Contribution of Awns to Seed Yield and Seed Shattering in Siberian Wildrye Grown under Irrigated and Rainfed Environments


Abstract
:1. Introduction
2. Materials and Methods
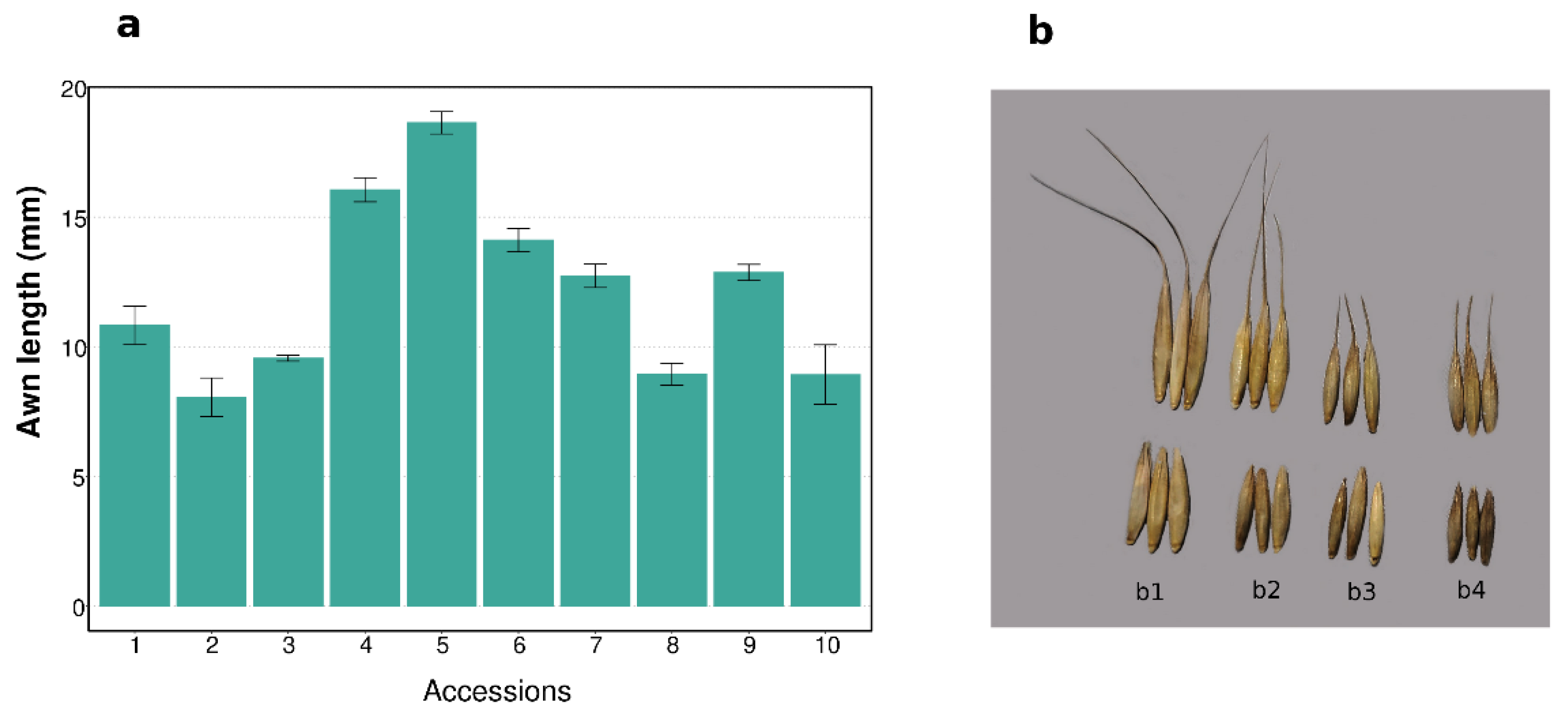
2.1. Plant Materials
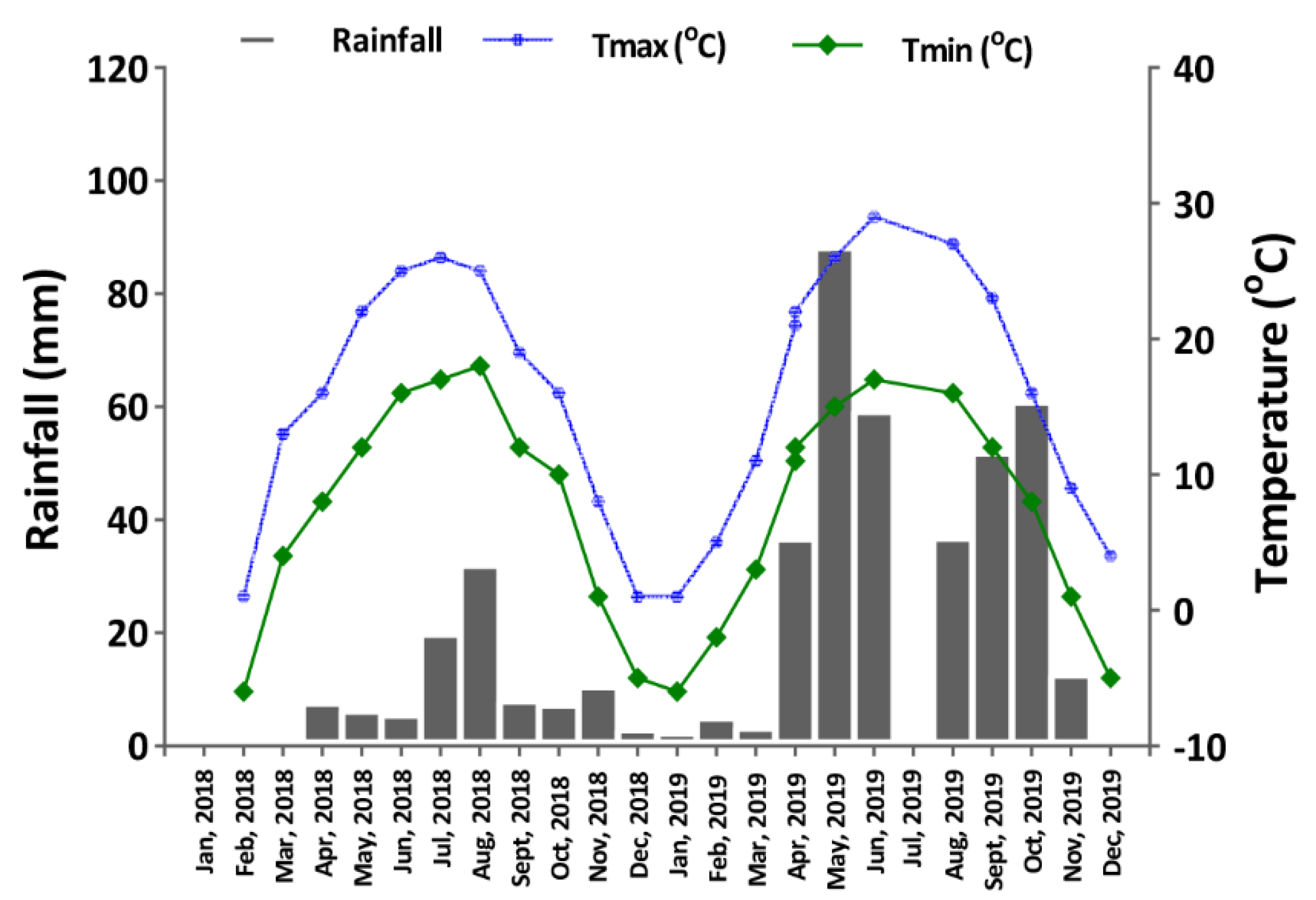
2.2. Experimental Conditions and Trial Design
2.3. Data Collection and Measurements
2.3.1. Awn Length and Seed Yield Components
2.3.2. Spike Dry Matter
2.3.3. Seed Yield
2.3.4. Seed Shattering
Breaking Tensile Strength (BTS)
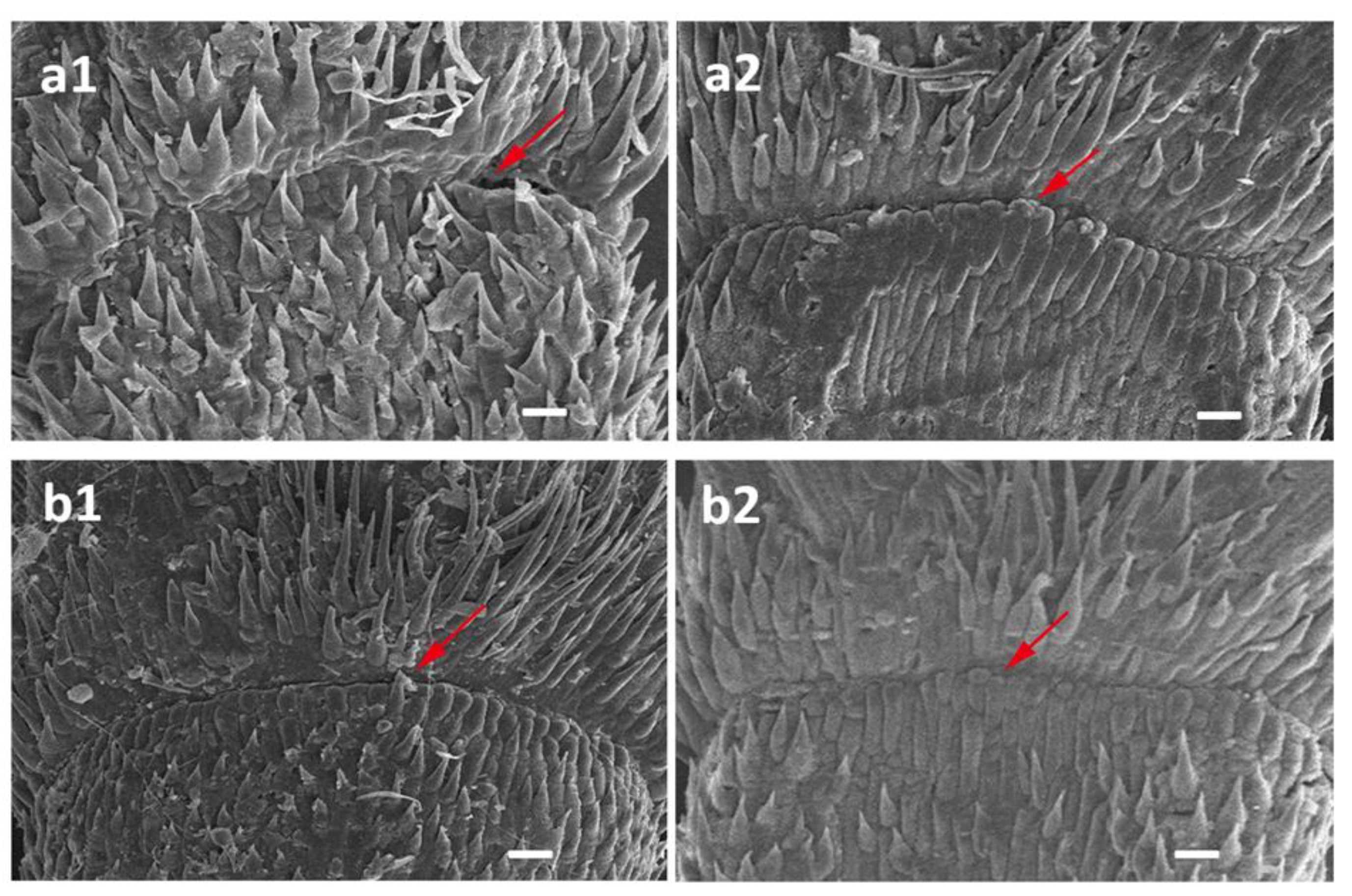
Scanning Electron Microscope (SEM)
2.4. Statistical Analysis
3. Results
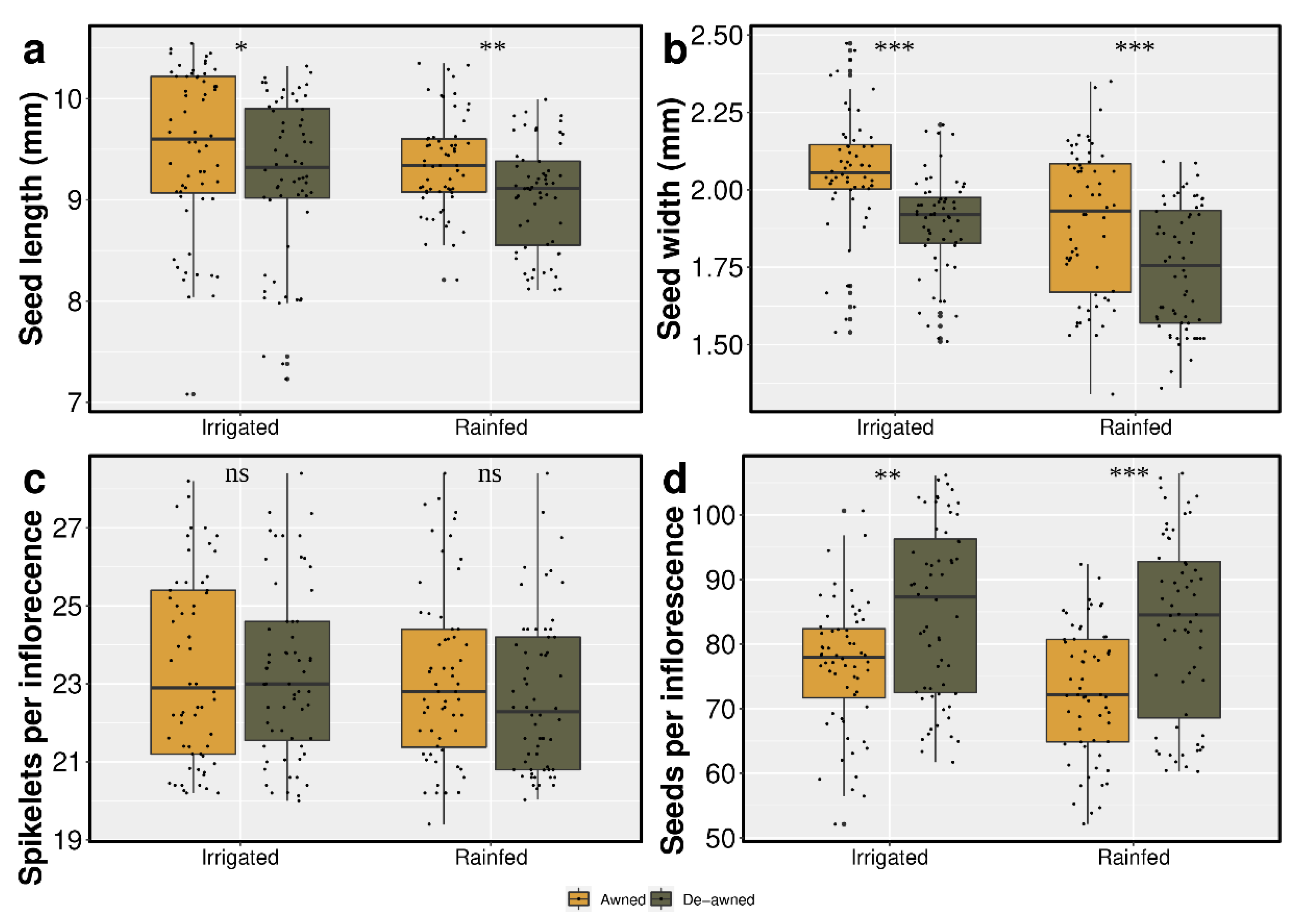
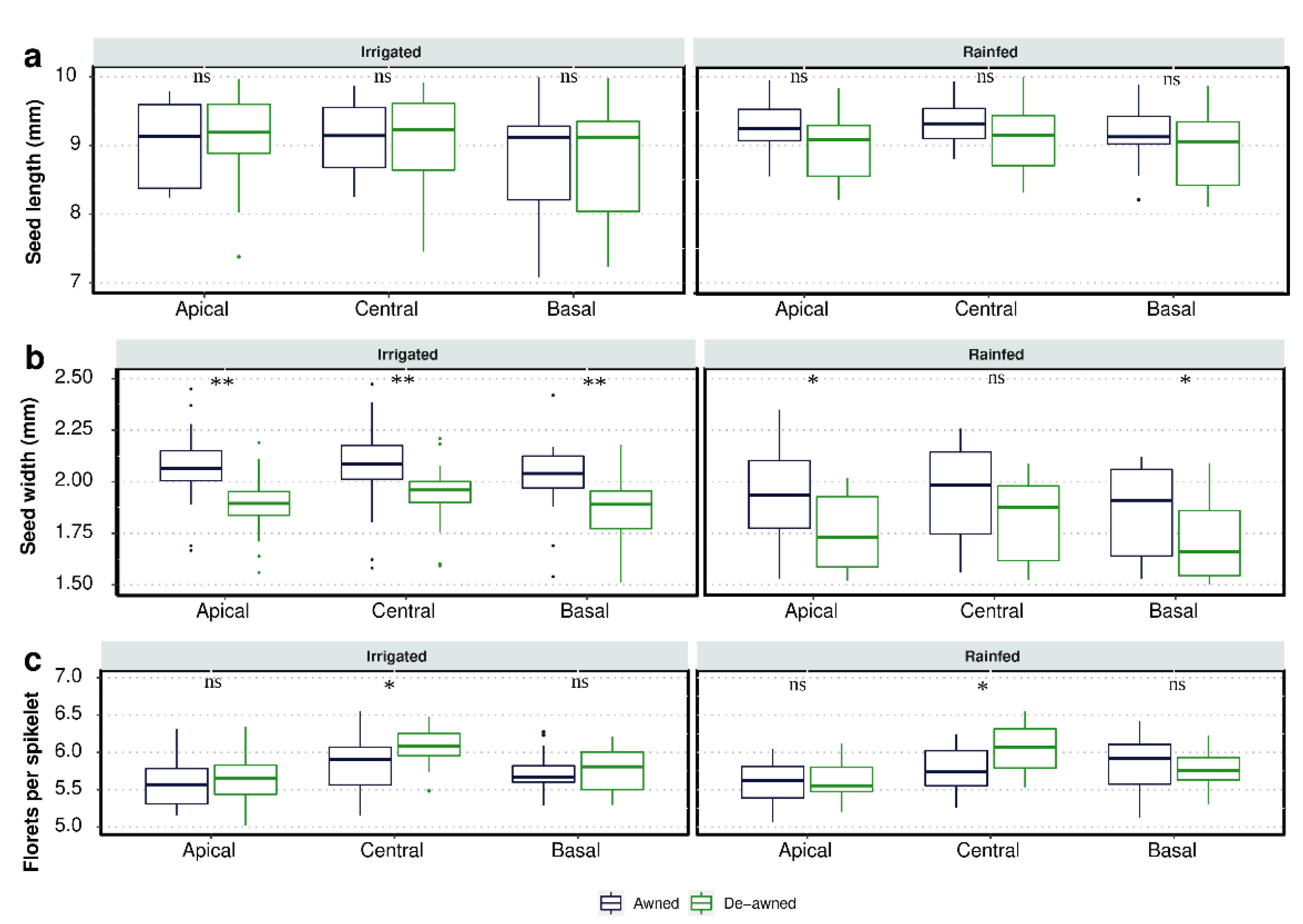
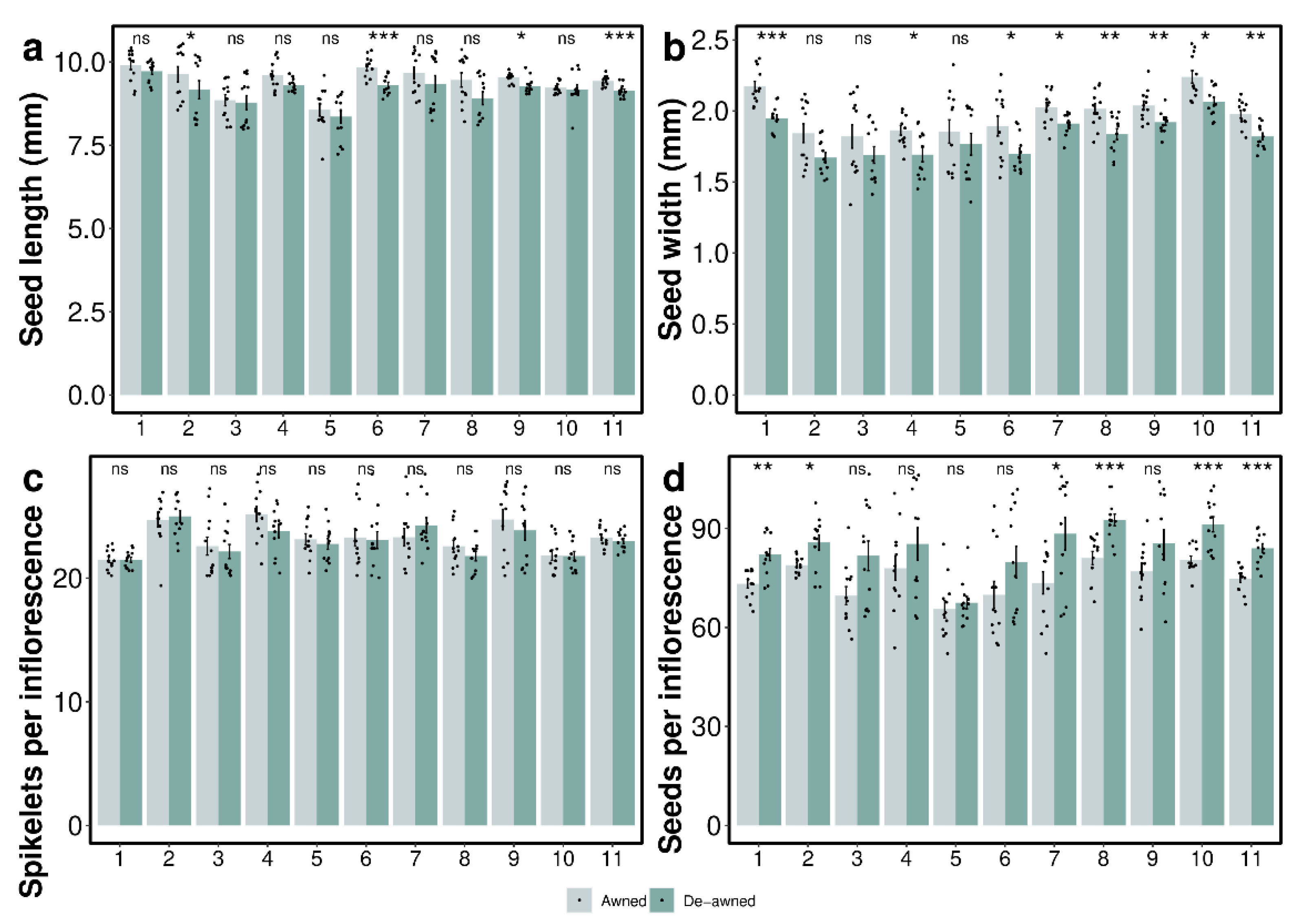
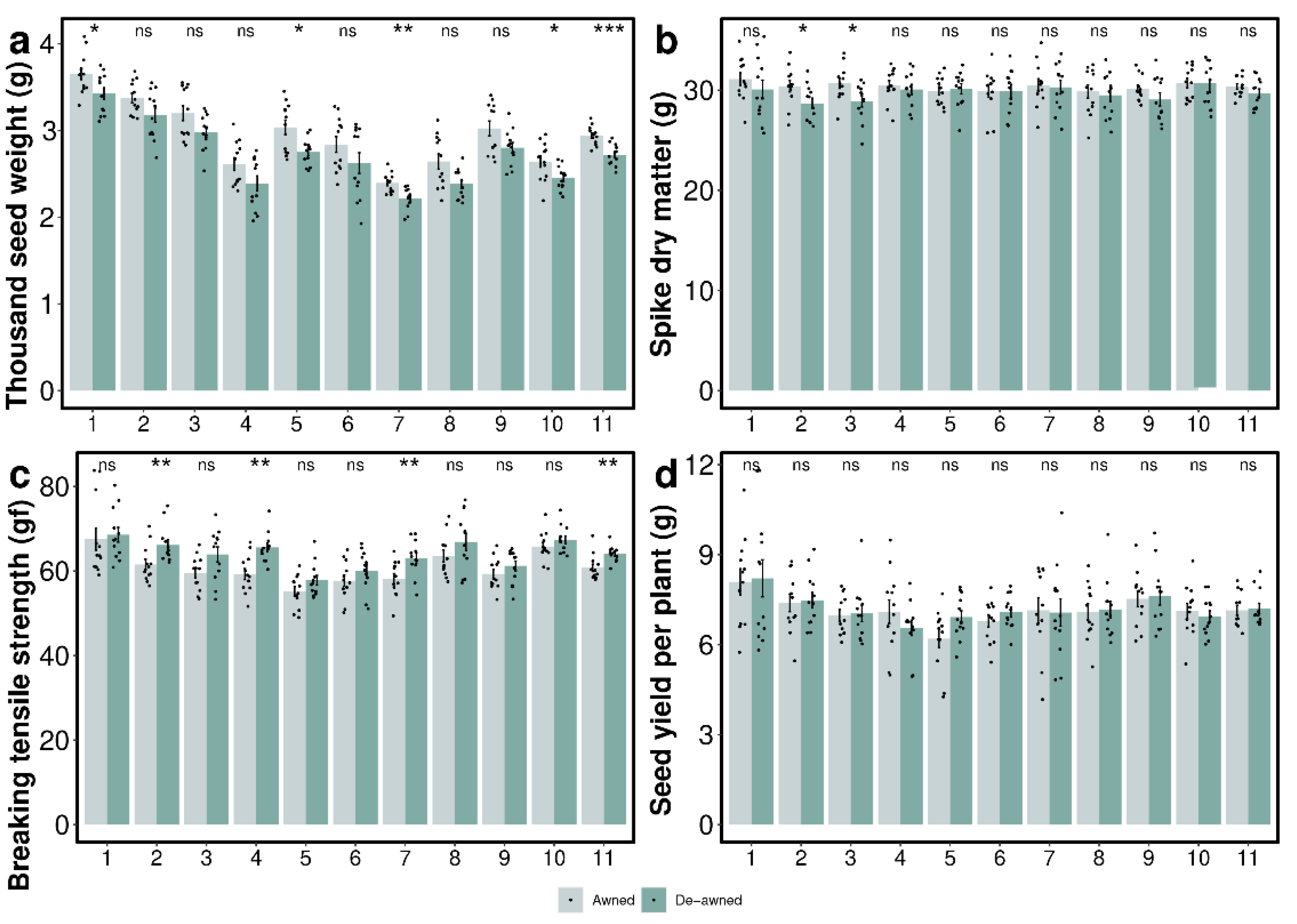
3.1. Awn Length and Seed Yield Components
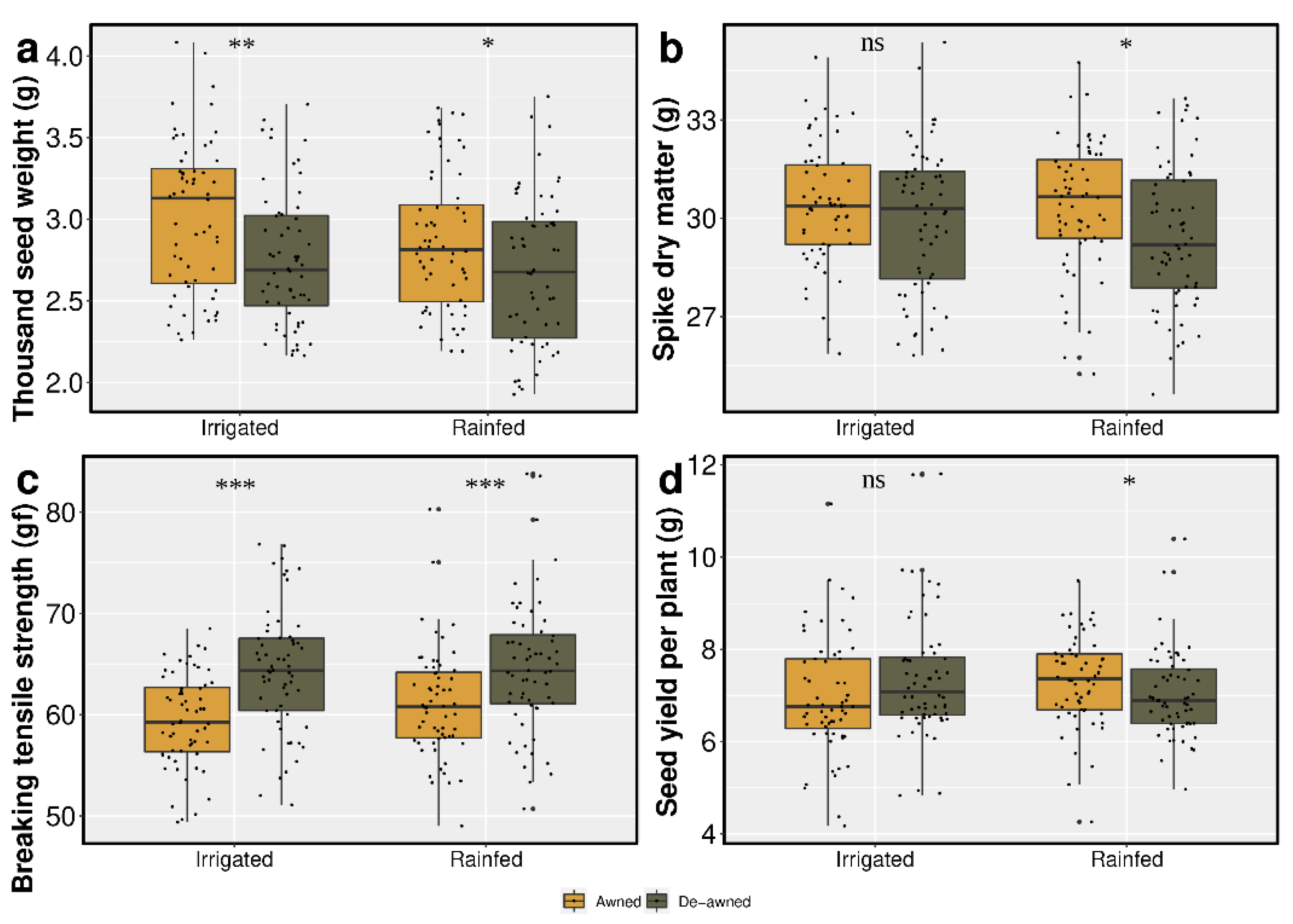
3.2. Spike Dry Matter Accumulation
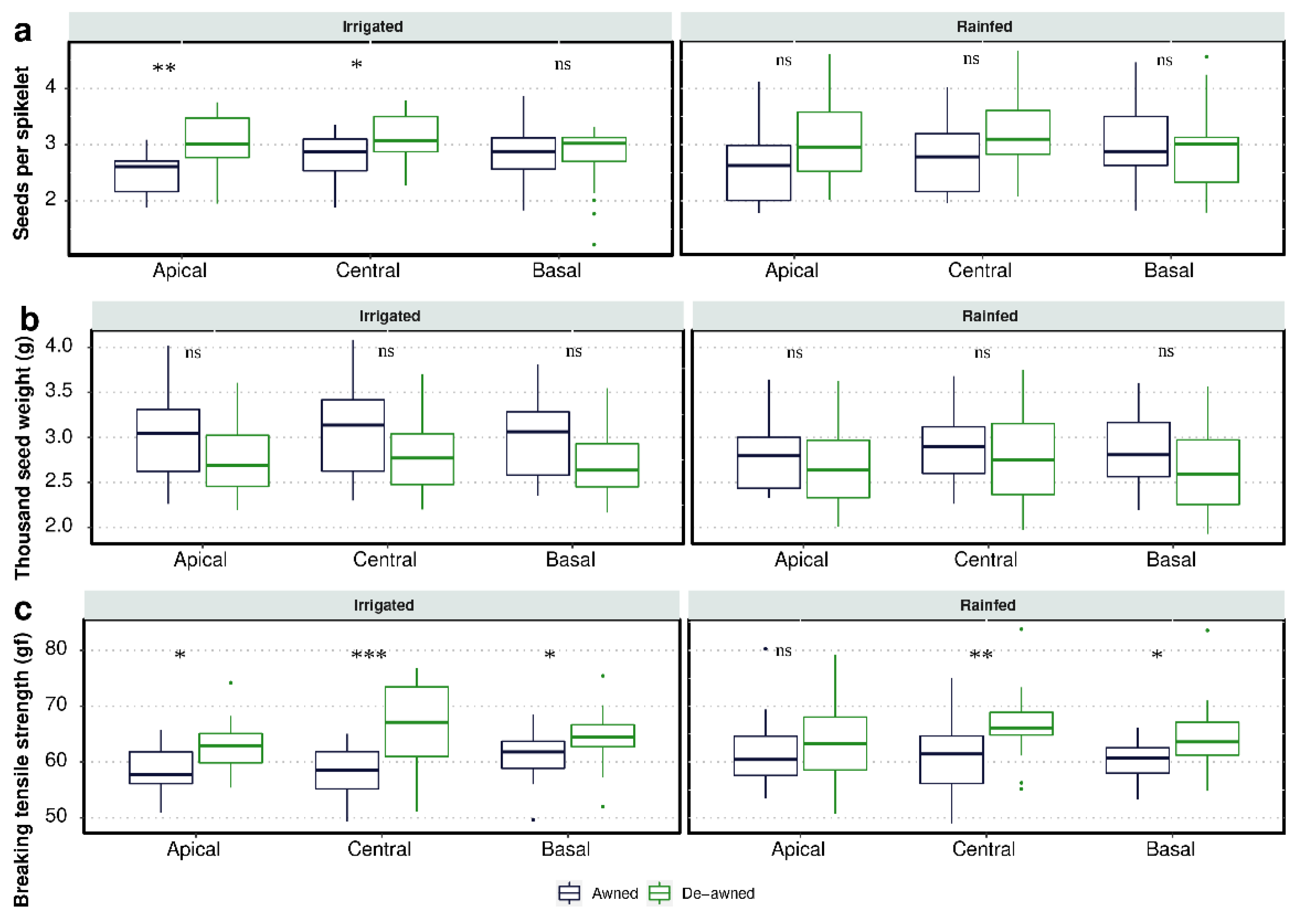
3.3. Seed Yield
3.4. Effect of Awn Excision on Seed Shattering
4. Discussion
4.1. Impacts of Awns on Seed Yield and Components
4.2. Role of Awns on Partitioning of Spike Dry Matter
4.3. The Impact of Awns on Seed Shattering
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, Z.; Xie, W.; Zhao, Y.; Zhang, J.; Wang, Y. EST-SSR marker development based on RNA-sequencing of E. sibiricus and its application for phylogenetic relationships analysis of seventeen Elymus species. BMC Plant Biol. 2019, 19, 235. [Google Scholar] [CrossRef]
- Barkworth, M.E. Flora of North America: North of Mexico: Magnoliophyta: Commelinidae (In Part): Poaceae, Part 1. 2006, Volume 24. Available online: http://floranorthamerica.org/volumes (accessed on 20 July 2020).
- Xie, W.; Zhang, J.; Zhao, X.; Zhang, Z.; Wang, Y. Transcriptome profiling of Elymus sibiricus, an important forage grass in Qinghai-Tibet plateau, reveals novel insights into candidate genes that potentially connected to seed shattering. BMC Plant Biol. 2017, 17, 78. [Google Scholar] [CrossRef] [PubMed]
- Ntakirutimana, F.; Xiao, B.; Xie, W.; Zhang, J.; Zhang, Z.; Wang, N.; Yan, J. Potential effects of awn length variation on seed yield and components, seed dispersal and germination performance in Siberian wildrye (Elymus sibiricus L.). Plants 2019, 8, 561. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Hou, L.; Zhu, Y.; Zhang, Q.; Wang, H.; Xia, F.; Chen, L.; Mao, P.; Hannaway, D.B. Siberian wildrye seed yield limited by assimilate source. Field Crop Res. 2018, 218, 18–23. [Google Scholar] [CrossRef]
- Capstaff, N.M.; Miller, A.J. Improving the Yield and Nutritional Quality of Forage Crops. Front. Plant Sci. 2018, 9, 535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Z.; Schnurbusch, T. Costs and benefits of awns. J. Exp. Bot. 2016, 67, 2533–2535. [Google Scholar] [CrossRef] [Green Version]
- Rebetzke, G.J.; Bonnett, D.G.; Reynolds, M.P. Awns reduce grain number to increase grain size and harvestable yield in irrigated and rainfed spring wheat. J. Exp. Bot. 2016, 67, 2573–2586. [Google Scholar] [CrossRef] [PubMed]
- Merah, O.; Monneveux, P. Contribution of different organs to grain filling in durum wheat under Mediterranean conditions I. Contribution of post-anthesis photosynthesis and remobilization. J. Agron. Crop Sci. 2015, 201, 344–352. [Google Scholar] [CrossRef] [Green Version]
- Gu, X.Y.; Kianian, S.F.; Foley, M.E. Phenotypic selection for dormancy introduced a set of adaptive haplotypes from weedy into cultivated rice. Genetics 2005, 171, 695–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bommert, P.; Whipple, C. Grass Inflorescence Architecture and Meristem Determinacy. Semin. Cell Dev. Biol. 2017, 79, 6429. [Google Scholar] [CrossRef]
- Elbaum, R.; Zaltzman, L.; Burgert, I.; Fratzl, P. The role of wheat awns in the seed dispersal unit. Science 2007, 316, 884–886. [Google Scholar] [CrossRef] [PubMed]
- Yuo, T.; Yamashita, Y.; Kanamori, H.; Matsumoto, T.; Lundqvist, U.; Sato, K.; Ichii, M.; Jobling, S.A.; Taketa, S. A SHORT INTERNODES (SHI) family transcription factor gene regulates awn elongation and pistil morphology in barley. J. Exp. Bot. 2012, 63, 5223–5232. [Google Scholar] [CrossRef] [Green Version]
- Liller, C.; Walla, A.; Boer, M.P.; Hedley, P.; Macaulay, M.; Effgen, S.; Korff, M.V.; Esse, G.V.; Koornneef, M. Fine mapping of a major QTL for awn length in barley using a multiparent mapping population. Theor. Appl. Genet. 2017, 130, 269–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ntakirutimana, F.; Xie, W. Unveiling the actual functions of awns in grasses: From yield potential to quality traits. Int. J. Mol. Sci. 2020, 21, 7593. [Google Scholar] [CrossRef] [PubMed]
- Motzo, R.; Giunta, F. Awnedness affects grain yield and kernel weight in near-isogenic lines of durum wheat. Aust. J. Agric. Res. 2002, 53, 1285–1293. [Google Scholar] [CrossRef]
- Blum, A. Photosynthesis and transpiration in leaves and ears of wheat and barley varieties. J. Exp. Bot. 1985, 36, 432–440. [Google Scholar] [CrossRef]
- Maydup, M.L.; Antonietta, M.; Graciano, C.; Guiamet, J.J.; Tambussi, E.A. The contribution of the awns of bread wheat (Triticum aestivum L.) to grain filling: Responses to water deficit and the effects of awns on ear temperature and hydraulic conductance. Field Crops Res. 2014, 167, 102–111. [Google Scholar] [CrossRef]
- Furuta, T.; Komeda, N.; Asano, K.; Uehara, K.; Gamuyao, R.; Angeles-Shim, R.B.; Nagai, K.; Doi, K.; Wang, D.R.; Yasui, H. Convergent loss of awn in two cultivated rice species Oryza sativa and Oryza glaberrima is caused by mutations in different loci. G3 Genesgenetics 2015, 5, 2267–2274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, F.X.; Kim, D.M.; Ju, H.G.; Ahn, S.N. Mapping quantitative trait loci for awnness and yield component traits in isogenic lines derived from an Oryza sativa/O. rufipogon cross. J. Crop Sci. Biotechnol. 2009, 12, 9–15. [Google Scholar] [CrossRef]
- Singh, S.; Singh, T.N.; Chauhan, J.S. Architectural engineering of rice panicle for increased productivity: A powerful Biological Tool for Combating Agricultural Water Crisis. J. Crop Improv. 2009, 23, 451–466. [Google Scholar] [CrossRef]
- Zhao, X.; Xie, W.; Zhang, J.; Zhang, Z.; Wang, Y. Histological Characteristics, Cell Wall hydrolytic enzymes activity and candidate genes expression associated with seed shattering of Elymus sibiricus accessions. Front. Plant Sci. 2017, 8, 606. [Google Scholar] [CrossRef] [Green Version]
- Konopatskaia, I.; Vavilova, V.; Blinov, A.; Goncharov, N.P. Spike morphology genes in wheat species (Triticum L.). Proc. Latv. Acad. Sci. 2016, 70, 345–355. [Google Scholar] [CrossRef] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Duwayri, M. Effect of flag leaf and awn removal on grain yield and yield components of wheat grown under dryland conditions. Field Crops Res. 1984, 8, 307–313. [Google Scholar] [CrossRef]
- Huo, X.; Wu, S.; Zhu, Z.; Liu, F.; Fu, Y.; Cai, H.; Sun, X.; Gu, P.; Xie, D.; Tan, L. NOG1 increases grain production in rice. Nat. Commun. 2017, 8, 1497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chhabra, A.K.; Sethi, S.K. Inheritance of cleistogamic flowering in durum wheat (Triticum durum). Euphytica 1991, 55, 147–150. [Google Scholar] [CrossRef]
- Khaliq, I.; Irshad, A.; Ahsan, M. Awns and flag leaf contribution towards grain yield in spring wheat (Triticum aestivum L.). Cereal Res. Commun. 2008, 36, 65–76. [Google Scholar] [CrossRef]
- Chen, G.D.; Li, H.B.; Zheng, Z.; Wei, Y.M.; Zheng, Y.L.; McIntyre, C.L.; Zhou, M.X.; Liu, C.J. Characterization of a QTL affecting spike morphology on the long arm of chromosome 3H in barley (Hordeum vulgare L.) based on near isogenic lines and a NIL-derived population. Theor. Appl. Genet. 2012, 125, 1385–1392. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.Z.; Roche, D.; Durham, S.; Hole, D. Awn contribution to gas exchanges of barley ears. Photosynthetica 2006, 44, 536–541. [Google Scholar] [CrossRef]
- Grundbacher, F.J. The physiological function of the cereal awn. Bot. Rev. 1963, 29, 366–381. [Google Scholar] [CrossRef]
- Teich, A.H. Interaction of awns and environment on grain yield in winter wheat (Triticum aestivum L.). Cereal Res. Commun. 1982, 10, 1–15. Available online: https://www.jstor.org/stable/23781379 (accessed on 15 May 2021).
- Bort, J.; Febrero, A.; Amaro, T.; Araus, J. Role of awns in ear water-use efficiency and grain weight in barley. Agronomie 1994, 14, 133–139. [Google Scholar] [CrossRef]
- Wang, Q.; Zhang, T.; Cui, J.; Wang, X.; He, Z.; Han, J.; René, G.; Fuller, D.Q. Path and ridge regression analysis of seed yield and seed yield components of Russian Wildrye (Psathyrostachys juncea Nevski) under Field Conditions. PLoS ONE 2011, 6, e18245. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Li, Z.Z. Seasonal characteristics of soil water and biomass of siberian wildrye grass (Elymus sibiricus L.) in Bashang Plateau. Agric. Res. Arid Areas 2009, 27, 90–95. [Google Scholar]
- Keller, M.; Koblet, W. Dry matter and leaf area partitioning, bud fertility and second season growth of Vitis vinifera L.: Responses to nitrogen supply and limiting irradiance. Vitis 1995, 34, 77–83. [Google Scholar]
- Sulpice, R.; Nikoloski, Z.; Tschoep, H.; Antonio, C.; Kleessen, S.; Larhlimi, A.; Selbig, J.; Ishihara, H.; Gibon, Y.; Stitt, F.M. Impact of the carbon and nitrogen supply on relationships and connectivity between metabolism and biomass in a broad panel of Arabidopsis accessions. Plant Physiol. 2013, 162, 347–363. [Google Scholar] [CrossRef] [Green Version]
- González, F.; Miralles, D.J.; Slafer, G.A. Wheat floret survival as related to pre-anthesis spike growth. J. Exp. Bot. 2011, 62, 4889–4901. [Google Scholar] [CrossRef] [PubMed]
- Martinez, D.E.; Luquez, V.M.; Bartoli, C.G.; Guiamét, J. Persistence of photosynthetic components and photochemical efficiency in ears of water-stressed wheat (Triticum aestivum). Physiol. Plant. 2003, 119, 519–525. [Google Scholar] [CrossRef]
- Yang, D.; Wang, Y.Z. Seed shattering: From models to crops. Front. Plant Sci. 2015, 6, 476. [Google Scholar] [CrossRef]
- Ji, H.; Kim, S.; Kim, Y.; Kim, H.; Eun, M.; Jin, I.; Cha, Y.; Yun, D.; Ahn, B.; Lee, M. Inactivation of the CTD phosphatase-like gene OsCPL1 enhances the development of the abscission layer and seed shattering in rice. Plant J. 2010, 61, 96–106. [Google Scholar] [CrossRef]
- Li, W.; Gill, B.S. Multiple genetic pathways for seed shattering in the grasses. Funct. Integr. Genom. 2006, 6, 300–309. [Google Scholar] [CrossRef] [PubMed]
- Thomson, M.J.; Tai, T.H.; McClung, A.M.; Lai, X.H.; Hinga, M.E.; Lobos, K.B.; Xu, Y.; Martinez, C.P.; McCouch, S.R. Mapping quantitative trait loci for yield, yield components and morphological traits in an advanced backcross population between Oryza rufipogonand and the Oryza sativa cultivar Jefferson. Theor. Appl. Genet. 2003, 107, 479–493. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Lu, D.; Li, C.; Luo, J.; Zhu, B.F.; Zhu, J.; Shangguan, Y.; Wang, Z.; Sang, T.; Zhou, B. Genetic control of seed shattering in rice by the APETALA2 transcription factor SHATTERING ABORTION1. Plant Cell 2012, 24, 1034–1048. [Google Scholar] [CrossRef] [Green Version]
- Magwa, R.A.; Zhao, H.U.; Wen, Y.; Xie, W.; Lin, Y. Genomewide association analysis for awn length linked to the seed shattering gene qSH1 in rice. J. Genet. 2016, 95, 639. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Running Code | Accessions | Status | Origin |
|---|---|---|---|
| 1 | PI499456 | Wild | Inner Mongolia, China |
| 2 | LT02 | Wild | Gansu, China |
| 3 | PI499615 | Wild | Xinjiang, China |
| 4 | PI531665 | Wild | Beijing, China |
| 5 | ZN03 | Wild | Gansu, China |
| 6 | PI598479 | Wild | Altai, Russia |
| 7 | PI598788 | Wild | Alma Ata, Kazakhstan |
| 8 | PI655097 | Wild | Alma Ata, Kazakhstan |
| 9 | PI659942 | Wild | Alma Ata, Kazakhstan |
| 10 | HZ03 | Wild | Gansu, China |
| Source of Variation | df | SL | SW | TKW | BTS | SpI | SI | SDM | SYP |
|---|---|---|---|---|---|---|---|---|---|
| Seasons (S) | 1 | ns | ns | ** | ns | *** | ** | ** | ns |
| Blocks | 2 | ns | ns | ns | ns | ns | ns | ns | ns |
| Awn (A) | 1 | *** | *** | *** | *** | ns | *** | ** | ns |
| Environment (E) | 1 | * | *** | * | ns | ns | * | * | * |
| S x A | 1 | ns | ns | ns | ns | ns | * | * | ns |
| S x E | 1 | ns | ns | ns | ns | ns | ns | * | ns |
| A x E | 1 | ns | ns | ns | ns | ns | * | * | * |
| A x E x S | 1 | ns | ns | ns | ns | ns | ns | ns | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ntakirutimana, F.; Wan, Y.; Liu, W.; Xie, W. Contribution of Awns to Seed Yield and Seed Shattering in Siberian Wildrye Grown under Irrigated and Rainfed Environments. Agronomy 2021, 11, 2219. https://doi.org/10.3390/agronomy11112219
Ntakirutimana F, Wan Y, Liu W, Xie W. Contribution of Awns to Seed Yield and Seed Shattering in Siberian Wildrye Grown under Irrigated and Rainfed Environments. Agronomy. 2021; 11(11):2219. https://doi.org/10.3390/agronomy11112219
Chicago/Turabian StyleNtakirutimana, Fabrice, Yiyang Wan, Wenhui Liu, and Wengang Xie. 2021. "Contribution of Awns to Seed Yield and Seed Shattering in Siberian Wildrye Grown under Irrigated and Rainfed Environments" Agronomy 11, no. 11: 2219. https://doi.org/10.3390/agronomy11112219