
The Shape Diversity of Olive Stones Resulting from Domestication and Diversification Unveils Traits of the Oldest Known 6500-Years-Old Table Olives from Hishuley Carmel Site (Israel)

 , , , , , and
, , , , , and 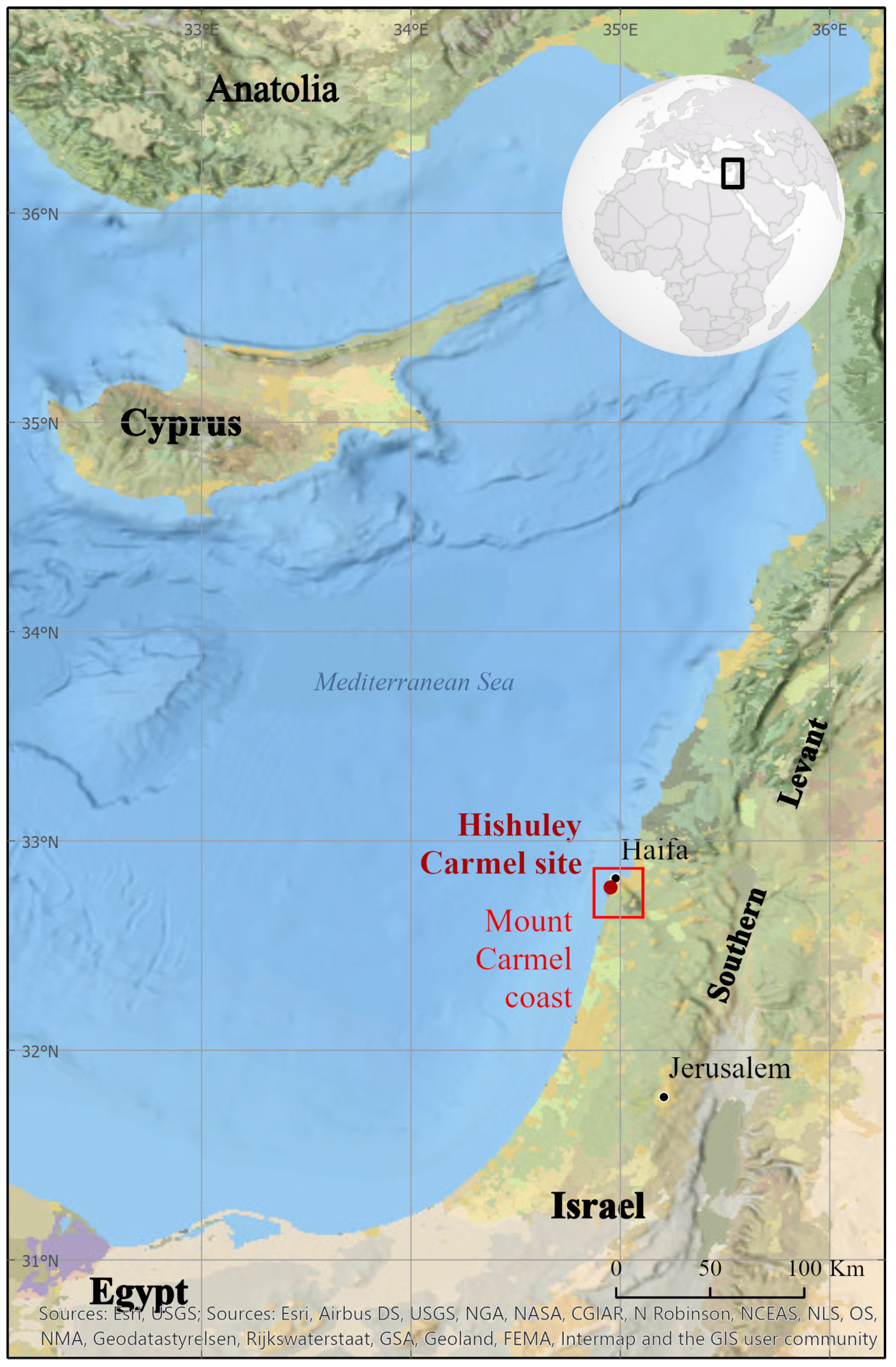{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
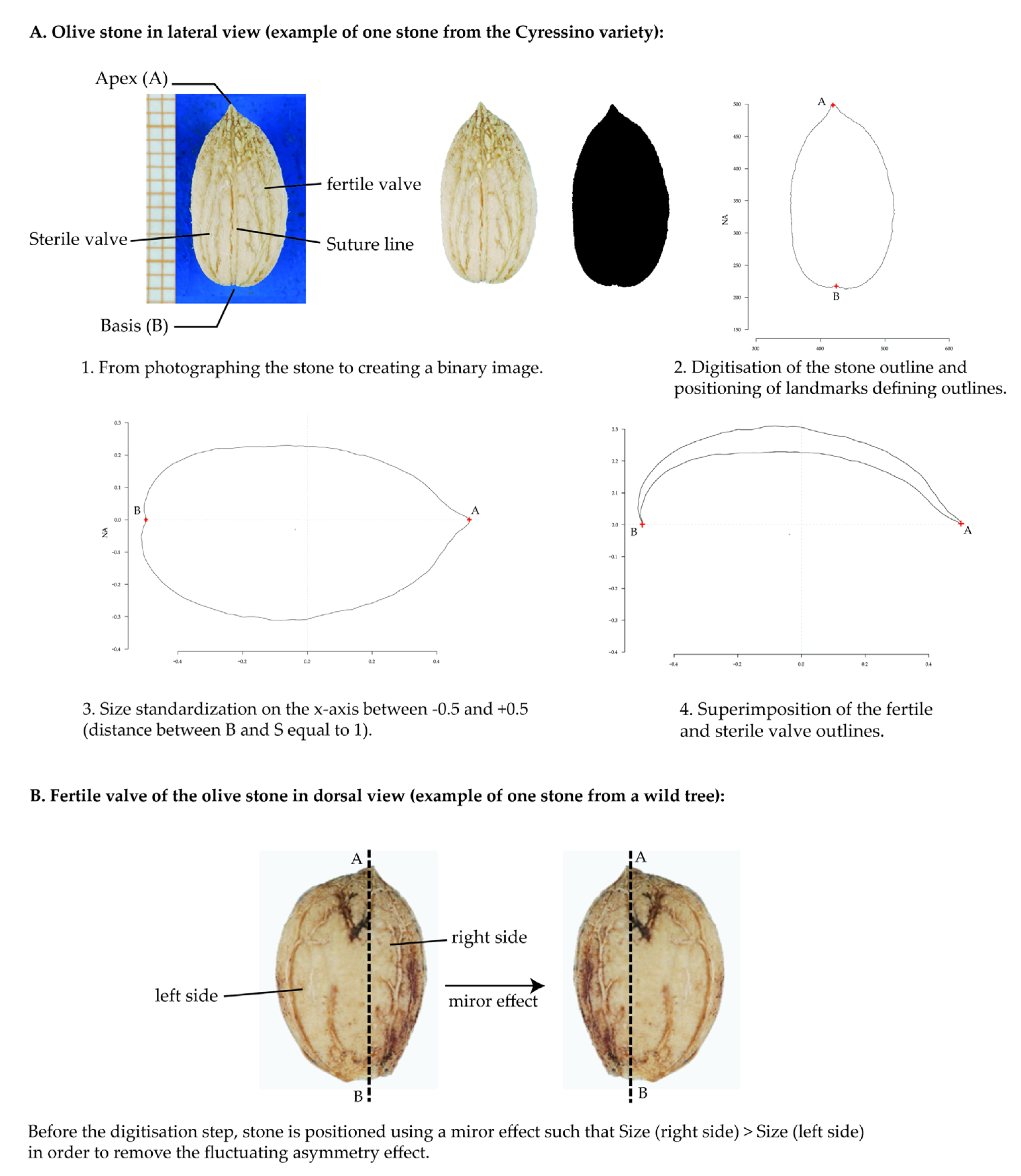
2.2. Stone Shape Analysis
2.3. Statistical Analyses
3. Results
3.1. Number of Landmarks Used to Define the Stone Outline
3.2. Morphological Differentiation between Current Wild Populations and Varieties
3.3. Morphological Variability of the Two Sets of Archaeological Stones (A and B) from Two Distinct Structures in the Hishuley Carmel Site
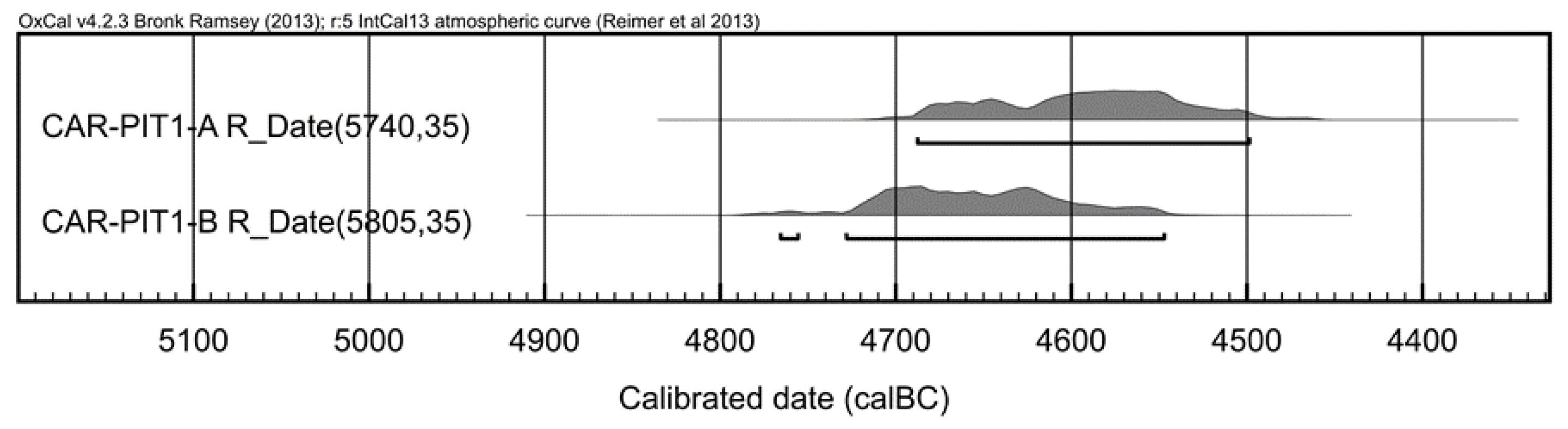
3.4. Dating of Archaeological Olive Stones
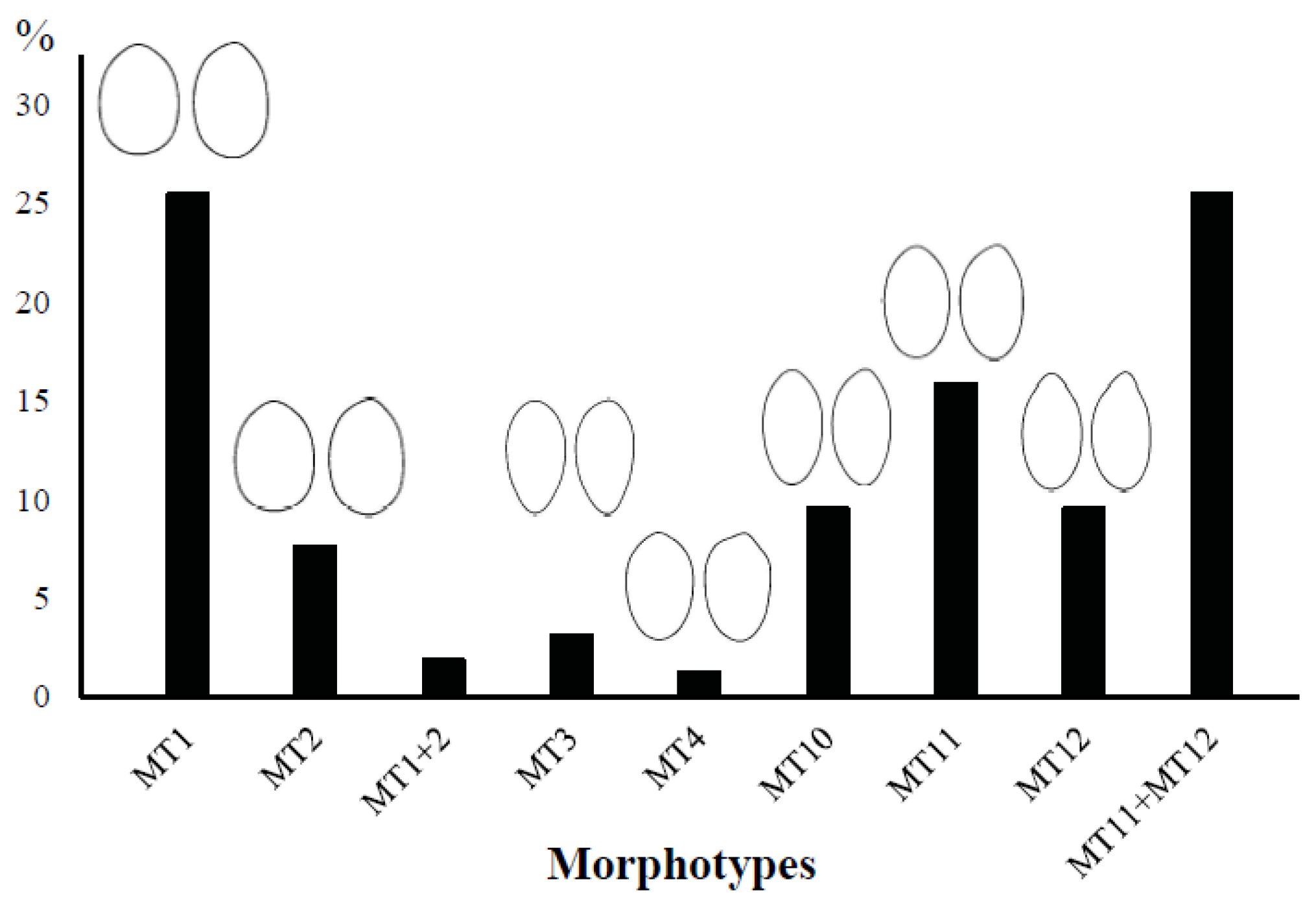
3.5. Identification of Morphotypes in the Archaeological Material
- MT1 (40 stones—25.5% of the classified archaeological material), consisting of stones from supposed wild populations and one variety with a ‘primitive’ morphology (Arbequina) differing from wild forms by its larger size, although this trait was not considered in this study. Arbequina was distinguished from wild populations with a rate of 86.7%;
- MT2 (12 stones—7.6%) defined by stones of varieties (all for table or mixed use) from the Eastern Mediterranean and one French variety. All varieties were allocated to the Eastern gene pool;
- MT10 (15 stones—9.6%), composed of varieties of the Eastern maternal lineage, which were relatively different in terms of geographical origin (cultivation area and nuclear genetic data) and use of the fruit;
- MT11 (25 stones—15.9%) composed of supposed wild populations and varieties with diverse origins, uses and maternal lineages;
- MT12 (15 stones—9.6%) including mainly oriental varieties used primarily for oil or mixed use.
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Suc, J.-P. Origin and evolution of the Mediterranean vegetation and climate in Europe. Nat. Cell Biol. 1984, 307, 429–432. [Google Scholar] [CrossRef]
- Palamarev, E. Paleobotanical evidences of the Tertiary history and origin of the Mediterranean sclerophyll dendroflora. Plant Syst. Evol. 1989, 162, 93–107. [Google Scholar] [CrossRef]
- Terral, J.-F.; Badal, E.; Heinz, C.; Roiron, P.; Thiebault, S.; Figueiral, I. A hydraulic conductivity model points to post-neogene survival of the mediterranean olive. Ecology 2004, 85, 3158–3165. [Google Scholar] [CrossRef]
- Green, P.S. A Revision of Olea L. (Oleaceae). Kew Bull. 2002, 57, 91. [Google Scholar] [CrossRef]
- Martinez, S.R. Memoria del Mapa de Series de Vegetacion de España. 1: 400000; Icona: Madrid, Spain, 1987. [Google Scholar]
- Barazani, O.; Keren-Keiserman, A.; Westberg, E.; Hanin, N.; Dag, A.; Ben-Ari, G.; Fragman-Sapir, O.; Tugendhaft, Y.; Kerem, Z.; Kadereit, J.W. Genetic variation of naturally growing olive trees in Israel: From abandoned groves to feral and wild? BMC Plant Biol. 2016, 16, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Zohary, D.; Hopf, M.; Weiss, E. Domestication of Plants in the Old World: The Origin and Spread of Cultivated Plants in Southwest Asia, Europe, and the Mediterranean Basin; Oxford University Press: London, UK, 2012. [Google Scholar]
- FAO. FAOSTAT. 2019. Available online: http://www.fao.org/faostat/en/#home (accessed on 26 December 2019).
- Figueiral, I.; Terral, J.-F. Late Quaternary refugia of Mediterranean taxa in the Portuguese Estremadura: Charcoal based palaeovegetation and climatic reconstruction. Quat. Sci. Rev. 2002, 21, 549–558. [Google Scholar] [CrossRef]
- Carrión, Y.; Ntinou, M.; Badal, E. Olea europaea L. in the North Mediterranean Basin during the Pleniglacial and the Early–Middle Holocene. Quat. Sci. Rev. 2010, 29, 952–968. [Google Scholar] [CrossRef] [Green Version]
- Kislev, M.; Nadel, D.; Carmi, I. Epipalaeolithic (19,000 BP) cereal and fruit diet at Ohalo II, Sea of Galilee, Israel. Rev. Palaeobot. Palynol. 1992, 73, 161–166. [Google Scholar] [CrossRef]
- Snir, A.; Nadel, D.; Groman-Yaroslavski, I.; Melamed, Y.; Sternberg, M.; Bar-Yosef, O.; Weiss, E. The Origin of Cultivation and Proto-Weeds, Long Before Neolithic Farming. PLoS ONE 2015, 10, e0131422. [Google Scholar] [CrossRef] [Green Version]
- Kaniewski, D.; Van Campo, E.; Boiy, T.; Terral, J.-F.; Khadari, B.; Besnard, G. Primary domestication and early uses of the emblematic olive tree: Palaeobotanical, historical and molecular evidence from the Middle East. Biol. Rev. 2012, 87, 885–899. [Google Scholar] [CrossRef] [Green Version]
- Langgut, D.; Adams, M.J.; Finkelstein, I. Climate, settlement patterns and olive horticulture in the southern Levant during the Early Bronze and Intermediate Bronze Ages (c.3600–1950 BC). Levant 2016, 48, 117–134. [Google Scholar] [CrossRef]
- Langgut, D.; Cheddadi, R.; Carrión, J.S.; Cavanagh, M.; Colombaroli, D.; Eastwood, W.J.; Greenberg, R.; Litt, T.; Mercuri, A.M.; Miebach, A.; et al. The origin and spread of olive cultivation in the Mediterranean Basin: The fossil pollen evidence. Holocene 2019, 29, 902–922. [Google Scholar] [CrossRef]
- Badal, E.; Bernabeu, J.; Vernet, J.; García, E.B. Vegetation changes and human action from the Neolithic to the Bronze Age (7000–4000 B.P.) in Alicante, Spain, based on charcoal analysis. Veg. Hist. Archaeobotany 1994, 3, 155–166. [Google Scholar] [CrossRef]
- Heinz, C.; Figueira, I.; Terral, J.J.-F.; Claustre, F. Holocene vegetation changes in the northwestern Mediterranean: New palaeoecological data from charcoal analysis and quantitative eco-anatomy. Holocene 2004, 14, 621–627. [Google Scholar] [CrossRef]
- Breton, C.; Terral, J.-F.; Pinatel, C.; Médail, F.; Bonhomme, F.; Bervillé, A. The origins of the domestication of the olive tree. Comptes Rendus Biol. 2009, 332, 1059–1064. [Google Scholar] [CrossRef]
- Besnard, G.; Baradat, P.; Breton, C.; Khadari, B.; Bervillé, A. Olive domestication from structure of oleasters and cultivars using nuclear RAPDs and mitochondrial RFLPs. Genet. Sel. Evol. 2001, 33, S251–S268. [Google Scholar] [CrossRef] [Green Version]
- Besnard, G.; Bakkali, A.E.; Haouane, H.; Baali-Cherif, D.; Moukhli, A.; Khadari, B. Population genetics of Mediterranean and Saharan olives: Geographic patterns of differentiation and evidence for early generations of admixture. Ann. Bot. 2013, 112, 1293–1302. [Google Scholar] [CrossRef] [Green Version]
- Besnard, G.; Khadari, B.; Navascués, M.; Fernández-Mazuecos, M.; Bakkali, A.E.; Arrigo, N.; Baali-Cherif, D.; De Caraffa, V.B.-B.; Santoni, S.; Vargas, P.; et al. The complex history of the olive tree: From Late Quaternary diversification of Mediterranean lineages to primary domestication in the northern Levant. Proc. R. Soc. B Boil. Sci. 2013, 280, 20122833. [Google Scholar] [CrossRef] [Green Version]
- Díez, C.M.; Trujillo, I.; Martinez-Urdiroz, N.; Barranco, D.; Rallo, L.; Marfil, P.; Gaut, B.S.; Ruiz, P.F.M. Olive domestication and diversification in the Mediterranean Basin. N. Phytol. 2015, 206, 436–447. [Google Scholar] [CrossRef]
- Besnard, G.; Terral, J.-F.; Cornille, A. On the origins and domestication of the olive: A review and perspectives. Ann. Bot. 2017, 121, 385–403. [Google Scholar] [CrossRef] [Green Version]
- Liphschitz, N.; Gophna, R.; Hartman, M.; Biger, G. The beginning of olive (olea europaea) cultivation in the old world: A reassessment. J. Archaeol. Sci. 1991, 18, 441–453. [Google Scholar] [CrossRef]
- Galili, E.; Stanley, D.J.; Sharvit, J.; Weinstein-Evron, M. Evidence for Earliest Olive-Oil Production in Submerged Settlements off the Carmel Coast, Israel. J. Archaeol. Sci. 1997, 24, 1141–1150. [Google Scholar] [CrossRef]
- Newton, C.; Lorre, C.; Sauvage, C.; Ivorra, S.; Terral, J.-F. On the origins and spread of Olea europaea L. (olive) domestication: Evidence for shape variation of olive stones at Ugarit, Late Bronze Age, Syria—A window on the Mediterranean Basin and on the westward diffusion of olive varieties. Veg. Hist. Archaeobotany 2014, 23, 567–575. [Google Scholar] [CrossRef]
- Dighton, A.; Fairbairn, A.S.; Bourke, S.; Faith, J.T.; Habgood, P. Bronze Age olive domestication in the north Jordan valley: New morphological evidence for regional complexity in early arboricultural practice from Pella in Jordan. Veg. Hist. Archaeobotany 2017, 26, 403–413. [Google Scholar] [CrossRef]
- Brun, J.-P. Archéologie du vin et de L’huile. De la Préhistoire à L’époque Hellénistique; Errance: Paris, France, 2004. [Google Scholar]
- Salavert, A. Olive cultivation and oil production in Palestine during the early Bronze Age (3500–2000 b.c.): The case of Tel Yarmouth, Israel. Veg. Hist. Archaeobotany 2008, 17, 53–61. [Google Scholar] [CrossRef]
- Lovell, J.L.; Meadows, J.; E Jacobsen, G. Upland Olive Domestication in the Chalcolithic Period: New 14C Determinations from El-Khawarij (Ajlun), Jordan. Radiocarbon 2010, 52, 364–371. [Google Scholar] [CrossRef] [Green Version]
- Neef, R. Introduction, Development and Environmental Implications of Olive Culture: The Evidence from Jordan. In Man’s Role in the Shaping of the Eastern Mediterranean Landscape; Bottema, S., Entjes-Nieborg, G., van Zeist, W., Eds.; Balkema Publishers: Rotterdam, The Netherlands, 1990; pp. 295–306. [Google Scholar]
- Newton, C.; Terral, J.-F.; Ivorra, S. The Egyptian olive (Olea europaeasubsp.europaea) in the later first millennium BC: Origins and history using the morphometric analysis of olive stones. Antiquity 2006, 80, 405–414. [Google Scholar] [CrossRef]
- Galili, E.; Langgut, D.; Terral, J.F.; Barazani, O.; Dag, A.; Horwitz, L.K.; Ramirez, I.O.; Rosen, B.; Weinstein-Evron, M.; Chaim, S.; et al. Early production of table olives at a mid-7th millennium BP submerged site off the Carmel coast (Israel). Sci. Rep. 2021, 11, 1–15. [Google Scholar] [CrossRef]
- Kislev, M.E. Wild olive endocarp at submerged Chalcolithic Kfar Samir, Haifa, Israel. J. Israel Prehist. Soc. 1995, 26, 134–145. [Google Scholar]
- Terral, J.-F.; Arnold-Simard, G. Beginnings of Olive Cultivation in Eastern Spain in Relation to Holocene Bioclimatic Changes. Quat. Res. 1996, 46, 176–185. [Google Scholar] [CrossRef]
- Terral, J.-F. Exploitation and Management of the Olive Tree During Prehistoric Times in Mediterranean France and Spain. J. Archaeol. Sci. 2000, 27, 127–133. [Google Scholar] [CrossRef]
- Rowan, E. Olive Oil Pressing Waste as a Fuel Source in Antiquity. Am. J. Archaeol. 2015, 119, 465–482. [Google Scholar] [CrossRef] [Green Version]
- Tanasi, D.; Greco, E.; Noor, R.E.; Feola, S.; Kumar, V.; Crispino, A.; Gelis, I. 1H NMR, 1H–1H 2D TOCSY and GC-MS analyses for the identification of olive oil in Early Bronze Age pottery from Castelluccio (Noto, Italy). Anal. Methods 2018, 10, 2756–2763. [Google Scholar] [CrossRef]
- Gros-Balthazard, M.; Besnard, G.; Sarah, G.; Holtz, Y.; Leclercq, J.; Santoni, S.; Wegmann, D.; Glémin, S.; Khadari, B. Evolutionary transcriptomics reveals the origins of olives and the genomic changes associated with their domestication. Plant J. 2019, 100, 143–157. [Google Scholar] [CrossRef] [PubMed]
- Terral, J.-F.; Alonso, N.; Capdevila, R.B.I.; Chatti, N.; Fabre, L.; Fiorentino, G.; Marinval, P.; Jordà, G.P.; Pradat, B.; Rovira, N.; et al. Historical biogeography of olive domestication (Olea europaea L.) as revealed by geometrical morphometry applied to biological and archaeological material. J. Biogeogr. 2003, 31, 63–77. [Google Scholar] [CrossRef]
- Brun, J.-P. Archéologie du vin et de L’huile Dans l’Empire Romain; Errance: Paris, France, 2004. [Google Scholar]
- Laubenheimer, F. Le Temps des Amphores en Gaule. Vins, Huiles et Sauces; Errance: Paris, France, 1990. [Google Scholar]
- André, J. L’Alimentation et la Cuisine à Rome; Les Belles Lettres: Paris, France, 1981. [Google Scholar]
- Terral, J.-F.; Tabard, E.; Bouby, L.; Ivorra, S.; Pastor, T.; Figueiral, I.; Picq, S.; Chevance, J.-B.; Jung, C.; Fabre, L.; et al. Evolution and history of grapevine (Vitis vinifera) under domestication: New morphometric perspectives to understand seed domestication syndrome and reveal origins of ancient European cultivars. Ann. Bot. 2009, 105, 443–455. [Google Scholar] [CrossRef]
- Bonhomme, V.; Picq, S.; Ivorra, S.; Evin, A.; Pastor, T.; Bacilieri, R.; Lacombe, T.; Figueiral, I.; Terral, J.-F.; Bouby, L. Eco-evo-devo implications and archaeobiological perspectives of trait covariance in fruits of wild and domesticated grapevines. PLoS ONE 2020, 15, e0239863. [Google Scholar] [CrossRef] [PubMed]
- R. R: The R Project for Statistical Computing. R, 2021. Available online: https://www.r-project.org/ (accessed on 16 September 2021).
- Bonhomme, V.; Picq, S.; Gaucherel, C.; Claude, J. Momocs: Outline Analysis Using R. J. Stat. Softw. 2014, 56, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Bonhomme, V.; Picq, S.; Claude, J. Momocs Package 2020. Available online: https://cran.r-project.org (accessed on 16 September 2021).
- Venables, V.N.; Ripley, B.D. Modern Applied Statistics with R; Springer: New York, NY, USA, 2002. [Google Scholar]
- Paradis, E.; Schliep, K. ape 5.0: An environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics 2018, 35, 526–528. [Google Scholar] [CrossRef]
- Burger, P.; Terral, J.-F.; Ruas, M.-P.; Ivorra, S.; Picq, S. Assessing past agrobiodiversity of Prunus avium L. (Rosaceae): A morphometric approach focussed on the stones from the archaeological site Hôtel-Dieu (16th century, Tours, France). Veg. Hist. Archaeobotany 2011, 20, 447–458. [Google Scholar] [CrossRef]
- Szokefalvi-Nagy, B. Introduction to Real Functions and Orthogonal Expansions. University Texts in the Mathematical Sciences; Oxford University Press: Oxford, UK, 1965. [Google Scholar]
- Blondel, J.; Aronson, J. Biodiversity and ecosystem function in the Mediterranean Basin: Human and non-human determi-nants. In The Function of Biodiversity; David, G.X., Richardson, D.M., Eds.; Springer: New York, NY, USA, 1995; pp. 43–119. [Google Scholar]
- Bakkali, A.E.; Haouane, H.; Moukhli, A.; Costes, E.; Van Damme, P.; Khadari, B. Construction of Core Collections Suitable for Association Mapping to Optimize Use of Mediterranean Olive (Olea europaea L.) Genetic Resources. PLoS ONE 2013, 8, e61265. [Google Scholar] [CrossRef] [Green Version]
- Khadari, B.; Bakkali, A.E.; Essalouh, L.; Tollon, C.; Pinatel, C.; Besnard, G. Cultivated Olive Diversification at Local and Regional Scales: Evidence From the Genetic Characterization of French Genetic Resources. Front. Plant Sci. 2019, 10, 1593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terral, J.-F. La Domestication de L’olivier (Olea europaea L.) en Méditerranée Nord-Occidentale: Approches Morphométriques et Implications Paléoclimatiques. Ph.D. Thesis, Université de Montpellier 2, Montpellier, France, 1997. [Google Scholar]
- Margaritis, E. Distinguishing exploitation, domestication, cultivation and production: The olive in the third millennium Aegean. Antiquity 2013, 87, 746–757. [Google Scholar] [CrossRef]
- Ramos-Madrigal, J.; Runge, A.K.W.; Bouby, L.; Lacombe, T.; Castruita, J.A.S.; Adam-Blondon, A.-F.; Figueiral, I.; Hallavant, C.; Martínez-Zapater, J.M.; Schaal, C.; et al. Palaeogenomic insights into the origins of French grapevine diversity. Nat. Plants 2019, 5, 595–603. [Google Scholar] [CrossRef] [PubMed]
- Bonhomme, V.; Terral, J.-F.; Zech-Matterne, V.; Ivorra, S.; Lacombe, T.; Deborde, G.; Kuchler, P.; Limier, B.; Pastor, T.; Rollet, P.; et al. Seed morphology uncovers 1500 years of vine agrobiodiversity before the advent of the Champagne wine. Sci. Rep. 2021, 11, 1–14. [Google Scholar] [CrossRef]
- Terral, J.-F.; Newton, C.; Ivorra, S.; Gros-Balthazard, M.; de Morais, C.T.; Picq, S.; Tengberg, M.; Pintaud, J.-C. Insights into the historical biogeography of the date palm (Phoenix dactylifera L.) using geometric morphometry of modern and ancient seeds. J. Biogeogr. 2012, 39, 929–941. [Google Scholar] [CrossRef]
- Gros-Balthazard, M.; Newton, C.; Ivorra, S.; Pierre, M.-H.; Pintaud, J.-C.; Terral, J.-F. The Domestication Syndrome in Phoenix dactylifera Seeds: Toward the Identification of Wild Date Palm Populations. PLoS ONE 2016, 11, e0152394. [Google Scholar] [CrossRef]
- Besnard, G.; De Casas, R.R. Single vs multiple independent olive domestications: The jury is (still) out. New Phytol. 2016, 209, 466–470. [Google Scholar] [CrossRef] [Green Version]
- Bourgeon, O.; Pagnoux, C.; Mauné, S.; Vargas, E.G.; Ivorra, S.; Bonhomme, V.; Ater, M.; Moukhli, A.; Terral, J.-F. Olive tree varieties cultivated for the great Baetican oil trade between the 1st and the 4th centuries ad: Morphometric analysis of olive stones from Las Delicias (Ecija, Province of Seville, Spain). Veg. Hist. Archaeobotany 2017, 27, 1–14. [Google Scholar] [CrossRef]
- Bouby, L.; Figueiral, I.; Bouchette, A.; Rovira, N.; Ivorra, S.; Lacombe, T.; Pastor, T.; Picq, S.; Marinval, P.; Terral, J.-F. Bioarchaeological Insights into the Process of Domestication of Grapevine (Vitis vinifera L.) during Roman Times in Southern France. PLoS ONE 2013, 8, e63195. [Google Scholar] [CrossRef] [Green Version]
- Gros-Balthazard, M.; Galimberti, M.; Kousathanas, A.; Newton, C.; Ivorra, S.; Paradis, L.; Vigouroux, Y.; Carter, R.; Tengberg, M.; Battesti, V.; et al. The Discovery of Wild Date Palms in Oman Reveals a Complex Domestication History Involving Centers in the Middle East and Africa. Curr. Biol. 2017, 27, 2211–2218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ater, M.; Barbara, H.; Kassout, J. Importance des variétés locales, de l’oléastre et des pratiques traditionnelles de l’oléiculture dans la région de Chefchaouen (Nord du Maroc). In L’oléiculture au Maroc de la Préhistoire à nos Jours: Pratiques, Diversité, Adaptation, Usages, Commerce et Politiques; Ater, M., Essalouh, L., Ilbert, H., Moukhli, A., Khadari, B., Eds.; Options Mé-diterranéennes, CIHEAM: Montpellier, France, 2016; pp. 109–121. [Google Scholar]
- Aumeeruddy-Thomas, Y.; Moukhli, A.; Haouane, H.; Khadari, B. Ongoing domestication and diversification in grafted olive–oleaster agroecosystems in Northern Morocco. Reg. Environ. Chang. 2017, 17, 1315–1328. [Google Scholar] [CrossRef]
- Waddington, C.H. Canalization of Development and The Inheritance of Acquired Characters. Nat. Cell Biol. 1942, 150, 563–565. [Google Scholar] [CrossRef]
- Siegal, M.; Bergman, A. Waddington’s canalization revisited: Developmental stability and evolution. Proc. Natl. Acad. Sci. USA 2002, 99, 10528–10532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pagnoux, C. Emergence, Développement et Diversification de L’arboriculture en Grèce du Néolithique à L’époque Romaine. Confrontation des Données Archéobotaniques, Morphométriques, Epigraphiques et Littéraires. Ph.D. Thesis, Université Paris 1—Panthéon-Sorbonne, Paris, France, 2016. [Google Scholar]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Terral, J.-F.; Bonhomme, V.; Pagnoux, C.; Ivorra, S.; Newton, C.; Paradis, L.; Ater, M.; Kassout, J.; Limier, B.; Bouby, L.; et al. The Shape Diversity of Olive Stones Resulting from Domestication and Diversification Unveils Traits of the Oldest Known 6500-Years-Old Table Olives from Hishuley Carmel Site (Israel). Agronomy 2021, 11, 2187. https://doi.org/10.3390/agronomy11112187
Terral J-F, Bonhomme V, Pagnoux C, Ivorra S, Newton C, Paradis L, Ater M, Kassout J, Limier B, Bouby L, et al. The Shape Diversity of Olive Stones Resulting from Domestication and Diversification Unveils Traits of the Oldest Known 6500-Years-Old Table Olives from Hishuley Carmel Site (Israel). Agronomy. 2021; 11(11):2187. https://doi.org/10.3390/agronomy11112187
Chicago/Turabian StyleTerral, Jean-Frédéric, Vincent Bonhomme, Clémence Pagnoux, Sarah Ivorra, Claire Newton, Laure Paradis, Mohammed Ater, Jalal Kassout, Bertrand Limier, Laurent Bouby, and et al. 2021. "The Shape Diversity of Olive Stones Resulting from Domestication and Diversification Unveils Traits of the Oldest Known 6500-Years-Old Table Olives from Hishuley Carmel Site (Israel)" Agronomy 11, no. 11: 2187. https://doi.org/10.3390/agronomy11112187