dough quality traits in elite spring bread wheat (Triticum aestivum) lines through association study. J. Cereal Sci. 2021, 100, 103214. [Google Scholar] [CrossRef]
- Naraghi, S.M.; Simsek, S.; Kumar, A.; Al Rabbi, S.M.H.; Alamri, M.S.; Elias, E.M.; Mergoum, M. Deciphering the genetics of major end-use quality traits in wheat. G3 Genes Genomes Genet. 2019, 9, 1405–1427. [Google Scholar] [CrossRef] [Green Version]
- Prashant, R.; Mani, E.; Rai, R.; Gupta, R.; Tiwari, R.; Dholakia, B.; Oak, M.; Röder, M.; Kadoo, N.; Gupta, V. Genotype × environment interactions and QTL clusters underlying dough rheology traits in Triticum aestivum L. J. Cereal Sci. 2015, 64, 82–91. [Google Scholar] [CrossRef]
- Akiyama, T.; Pillai, M.A.; Sentoku, N. Cloning, characterization and expression of OsGLN2, a rice endo-1,3-β-glucanase gene regulated developmentally in flowers and hormonally in germinating seeds. Planta 2004, 220, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Aoun, M.; Carter, A.H.; Ward, B.P.; Morris, C.F. Genome-wide association mapping of the ‘super-soft’ kernel texture in white winter wheat. Theor. Appl. Genet. 2021, 134, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhou, R.; Wang, J.; Liao, X.; Branlard, G.; Jia, J. Novel and favorable QTL allele clusters for end-use quality revealed by introgression lines derived from synthetic wheat. Mol. Breed. 2011, 29, 627–643. [Google Scholar] [CrossRef]
- Sherman, J.D.; Nash, D.; Lanning, S.P.; Martin, J.M.; Blake, N.K.; Morris, C.F.; Talbert, L.E. Genetics of end-use quality differences between a modern and historical spring wheat. Crop Sci. 2014, 54, 1972–1980. [Google Scholar] [CrossRef]
- Chen, Q.; Zhang, W.; Gao, Y.; Yang, C.; Gao, X.; Peng, H.; Hu, Z.; Xin, M.; Ni, Z.; Zhang, P.; et al. High molecular weight glutenin subunits 1Bx7 and 1By9 encoded by Glu-B1 locus affect wheat dough properties and sponge cake quality. J. Agric. Food Chem. 2019, 67, 11796–11804. [Google Scholar] [CrossRef]
- Jin, H.; Zhang, Y.; Li, G.; Mu, P.; Fan, Z.; Xia, X.; He, Z. Effects of allelic variation of HMW-GS and LMW-GS on mixograph properties and Chinese noodle and steamed bread qualities in a set of Aroona near-isogenic wheat lines. J. Cereal Sci. 2013, 57, 146–152. [Google Scholar] [CrossRef]
- I Payne, P. Genetics of wheat storage proteins and the effect of allelic variation on bread-making quality. Annu. Rev. Plant Physiol. 1987, 38, 141–153. [Google Scholar] [CrossRef]
- Liang, D.; Tang, J.; Peña, R.J.; Singh, R.; He, X.; Shen, X.; Yao, D.; Xia, X.; He, Z. Characterization of CIMMYT bread wheats for high- and low-molecular weight glutenin subunits and other quality-related genes with SDS-PAGE, RP-HPLC and molecular markers. Euphytica 2009, 172, 235–250. [Google Scholar] [CrossRef]
- Payne, P.; Holt, L.; Krattiger, A.; Carrillo, J. Relationships between seed quality characteristics and HMW glutenin subunit composition determined using wheats grown in Spain. J. Cereal Sci. 1988, 7, 229–235. [Google Scholar] [CrossRef]
- Dong, L.; Huo, N.; Wang, Y.; Deal, K.; Wang, D.; Hu, T.; Dvorak, J.; Anderson, O.D.; Luo, M.; Gu, Y.Q. Rapid evolutionary dynamics in a 2.8-Mb chromosomal region containing multiple prolamin and resistance gene families in Aegilops tauschii. Plant J. 2016, 87, 495–506. [Google Scholar] [CrossRef]
- Juliana, P.; Poland, J.; Huerta-Espino, J.; Shrestha, S.; Crossa, J.; Crespo-Herrera, L.; Toledo, F.H.; Govindan, V.; Mondal, S.; Kumar, U.; et al. Improving grain yield, stress resilience and quality of bread wheat using large-scale genomics. Nat. Genet. 2019, 51, 1530–1539. [Google Scholar] [CrossRef]
- Asri, N.; Rostami-Nejad, M.; Anderson, R.P.; Rostami, K. The Gluten Gene: Unlocking the understanding of gluten sensitivity and intolerance. Appl. Clin. Genet. 2021, 14, 37–50. [Google Scholar] [CrossRef] [PubMed]
- Stein, A.; Qaim, M. The human and economic cost of hidden hunger. Food Nutr. Bull. 2007, 28, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Wessells, K.R.; Brown, K.H. Estimating the global prevalence of zinc deficiency: Results based on zinc availability in national food supplies and the prevalence of stunting. PLoS ONE 2012, 7, e50568. [Google Scholar] [CrossRef] [Green Version]
- Gupta, P.K.; Balyan, H.S.; Sharma, S.; Kumar, R. Biofortification and bioavailability of Zn, Fe and Se in wheat: Present status and future prospects. Theor. Appl. Genet. 2021, 134, 1–35. [Google Scholar] [CrossRef]
- Crespo-Herrera, L.; Velu, G.; Singh, R. Quantitative trait loci mapping reveals pleiotropic effect for grain iron and zinc concentrations in wheat. Ann. Appl. Biol. 2016, 169, 27–35. [Google Scholar] [CrossRef]
- A Novotny, J.; A Peterson, C. Molybdenum. Adv. Nutr. 2018, 9, 272–273. [Google Scholar] [CrossRef]
- Kaiser, B.N.; Gridley, K.L.; Brady, J.N.; Phillips, T.; Tyerman, S. The role of molybdenum in agricultural plant production. Ann. Bot. 2005, 96, 745–754. [Google Scholar] [CrossRef] [PubMed]
- Descalsota-Empleo, G.I.; Amparado, A.; Inabangan-Asilo, M.A.; Tesoro, F.; Stangoulis, J.; Reinke, R.; Swamy, B.M. Genetic mapping of QTL for agronomic traits and grain mineral elements in rice. Crop J. 2019, 7, 560–572. [Google Scholar] [CrossRef]
- Wang, C.; Tang, Z.; Zhuang, J.-Y.; Tang, Z.; Huang, X.-Y.; Zhao, F.-J. Genetic mapping of ionomic quantitative trait loci in rice grain and straw reveals OsMOT1;1 as the putative causal gene for a molybdenum QTL qMo8. Mol. Genet. Genom. 2019, 295, 391–407. [Google Scholar] [CrossRef]
- Bhatta, M.; Baenziger, P.S.; Waters, B.M.; Poudel, R.; Belamkar, V.; Poland, J.; Morgounov, A. Genome-wide association study reveals novel genomic regions associated with 10 grain minerals in synthetic hexaploid wheat. Int. J. Mol. Sci. 2018, 19, 3237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knox, R.E.; Pozniak, C.J.; Clarke, F.R.; Clarke, J.M.; Houshmand, S.; Singh, A.K. Chromosomal location of the cadmium uptake gene (Cdu1) in durum wheat. Genome 2009, 52, 741–747. [Google Scholar] [CrossRef] [PubMed]
- Bálint, A.F.; Szira, F.; Röder, M.S.; Galiba, G.; Börner, A. Mapping of loci affecting copper tolerance in wheat—the possible impact of the vernalization gene Vrn-A1. Environ. Exp. Bot. 2009, 65, 369–375. [Google Scholar] [CrossRef]



{kind=link}
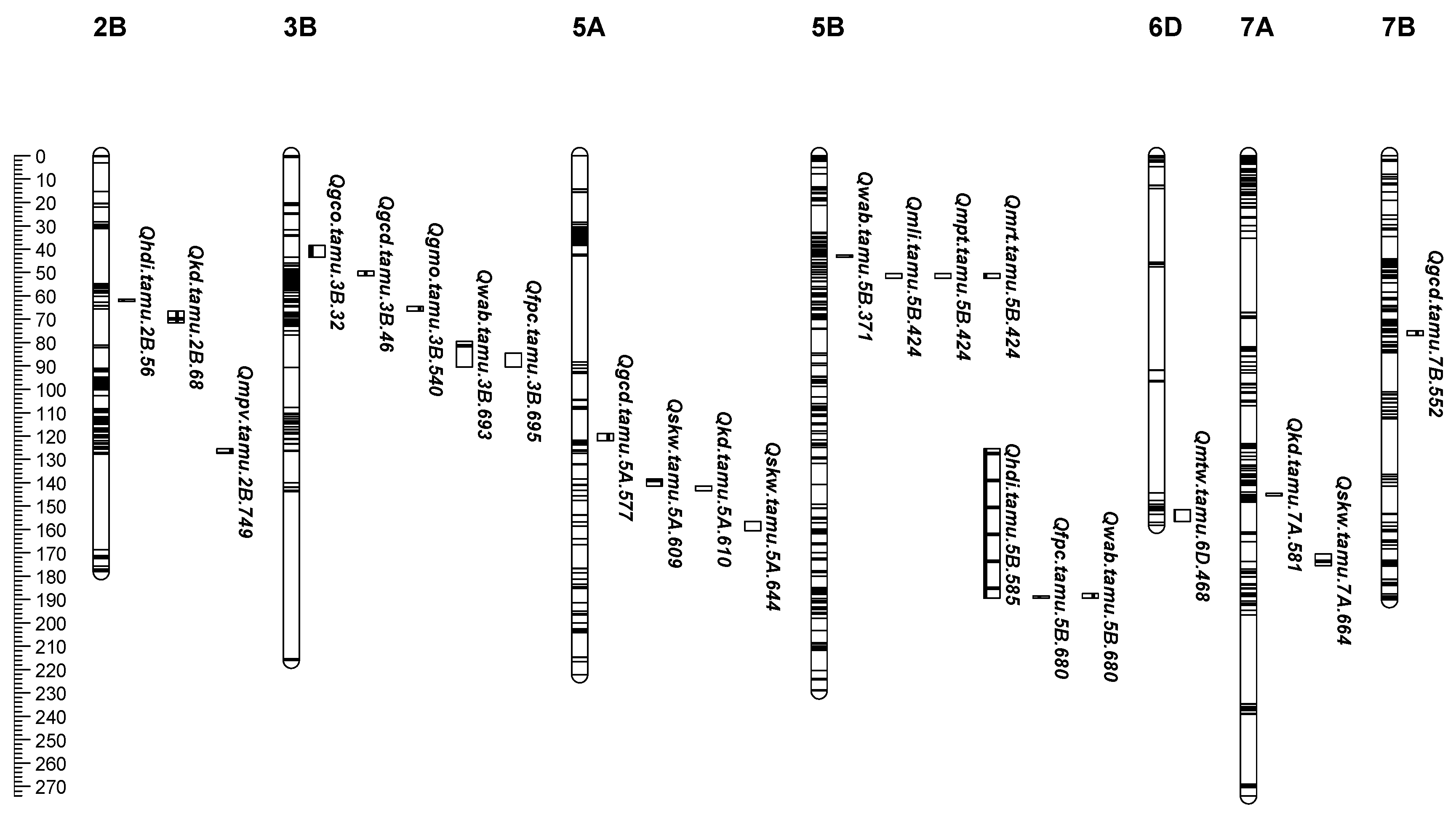{kind=link}
| QTL Name | Chr | Peak (Mbp) | Trait a | ENV b | LOD Threshold | LOD c | LOD (A) | LOD (AbyE) | PVE d | PVE (A) | PVE (AbyE) | Additive | Alleles Increase trait e | Pleiotropic QTL |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Qhdi.tamu.2B.56 | 2B | 56 | HDI | MET-ADD, BS14, ET14 | 3.2–4.6 | 9.0–23.8 | 16.6 | 7.2 | 16.7–20.3 | 12.6 | 6.2 | 1.4–2.4 | CO960293-2 | |
| Qkd.tamu.2B.68 | 2B | 68 | KD | MET-ADD, ET14, HY13 | 3.2–8.0 | 3.7–10.0 | 4.5 | 5.5 | 6.1–8.3 | 3.7 | 4.5 | −0.01 | TAM 111 | |
| Qfpc.tamu.3B.695 | 3B | 695 | FPC | MET-ADD, BS14, ET14 | 3.2–4.5 | 5.4–13.3 | 9.5 | 3.8 | 9.1–11.3 | 8.9 | 2.4 | −0.1–(−0.2) | TAM 111 | Y |
| Qmpt.tamu.1A.3 | 1A | 3 | MPT | MET-ADD, BS14, ET14 | 3.2–4.6 | 8.9–12.8 | 9.2 | 3.6 | 5.2–7.1 | 3.6 | 1.7 | −0.2–(−0.4) | TAM 111 | Y |
| Qmpt.tamu.1B.5 | 1B | 5 | MPT | MET-ADD, BS14, ET14, HY13 | 3.2–8.0 | 14.4–59.8 | 57.2 | 2.5 | 17.3–28.8 | 27.7 | 1.1 | 0.5–0.8 | CO960293-2 | Y |
| Qmpt.tamu.1D.417 | 1D | 417 | MPT | MET-ADD, BS14, HY13 | 3.2–8.0 | 28.8–50.0 | 40.7 | 9.3 | 27.6–34.2 | 17.8 | 9.8 | 0.5–1.0 | CO960293-2 | Y |
| Qmlt.tamu.1B.5 | 1B | 5 | MLT | MET-ADD, BS14, ET14, HY13 | 3.2–8.0 | 20.6–47.4 | 46.8 | 0.6 | 20.1–27.2 | 25.5 | 1.7 | 0.6–0.8 | CO960293-2 | Y |
| Qmlt.tamu.1D.1 | 1D | 1 | MLT | MET-ADD, BS14, ET14 | 3.2–4.6 | 4.8–7.9 | 7.6 | 0.4 | 3.6–4.3 | 3.4 | 0.9 | −0.2–(−0.3) | TAM 111 | Y |
| Qmli.tamu.1B.5 | 1B | 5 | MLI | MET-ADD, BS14, ET14, HY13 | 3.2–8.0 | 13.7–47.0 | 45.6 | 1.4 | 13.4–27.1 | 26.3 | 0.8 | 19.6–24.2 | CO960293-2 | Y |
| Qmls.tamu.1B.5 | 1B | 5 | MLS | MET-ADD, BS14, ET14, HY13 | 3.2–8.0 | 10.2–15.6 | 11.2 | 4.4 | 14.2–21.9 | 14.0 | 7.9 | −1.1–(−3.2) | TAM 111 | Y |
| Qmpi.tamu.1B.5 | 1B | 5 | MPI | MET-ADD, BS14, ET14, HY13 | 3.2–8.0 | 9.0–44.0 | 40.3 | 3.7 | 10.3–22.1 | 21.7 | 0.4 | 18.4–23.5 | CO960293-2 | Y |
| Qmrt.tamu.1B.5 | 1B | 5 | MRT | MET-ADD, BS14, ET14 | 3.2–4.5 | 6.3–19.3 | 17.3 | 2.0 | 9.3–18.8 | 16.1 | 2.7 | 0.3–0.4 | CO960293-2 | Y |
| Qmli.tamu.1D.413 | 1D | 413 | MLI | MET-ADD, BS14, ET14 | 3.2–4.8 | 29.2–36.2 | 12.5 | 16.7 | 17.6–31.3 | 6.3 | 11.3 | 9.6–29.4 | CO960293-2 | Y |
| Qmtw.tamu.1D.413 | 1D | 413 | MTW | MET-ADD, BS14, ET14 | 3.2–4.5 | 9.1–10.4 | 3.3 | 5.7 | 9.7–13.6 | 3.1 | 6.6 | 0.5–2.0 | CO960293-2 | Y |
| Qmtxw.tamu.1D.413 | 1D | 413 | MTXW | MET-ADD, BS14, ET14 | 3.2–4.6 | 7.5–10.4 | 2.8 | 4.7 | 7.4–13.6 | 2.4 | 5.1 | 0.5–2.0 | CO960293-2 | Y |
| Qmli.tamu.1A.3 | 1A | 3 | MLI | ET14, BS14 | 3.2 | 10.7–15.8 | 8.4~11.1 | −5.4–(−17.4) | TAM 111 | Y | ||||
| Qmpi.tamu.1A.5 | 1A | 5 | MPI | MET-ADD, BS14, ET14 | 3.2–4.5 | 11.5–14.0 | 10.7 | 3.3 | 6.7–10.6 | 5.1 | 1.6 | −9.3–(−17.3) | TAM 111 | |
| Qmrs.tamu.1B.8 | 1B | 8 | MRS | MET-ADD, BS14, ET14 | 3.2–4.5 | 5.6–12.6 | 12.1 | 0.5 | 9.5–14.7 | 11.0 | 3.7 | 0.3–0.5 | CO960293-2 | Y |
| Qmpi.tamu.1D.66 | 1D | 66 | MPI | MET-ADD, BS14, HY13 | 3.2–8.0 | 14.6–40.7 | 29.7 | 11.0 | 22.9–30.9 | 15.4 | 8.0 | 16.2–29.7 | CO960293-2 | Y |
| Qmpi.tamu.1D.1 | 1D | 1 | MPI | MET-ADD, BS14, ET14 | 3.2–4.5 | 5.7–6.3 | 4.4 | 1.9 | 2.9–3.9 | 2.0 | 0.8 | −5.83–(−10.5) | TAM 111 | Y |
| Qmtw.tamu.1D.1 | 1D | 1 | MTW | MET-ADD, BS14, ET14 | 3.2–4.5 | 6.5–9.2 | 7.0 | 2.2 | 6.9–9.8 | 6.5 | 3.3 | −0.8–(−1.6) | TAM 111 | Y |
| Qmtxw.tamu.1D.1 | 1D | 1 | MTXW | MET-ADD, BS14, ET14 | 3.2–4.6 | 6.5–7.9 | 6.2 | 1.7 | 6.9–8.9 | 5.4 | 2.5 | −0.7–(−1.6) | TAM 111 | Y |
| Qgco.tamu.3B.32 | 3B | 32 | GCO | ET14 | 3.4 | 5.6 | 11.6 | −0.003 | TAM 111 | |||||
| Qgcd.tamu.3B.46 | 3B | 46 | GCD | ET14 | 3.4 | 5.8 | 8.0 | −0.006 | TAM 111 | |||||
| Qgmo.tamu.3B.540 | 3B | 540 | GMO | ET14 | 3.4 | 4.8 | 8.6 | 0.036 | CO960293-2 | |||||
| Qgcd.tamu.5A.577 | 5A | 577 | GCD | ET14 | 3.4 | 5.5 | 8.0 | −0.006 | TAM 111 | |||||
| Qgcd.tamu.7B.552 | 7B | 552 | GCD | ET14 | 3.4 | 5.4 | 7.6 | −0.006 | TAM 111 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, S.; Assanga, S.O.; Awika, J.M.; Ibrahim, A.M.H.; Rudd, J.C.; Xue, Q.; Guttieri, M.J.; Zhang, G.; Baker, J.A.; Jessup, K.E.; et al. Genetic Mapping of Quantitative Trait Loci for End-Use Quality and Grain Minerals in Hard Red Winter Wheat. Agronomy 2021, 11, 2519. https://doi.org/10.3390/agronomy11122519
Yu S, Assanga SO, Awika JM, Ibrahim AMH, Rudd JC, Xue Q, Guttieri MJ, Zhang G, Baker JA, Jessup KE, et al. Genetic Mapping of Quantitative Trait Loci for End-Use Quality and Grain Minerals in Hard Red Winter Wheat. Agronomy. 2021; 11(12):2519. https://doi.org/10.3390/agronomy11122519
Chicago/Turabian StyleYu, Shuhao, Silvano O. Assanga, Joseph M. Awika, Amir M. H. Ibrahim, Jackie C. Rudd, Qingwu Xue, Mary J. Guttieri, Guorong Zhang, Jason A. Baker, Kirk E. Jessup, and et al. 2021. "Genetic Mapping of Quantitative Trait Loci for End-Use Quality and Grain Minerals in Hard Red Winter Wheat" Agronomy 11, no. 12: 2519. https://doi.org/10.3390/agronomy11122519