
Investigation of Citrus HLB Symptom Variations Associated with “Candidatus Liberibacter asiaticus” Strains Harboring Different Phages in Southern China

Abstract
:1. Introduction
2. Materials and Methods
2.1. Source of CLas Strains and Phage Types
2.2. Graft Transmission Experiments
2.3. PCR Assay
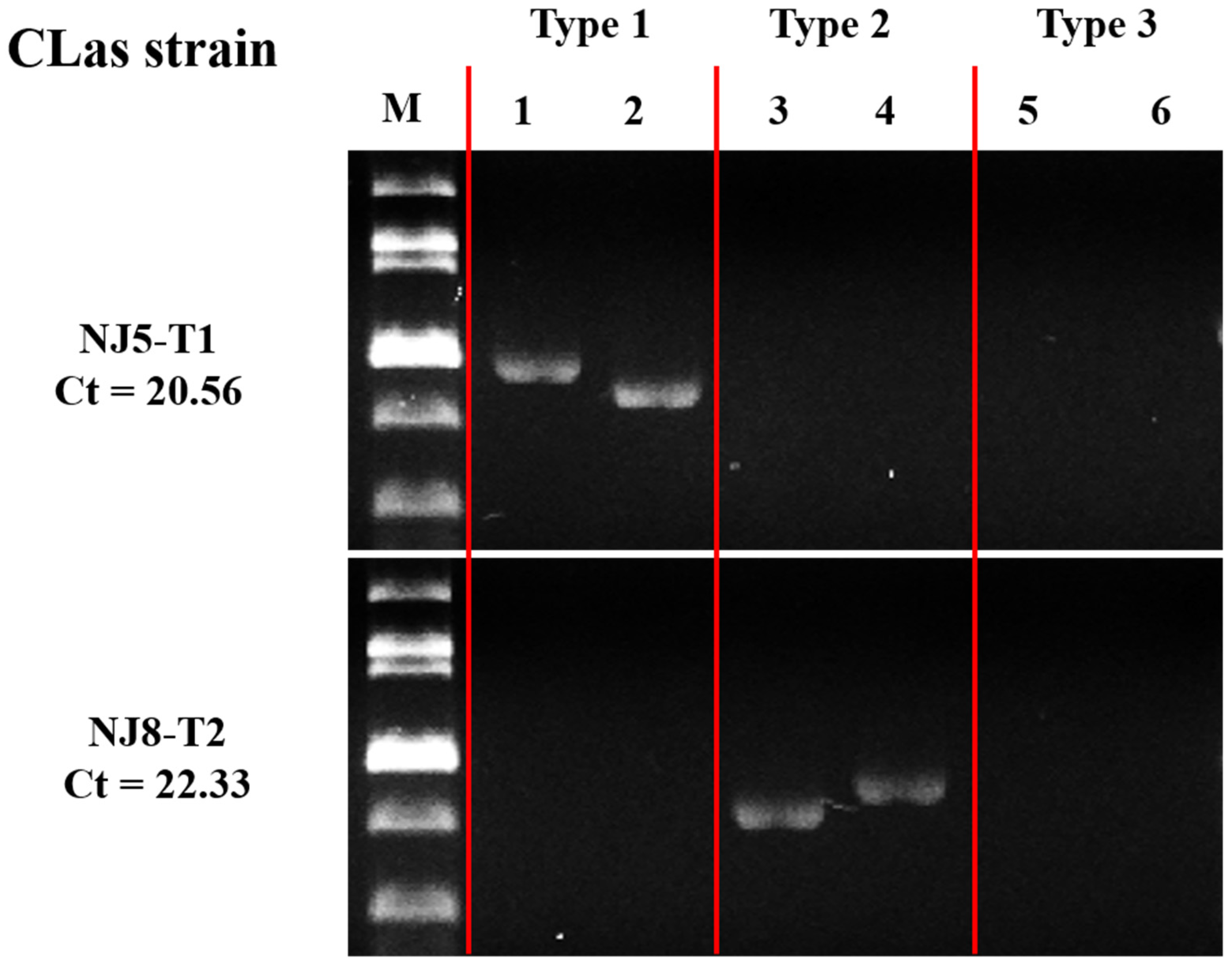
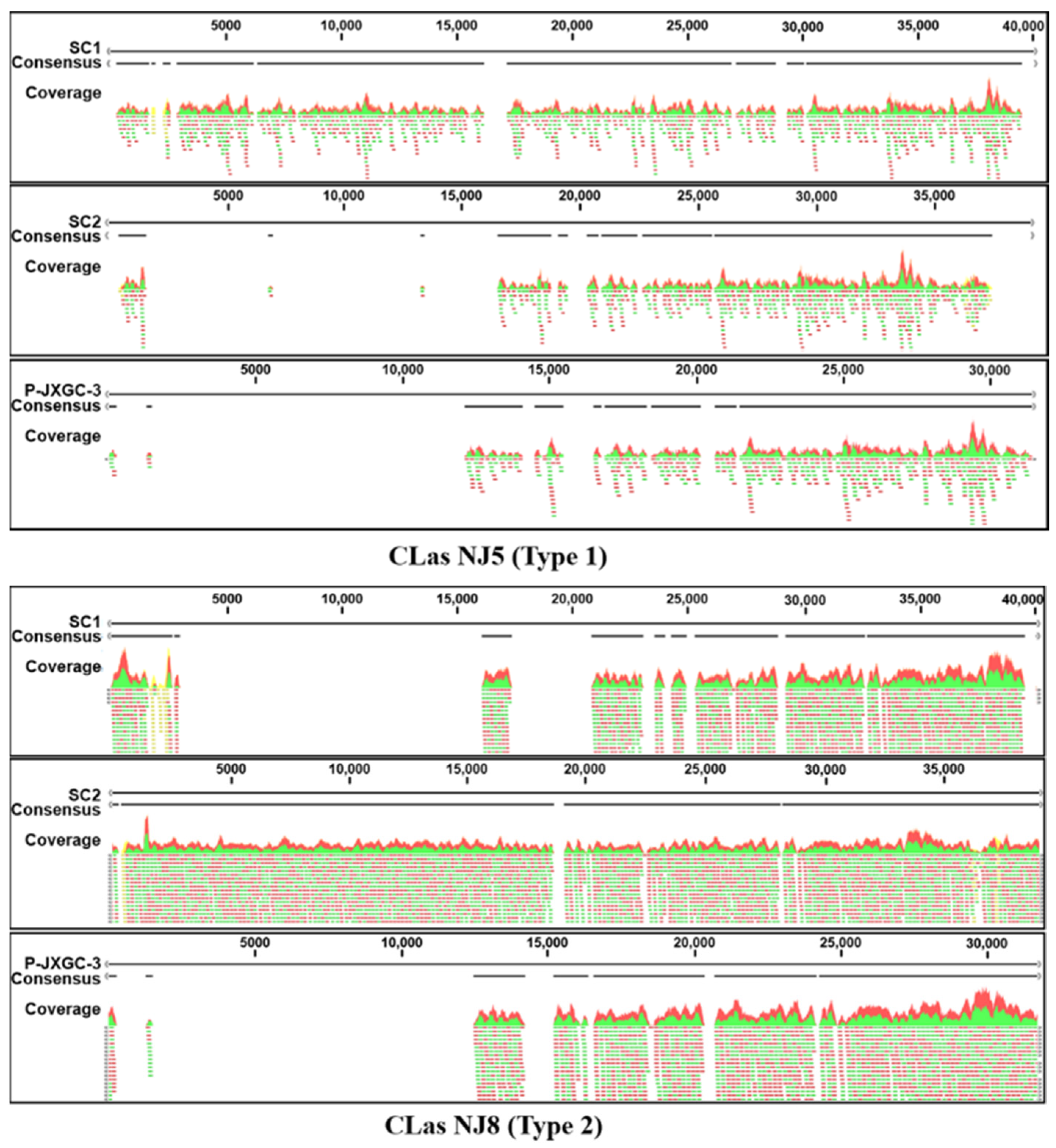
3. Results
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bové, J.M. Huanglongbing: A destructive, newly-emerging, century-old disease of citrus. J. Plant Pathol. 2006, 88, 7–37. [Google Scholar]
- Lin, K.H. Observations on yellow shoot of citrus. Acta Phytopathol. Sin. 1956, 2, 1–11. [Google Scholar]
- Duan, Y.; Zhou, L.; Hall, D.G.; Li, W.; Doddapaneni, H.; Lin, H.; Liu, L.; Vahling, C.M.; Gabriel, D.W.; Williams, K.P.; et al. Complete genome sequence of citrus huanglongbing bacterium, ‘Candidatus Liberibacter asiaticus’ obtained through metagenomics. Mol. Plant-Microbe Interact. 2009, 22, 1011–1020. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Z.; Deng, X.; Chen, J. Whole-genome sequence of ‘Candidatus Liberibacter asiaticus’ from Guangdong, China. Genome Announc. 2014, 2, e00273-14. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Z.; Deng, X.; Chen, J. Draft genome sequence of ‘Candidatus Liberibacter asiaticus’ from California. Genome Announc. 2014, 2, e00999-14. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Z.; Sun, X.; Deng, X.; Chen, J. Whole-Genome Sequence of “Candidatus Liberibacter asiaticus” from a Huanglongbing-Affected Citrus Tree in Central Florida. Genome Announc. 2015, 3, e00169-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Flores-Cruz, Z.; Zhou, L.; Kang, B.-H.; Fleites, L.A.; Gooch, M.D.; Wulff, N.A.; Davis, M.J.; Duan, Y.-P.; Gabriel, D.W. ‘Ca. Liberibacter asiaticus’ Carries an Excision Plasmid Prophage and a Chromosomally Integrated Prophage That Becomes Lytic in Plant Infections. Mol. Plant-Microbe Interact. 2011, 24, 458–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Z.; Bao, M.; Wu, F.; Van Horn, C.; Chen, J.; Deng, X. A Type 3 Prophage of ‘Candidatus Liberibacter asiaticus’ Carrying a Restriction-Modification System. Phytopathology 2018, 108, 454–461. [Google Scholar] [CrossRef] [Green Version]
- Boyd, E.; Brüssow, H. Common themes among bacteriophage-encoded virulence factors and diversity among the bacteriophages involved. Trends Microbiol. 2002, 10, 521–529. [Google Scholar] [CrossRef]
- Deng, X.; Lopes, S.; Wang, X.; Sun, X.; Jones, D.; Irey, M.; Civerolo, E.; Chen, J. Characterization of “Candidatus Liberibacter Asiaticus” Populations by Double-Locus Analyses. Curr. Microbiol. 2014, 69, 554–560. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.; Wu, F.; Zheng, Z.; Yokomi, R.; Kumagai, L.; Cai, W.; Rascoe, J.; Polek, M.; Chen, J.; Deng, X. Prophage Diversity of ‘Candidatus Liberibacter asiaticus’ Strains in California. Phytopathology 2019, 109, 551–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleites, L.A.; Jain, M.; Zhang, S.; Gabriel, D.W. “Candidatus Liberibacter asiaticus” Prophage Late Genes May Limit Host Range and Culturability. Appl. Environ. Microbiol. 2014, 80, 6023–6030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, M.; Fleites, L.A.; Gabriel, D.W. Prophage-Encoded Peroxidase in ‘Candidatus Liberibacter asiaticus’ Is a Secreted Effector That Suppresses Plant Defenses. Mol. Plant-Microbe Interact. 2015, 28, 1330–1337. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Chen, J.; Deng, X. Historical Perspectives, Management, and Current Research of Citrus HLB in Guangdong Province of China, Where the Disease has been Endemic for Over a Hundred Years. Phytopathology 2018, 108, 1224–1236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunta, M.; Zheng, Z.; Wu, F.; da Graca, J.V.; Park, J.-W.; Deng, X.; Chen, J. Draft Whole-Genome Sequence of “Candidatus Liberibacter asiaticus” Strain TX2351 Isolated from Asian Citrus Psyllids in Texas, USA. Genome Announc. 2017, 5, e00170-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Z.; Bao, M.; Wu, F.; Chen, J.; Deng, X. Predominance of Single Prophage Carrying a CRISPR/cas System in “Candidatus Liberibacter asiaticus” Strains in Southern China. PLoS ONE 2016, 11, e0146422. [Google Scholar] [CrossRef] [PubMed]
- Bao, M.; Zheng, Z.; Sun, X.; Chen, J.; Deng, X. Enhancing PCR Capacity to Detect ‘Candidatus Liberibacter asiaticus’ Utilizing Whole Genome Sequence Information. Plant Dis. 2020, 104, 527–532. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Wu, F.; Kumagai, L.B.; Polek, M.; Deng, X.; Chen, J. Two ‘Candidatus Liberibacter asiaticus’ Strains Recently Found in California Harbor Different Prophages. Phytopathology 2017, 107, 662–668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Deng, X.; Sun, X.; Jones, D.; Irey, M.; Civerolo, E. Guangdong and Florida Populations of ‘Candidatus Liberibacter asiaticus’ Distinguished by a Genomic Locus with Short Tandem Repeats. Phytopathology 2010, 100, 567–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5′-3′) | Size (bp) | Genomic Locus/Specificity | Reference | |
|---|---|---|---|---|---|
| CLas-4G | Forward | AGTCGAGCGCGTATGCGAAT | 78 | 16S rRNA gene/species | Bao et al. 2020 [17] |
| HLBr | Reverse | GCGTTATCCCGTAGAAAAAGGTAG | |||
| HLBp | Probe | FAM-AGACGGGTGAGTAACGCG-BHQ | |||
| T1-2F | Forward | TGGCTCGGGTTCAGGTAAAT | 975 | Endolysin/Type 1 phage | Zheng et al. 2018 [8] |
| T1-2R | Reverse | AAGGGCGACGCATGTATTTC | |||
| T1-3F | Forward | CTCACTGCGTCTTGATTCGG | 866 | Hypothetical protein gene/Type 1 phage | Same as above |
| T1-3R | Reverse | CGAACGAGCGGTATGTTTGT | |||
| T2-2F | Forward | ACCCTCGCACCATCATGTTA | 813 | Hypothetical protein gene/Type 1 phage | Same as above |
| T2-2R | Reverse | TCGTCTTGATTGGGCAGAGT | |||
| T2-3F | Forward | ACAGTTAAGAGCCACGGTGA | 918 | Hypothetical protein gene/Type 1 phage | Same as above |
| T2-3R | Reverse | AAGACGTGGGTGTTATGGGT | |||
| 891-1F | Forward | CTGATCCTTTACCATGCCGC | 950 | hsdS/Type 3 phage | Same as above |
| 891-1R | Reverse | CAGCGAAACCGATCTTGAGG | |||
| 891-2F | Forward | ACCGCGATCTACCCGTAATT | 884 | hsdR/Type 3 phage | Same as above |
| 891-2R | Reverse | TGTGTTTTGCGAGTGAAGGG |
| CLas Strain | Total Reads | SC1 (40,048 bp) | SC2 (38,997 bp) | P-JXGC-3 (31,449 bp) |
|---|---|---|---|---|
| NJ5-T1: | ||||
| Total (reads) | 74,443,296 | 1130 | 641 | 550 |
| Consensus (bp) | 36,326 | 20,678 | 17,277 | |
| % coverage to reference | 90.7 | 53.0 | 54.9 | |
| NJ8-T2: | ||||
| Total (reads) | 85,867,742 | 4491 | 8273 | 3835 |
| Consensus (bp) | 21,392 | 38,343 | 17,937 | |
| % coverage to reference | 53.5 | 98.3 | 57.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bao, M.; Zheng, Z.; Chen, J.; Deng, X. Investigation of Citrus HLB Symptom Variations Associated with “Candidatus Liberibacter asiaticus” Strains Harboring Different Phages in Southern China. Agronomy 2021, 11, 2262. https://doi.org/10.3390/agronomy11112262
Bao M, Zheng Z, Chen J, Deng X. Investigation of Citrus HLB Symptom Variations Associated with “Candidatus Liberibacter asiaticus” Strains Harboring Different Phages in Southern China. Agronomy. 2021; 11(11):2262. https://doi.org/10.3390/agronomy11112262
Chicago/Turabian StyleBao, Minli, Zheng Zheng, Jianchi Chen, and Xiaoling Deng. 2021. "Investigation of Citrus HLB Symptom Variations Associated with “Candidatus Liberibacter asiaticus” Strains Harboring Different Phages in Southern China" Agronomy 11, no. 11: 2262. https://doi.org/10.3390/agronomy11112262