
Genetic Diversity of Tamarixia radiata Populations and Their Associated Endosymbiont Wolbachia Species from China

 , , , ,
, , , ,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Insects and Rearing
2.2. Biological Aspects (Parasitism, Emergence, and Sex Ratio) of Different Populations of Tamarixia radiata
2.3. DNA Extraction and Isolation
2.4. Amplification of DNA and Sequencing
2.5. Sequence Composition and Phylogenetic Analysis
2.6. Genetic Diversity and Distribution Analysis
2.7. Data Analysis
3. Results
3.1. Efficacy of Parasitism, Emergence, and Sex Ratio on a Different Population of Tamarixia radiata
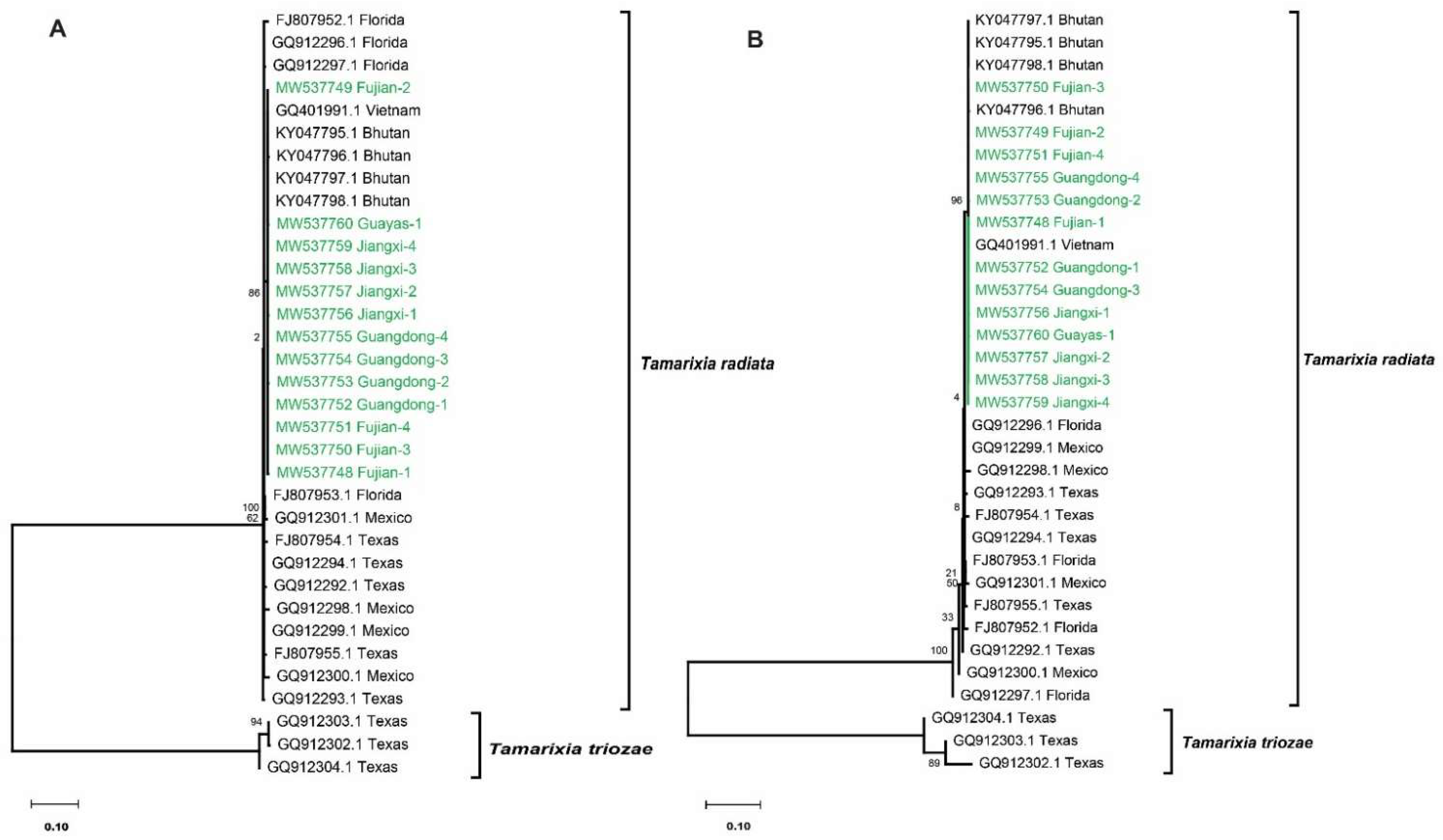
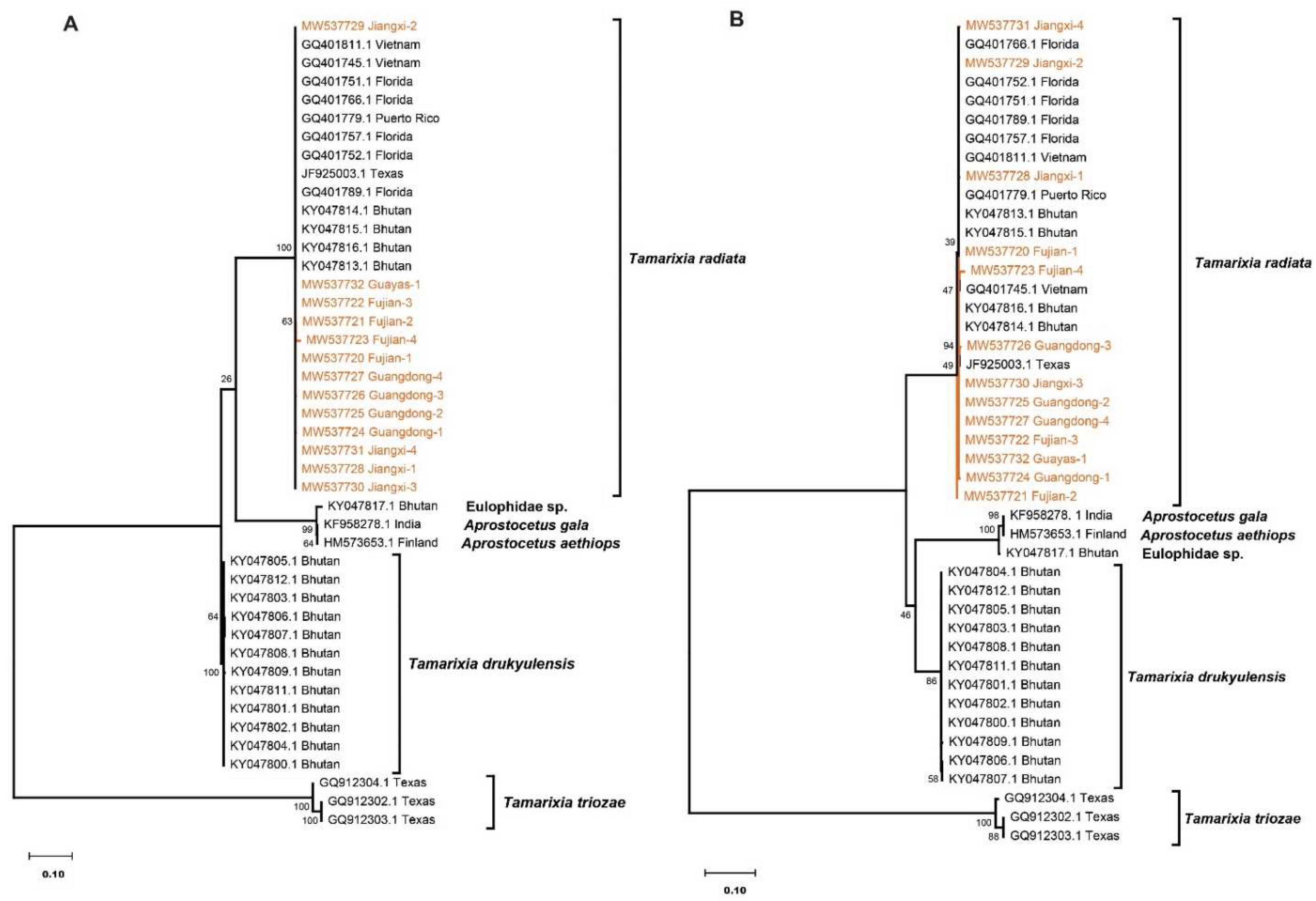
3.2. Sequence and Phylogenetic Analysis of the ITS1 Region
3.3. Sequence and Phylogenetic Analysis of the ITS2 Region
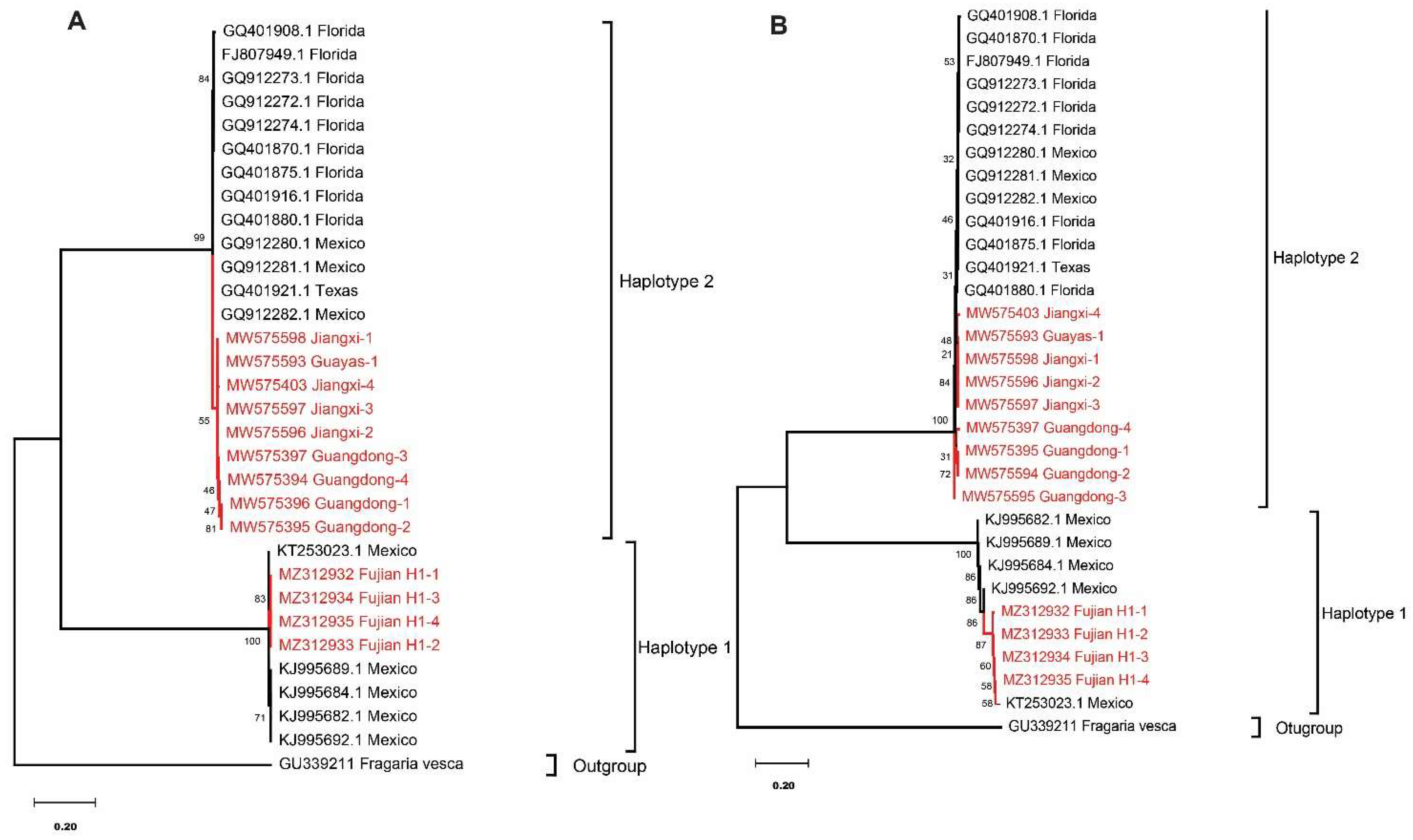
3.4. Sequence and Phylogenetic Analysis of COI
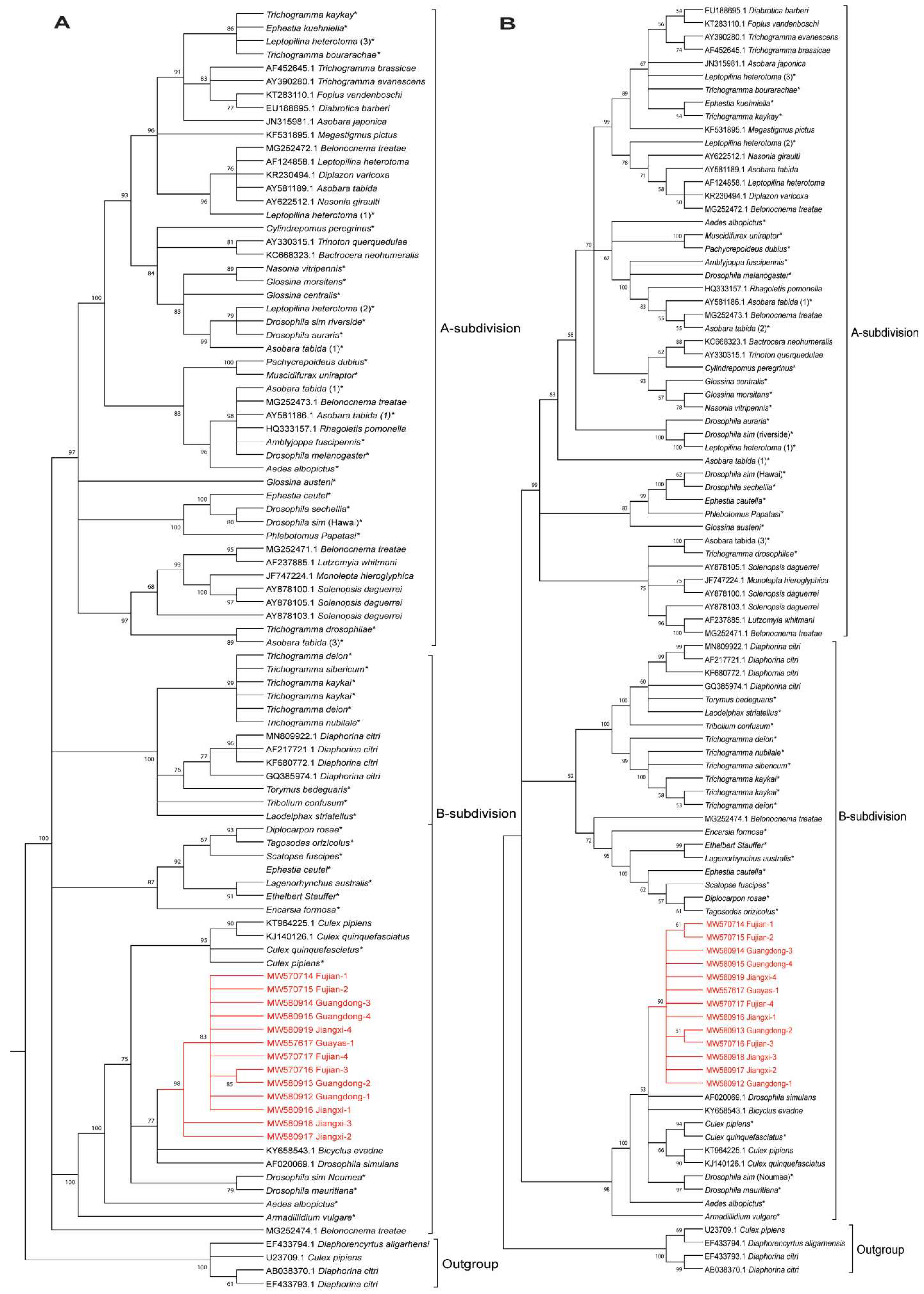
3.5. Phylogenetic Analysis of Wolbachia Based on the wsp Gene
3.6. Population Genetic Diversity Analysis
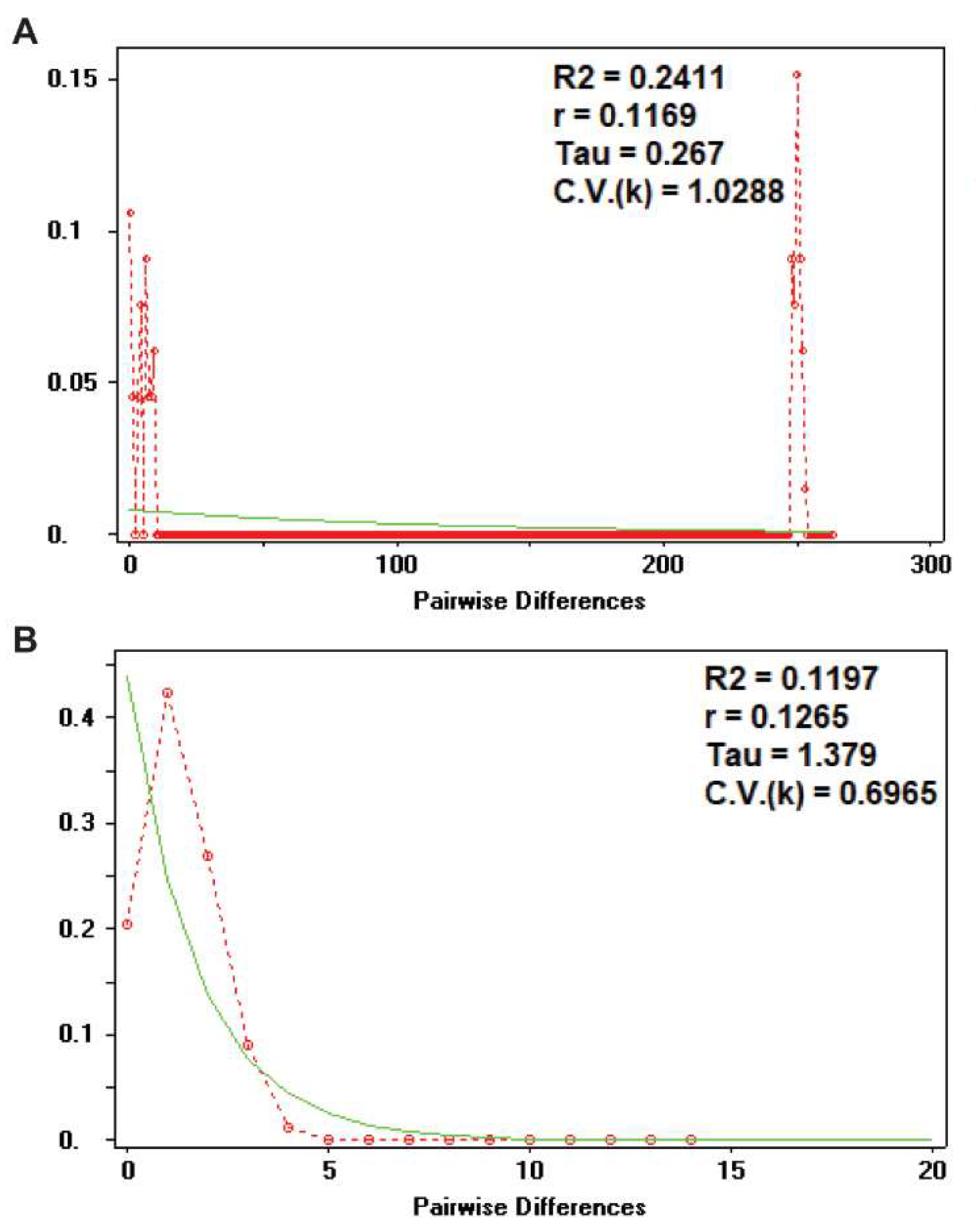
3.7. Demographic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- LaSalle, J.; Gauld, I. Parasitic Hymenoptera and the biodiversity crisis. Redia 1991, 74, 315–334. [Google Scholar]
- Kosheleva, O.V.; Kostjukov, V. V Types of parasitism in eulophid wasps (Hymenoptera, Eulophidae). Entomol. Rev. 2014, 94, 1202–1217. [Google Scholar] [CrossRef]
- Eggleton, P.; Gaston, K.J. “Parasitoid” species and assemblages: Convenient definitions or misleading compromises? Oikos 1990, 59, 417–421. [Google Scholar] [CrossRef]
- Kuris, A.M. Trophic Interactions: Similarity of Parasitic Castrators to Parasitoids. Q. Rev. Biol. 1974, 49, 129–148. [Google Scholar] [CrossRef] [Green Version]
- Chien, C.-C.; Chiu, S.-C.; Ku, S.-C. Biological control of Diaphorina citri in Taiwan. Fruits 1989, 44, 401–407. [Google Scholar]
- Étienne, J.; Quilici, S.; Marival, D.; Franck, A.; Gonzalez Fernandez, C. Biological control of Diaphorina citri (Hemiptera: Psyllidae) in Guadeloupe by imported Tamarixia radiata (Hymenoptera: Eulophidae). Fruits 2001, 56, 307–315. [Google Scholar] [CrossRef] [Green Version]
- Hoddle, M.S. Foreign exploration for natural enemies of Asian citrus psyllid, Diaphorina citri (Hemiptera: Psyllidae), in the Punjab of Pakistan for use in a classical biological control program in California USA. Pakistan Entomol. 2012, 34, 1–5. [Google Scholar]
- Hoy, M.A.; Nguyen, R.; Jeyaprakash, A. Classical biological control of Asian citrus psylla. Citrus Ind. 2001, 81, 48–50. [Google Scholar]
- Qureshi, J.A.; Rogers, M.E.; Hall, D.G.; Stansly, P.A. Incidence of invasive Diaphorina citri (Hemiptera: Psyllidae) and its introduced parasitoid Tamarixia radiata (Hymenoptera: Eulophidae) in Florida citrus. J. Econ. Entomol. 2009, 102, 247–256. [Google Scholar] [CrossRef]
- Kumagai, L.B.; LeVesque, C.S.; Blomquist, C.L.; Madishetty, K.; Guo, Y.; Woods, P.W.; Rooney-Latham, S.; Rascoe, J.; Gallindo, T.; Schnabel, D.; et al. First report of Candidatus liberibacter asiaticus associated with citrus huanglongbing in California. Plant Dis. 2013, 97, 283. [Google Scholar] [CrossRef] [PubMed]
- Razi, M.F.; Keremane, M.L.; Ramadugu, C.; Roose, M.; Khan, I.A.; Lee, R.F. Detection of citrus huanglongbing-associated “Candidatus Liberibacter asiaticus” in citrus and Diaphorina citri in Pakistan, seasonal variability, and implications for disease management. Phytopathology 2014, 104, 257–268. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Yin, Y.; Hu, H.; Yuan, Q.; Peng, G.; Xia, Y. Development and application of molecular-based diagnosis for ‘Candidatus Liberibacter asiaticus’, the causal pathogen of citrus huanglongbing. Plant Pathol. 2006, 55, 630–638. [Google Scholar] [CrossRef]
- Deng, X.-L.; Gao, Y.-D.; Jian-Chi, C.; Xue-Lian, P.U.; Kong, W.-W.; Li, H.-P. Curent Situation of “Candidatus Liberibacter asiaticus” in Guangdong, China, Where Citrus Huanglongbing Was First Described. J. Integr. Agric. 2012, 11, 424–429. [Google Scholar] [CrossRef]
- Fan, G.-C.; Liu, B.; Wu, R.-J.; Li, T.; Cai, Z.-J.; Ke, C. Thirty years of research on citrus huanglongbing in China. Fujian J. Agric. Sci. 2009, 24, 183–190. [Google Scholar]
- Hall, D.G.; Richardson, M.L.; Ammar, E.D.; Halbert, S.E. Asian citrus psyllid, Diaphorina citri, vector of citrus huanglongbing disease. Entomol. Exp. Appl. 2013, 146, 207–223. [Google Scholar] [CrossRef]
- Yang, Y.; Huang, M.; Andrew, G.; Beattie, C.; Xia, Y.; Ouyang, G.; Xiong, J. Distribution, biology, ecology and control of the psyllid Diaphorina citri Kuwayama, a major pest of citrus: A status report for China. Taylor Fr. 2006, 52, 343–352. [Google Scholar]
- Ke, S.; Li, K.B.; Ke, C.; Tsai, J.H. Transmission of the huanglungbin agent from citrus to periwinkle by dodder. In Proceedings of the International Organization of Citrus Virologists Conference, California, CA, USA, 1988; Volume 10. [Google Scholar]
- Ibarra-Cortés, K.H.; González-Hernández, H.; Guzmán-Franco, A.W. Susceptibility of nymphs and adults of Diaphorina citri to the entomophathogenic fungus Hirsutella citriformis. Biocontrol Sci. Technol. 2017, 27, 433–438. [Google Scholar] [CrossRef]
- Juan-Blasco, M.; Qureshi, J.A.; Urbaneja, A.; Stansly, P.A. Predatory mite, Amblyseius swirskii (Acari: Phytoseiidae), for biological control of Asian citrus psyllid, Diaphorina citri (Hemiptera: Psyllidae). Florida Entomol. 2012, 95, 543–551. [Google Scholar] [CrossRef]
- Patt, J.M.; Rohrig, E. Laboratory evaluations of the foraging success of Tamarixia radiata (Hymenoptera: Eulophidae) on flowers and extrafloral nectaries: Potential use of nectar plants for conservation biological control of Asian citrus psyllid (Hemiptera: Liviidae). Florida Entomol. 2017, 100, 149–156. [Google Scholar] [CrossRef] [Green Version]
- Pluke, R.W.H.; Qureshi, J.A.; Stansly, P.A. Citrus flushing patterns, Diaphorina citri (Hemiptera: Psyllidae) populations and parasitism by Tamarixia radiata (Hymenoptera: Eulophidae) in Puerto Rico. Florida Entomol. 2008, 91, 36–42. [Google Scholar] [CrossRef]
- Li, Y.-H.; Zhou, Y.-T.; Guo, C.-F.; Ou, D.; Qureshi, J.A.; Sang, W.; Qiu, B.-L. Temperature and host age effects on the life history of Tamarixia radiata, a dominant parasitoid of citrus psyllid Diaphorina citri. Crop Prot. 2018, 114, 32–38. [Google Scholar] [CrossRef]
- Gómez-Torres, M.L.; Nava, D.E.; Parra, J.R.P. Life table of Tamarixia radiata (Hymenoptera: Eulophidae) on Diaphorina citri (Hemiptera: Psyllidae) at different temperatures. J. Econ. Entomol. 2012, 105, 338–343. [Google Scholar] [CrossRef] [PubMed]
- Barr, N.B.; Hall, D.G.; Weathersbee, A.A.; Nguyen, R.; Stansly, P.; Qureshi, J.A.; Flores, A.D. Comparison of Laboratory Colonies and Field Populations of Tamarixia radiata, an Ectoparasitoid of the Asian Citrus Psyllid, Using Internal Transcribed Spacer and Cytochrome Oxidase Subunit I DNA Sequences. J. Econ. Entomol. 2009, 102, 2325–2332. [Google Scholar] [CrossRef]
- Om, N.; Yefremova, Z.A.; Yegorenkova, E.N.; Beattie, G.A.C.; Donovan, N.J.; Holford, P. A new species of Tamarixia Mercet (Hymenoptera, Eulophidae), a parasitoid of Diaphorina communis Mathur (Hemiptera, Liviidae) in Bhutan. J. Asia. Pac. Entomol. 2017, 20, 728–738. [Google Scholar] [CrossRef]
- Qasim, M.; Baohua, W.; Zou, H.; Lin, Y.; Dash, C.K.; Bamisile, B.S.; Hussain, M.; Zhiwen, Z.; Wang, L. Phylogenetic relationship and genetic diversity of citrus psyllid populations from China and Pakistan and their associated Candidatus bacterium. Mol. Phylogenet. Evol. 2018, 126, 173–180. [Google Scholar] [CrossRef]
- De León, J.H.; Sétamou, M. Molecular evidence suggests that populations of the Asian citrus psyllid parasitoid Tamarixia radiata (Hymenoptera: Eulophidae) from Texas, Florida, and Mexico represent a single species. Ann. Entomol. Soc. Am. 2010, 103, 100–110. [Google Scholar] [CrossRef]
- Lunt, D.H.; Zhang, D.X.; Szymura, J.M.; Hewitt, G.M. The insect cytochrome oxidase I gene: Evolutionary patterns and conserved primers for phylogenetic studies. Insect Mol. Biol. 1996, 5, 153–165. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Ratnasingham, S.; Dewaard, J.R. Barcoding animal life: Cytochrome c oxidase subunit 1 divergences among closely related species. Proc. R. Soc. B Biol. Sci. 2003, 270, s96–s99. [Google Scholar] [CrossRef] [Green Version]
- Ji, Y.-J.; Zhang, D.-X.; He, L.-J. Undefined Evolutionary conservation and versatility of a new set of primers for amplifying the ribosomal internal transcribed spacer regions in insects and other invertebrates. Mol. Ecol. Notes 2003, 3, 581–585. [Google Scholar] [CrossRef] [Green Version]
- Grenier, S.; Gomes, S.M.; Pintureau, B.; Lassablière, F.; Bolland, P. Use of tetracycline in larval diet to study the effect of Wolbachia on host fecundity and clarify taxonomic status of Trichogramma species in cured bisexual lines. J. Invertebr. Pathol. 2002, 80, 13–21. [Google Scholar] [CrossRef]
- Douglas, A.E. Buchnera bacteria and other symbionts of aphids. Insect Symbiosis 2003, 1, 23–38. [Google Scholar]
- Hosokawa, T.; Koga, R.; Kikuchi, Y.; Meng, X.-Y.; Fukatsu, T. Wolbachia as a bacteriocyte-associated nutritional mutualist. Proc. Natl. Acad. Sci. USA 2010, 107, 769–774. [Google Scholar] [CrossRef] [Green Version]
- Huigens, M.E.; Hohmann, C.L.; Luck, R.F.; Gort, G.; Stouthamer, R. Reduced competitive ability due to Wolbachia infection in the parasitoid wasp Trichogramma kaykai. Entomol. Exp. Appl. 2004, 110, 115–123. [Google Scholar] [CrossRef]
- Kikuchi, Y.; Hayatsu, M.; Hosokawa, T.; Nagayama, A.; Tago, K.; Fukatsu, T. Symbiont-mediated insecticide resistance. Proc. Natl. Acad. Sci. USA 2012, 109, 8618–8622. [Google Scholar] [CrossRef] [Green Version]
- Min, K.-T.; Benzer, S. Wolbachia, normally a symbiont of Drosophila, can be virulent, causing degeneration and early death. Proc. Natl. Acad. Sci. USA 1997, 94, 10792–10796. [Google Scholar] [CrossRef] [Green Version]
- O’Neill, S.L.; Hoffmann, A.; Werren, J. Influential Passengers: Inherited Microorganisms and Arthropod Reproduction; Oxford University Press: New York, NY, USA, 1997; ISBN 0198577869. [Google Scholar]
- Saridaki, A.; Bourtzis, K. Wolbachia: More than just a bug in insects genitals. Curr. Opin. Microbiol. 2010, 13, 67–72. [Google Scholar] [CrossRef]
- Tagami, Y.; Miura, K.; Stouthamer, R. Positive effect of fertilization on the survival rate of immature stages in a Wolbachia-associated thelytokous line of Trichogramma deion and T. kaykai. Entomol. Exp. Et Applicata. 2002, 105, 165–167. [Google Scholar] [CrossRef]
- Wong, Z.S.; Hedges, L.M.; Brownlie, J.C.; Johnson, K.N. Wolbachia-mediated antibacterial protection and immune gene regulation in Drosophila. PLoS ONE 2011, 6, e25430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masui, S.; Kamoda, S.; Sasaki, T. Distribution and Evolution of Bacteriophage WO in Wolbachia, the Endosymbiont Causing Sexual Alterations in Arthropods. J. Mol. Evol. 2000, 51, 491–497. [Google Scholar] [CrossRef]
- Subandiyah, S.; Nikoh, N.; Tsuyumu, S.; Somowiyarjo, S.; Fukatsu, T. Complex endosymbiotic microbiota of the citrus psyllid Diaphorina citri (Homoptera: Psylloidea). Zoolog. Sci. 2000, 17, 983–989. [Google Scholar] [CrossRef]
- Fagen, J.R.; Giongo, A.; Brown, C.T.; Davis-Richardson, A.G.; Gano, K.A.; Triplett, E.W. Characterization of the Relative Abundance of the Citrus Pathogen Ca. Liberibacter Asiaticus in the Microbiome of Its Insect Vector, Diaphorina citri, using High Throughput 16S rRNA Sequencing. Open Microbiol. J. 2012, 6, 29–33. [Google Scholar] [CrossRef] [Green Version]
- Ramos Aguila, L.C.; Atlihan, R.; Ashraf, H.J.; Keppanan, R.; Lei, L.; Bamisile, B.S.; Cerda, H.; Wang, L. Temperature-Dependent Biological Control Effectiveness of Tamarixia radiata (Hymenoptera: Eulophidea) Under Laboratory Conditions. J. Econ. Entomol. 2021. [Google Scholar] [CrossRef]
- Braig, H.R.; Zhou, W.; Dobson, S.L.; O’neill, S.L. Cloning and Characterization of a Gene Encoding the Major Surface Protein of the Bacterial Endosymbiont Wolbachia pipientis. J. Bacteriol. 1998, 180, 2373–2378. [Google Scholar] [CrossRef] [Green Version]
- Simon, C.; Frati, F.; Beckenbach, A.; Crespi, B.; Liu, H.; Flook, P. Evolution, weighting, and phylogenetic utility of mitochondrial gene sequences and a compilation of conserved polymerase chain reaction primers. Ann. Entomol. Soc. Am. 1994, 87, 651–701. [Google Scholar] [CrossRef]
- Zhou, W.; Rousset, F.; O’Neill, S. Phylogeny and PCR-based classification of Wolbachia strains using wsp gene sequences. Proc. R. Soc. B Biol. Sci. 1998, 265, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. Inf. Retr. Ltd. 1999, 41, 95–98. [Google Scholar]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Nei, M.; Kumar, S. Molecular evolution and phylogenetics; Oxford University Press: New York, NY, USA, 2000; ISBN 0195350510. [Google Scholar]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harpending, H.C. Signature of ancient population growth in a low-resolution mitochondrial DNA mismatch distribution. Hum. Biol. 1994, 66, 591–600. [Google Scholar]
- Meyer, J.M.; Hoy, M.A. Molecular Survey of Endosymbionts in Florida Populations of Diaphorina citri (Hemiptera: Psyllidae) and Its Parasitoids „Tamarixia radiata (Hymenoptera: Eulophidae) and Diaphorencyrtus aligarhensis (Hymenoptera: Encyrtidae). Florida Entomol. 2008, 91, 294–304. [Google Scholar] [CrossRef]
- Vavre, F.; Fleury, F.; Lepetit, D.; Fouillet, P.; Boulétreau, M. Phylogenetic evidence for horizontal transmission of Wolbachia in host- parasitoid associations. Mol. Biol. Evol. 1999, 16, 1711–1723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bové, J.M. Huanglongbing: A destructive, newly-emerging, century-old disease of citrus. J. Plant Pathol. 2006, 88, 7–37. [Google Scholar]
- Mann, R.S.; Stelinski, L.L. An asian citrus psyllid parasitoid Tamarixia radiata (Waterston) (Insecta: Hymenoptera: Eulophidae). EDIS 2010, 2010. Available online: https://edis.ifas.ufl.edu/publication/IN858 (accessed on 1 May 2021).
- Chien, C.C.; Chu, Y.I.; Ku, S.C. Parasitic strategy, morphology and life history of Tamarixia radiata (Hymenoptera; Eulophidae). Chin. J. Entomol. 1991, 11, 264–281. [Google Scholar]
- Skelley, L.H.; Hoy, M.A. A synchronous rearing method for the Asian citrus psyllid and its parasitoids in quarantine. Biol. Control 2004, 29, 14–23. [Google Scholar] [CrossRef]
- Chien, C.C.; Chu, Y.I.; Ku, S.C. Influences of host densities on the population increase of the eulophid wasp, Tamarixia radiata, and its host-killing ability. Plant Prot. Bull 1995, 37, 81–96. [Google Scholar]
- Peña-Carrillo, K.I.; González-Hernández, A.; López-Arroyo, J.I.; Mercado-Hernández, R.; Favela-Lara, S. Morphological and Genetic Variation in Mexican Wild Populations of Tamarixia radiata (Hymenoptera: Eulophidae). Fla. Entomol. 2015, 98, 1093–1100. [Google Scholar] [CrossRef]
- Hillis, D.M.; Dixon, M.T. Ribosomal DNA: Molecular evolution and phylogenetic inference. Q. Rev. Biol. 1991, 66, 411–446. [Google Scholar] [CrossRef]
- Taylor, D.B.; Szalanski, A.L. Identification of Muscidifurax spp. by polymerase chain reaction-restriction fragment length polymorphism. Biol. Control 1999, 15, 270–273. [Google Scholar] [CrossRef] [Green Version]
- González-Hernández, A.; Canales-Castillo, R.; López-Arroyo, J.I.; Villanueva-Segura, O.K.; Arredondo-Bernal, H.C.; Jasso-Argumedo, J. Variabilidad morfológica y genética de Tamarixia radiata (Waterston, 1922)(Hymenoptera: Eulophidae), parasitoide del psílido asiático de los cítricos en México. In Proceedings of the XXXIII Congreso Nacional de Control Biológico, Uruapan, Mexico; 2010; pp. 257–259. [Google Scholar]
- Grant, W.A.S.; Bowen, B.W. Shallow population histories in deep evolutionary lineages of marine fishes: Insights from sardines and anchovies and lessons for conservation. J. Hered. 1998, 89, 415–426. [Google Scholar] [CrossRef]
- Tajima, F. Statistical method for testing the neutral mutation hypothesis by DNA polymorphism. Genetics 1989, 123, 585–595. [Google Scholar] [CrossRef]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [Green Version]
- Hilgenboecker, K.; Hammerstein, P.; Schlattmann, P.; Telschow, A.; Werren, J.H. How many species are infected with Wolbachia?—A statistical analysis of current data. FEMS Microbiol. Lett. 2008, 281, 215–220. [Google Scholar] [CrossRef] [Green Version]
- Dobson, S.L.; Bourtzis, K.; Braig, H.R.; Jones, B.F.; Zhou, W.; Rousset, F.; O’Neill, S.L. Wolbachia infections are distributed throughout insect somatic and germ line tissues. Insect Biochem. Mol. Biol. 1999, 29, 153–160. [Google Scholar] [CrossRef]
- Werren, J.H.; Baldo, L.; Clark, M.E. Wolbachia: Master manipulators of invertebrate biology. Nat. Rev. Microbiol. 2008, 6, 741–751. [Google Scholar] [CrossRef] [PubMed]
- Fenollar, F.; La Scola, B.; Inokuma, H.; Dumler, J.S.; Taylor, M.J.; Raoult, D. Culture and phenotypic characterization of a Wolbachia pipientis isolate. J. Clin. Microbiol. 2003, 41, 5434–5441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldo, L.; Dunning Hotopp, J.C.; Jolley, K.A.; Bordenstein, S.R.; Biber, S.A.; Choudhury, R.R.; Hayashi, C.; Maiden, M.C.J.; Tettelin, H.; Werren, J.H. Multilocus sequence typing system for the endosymbiont Wolbachia pipientis. Appl. Environ. Microbiol. 2006, 72, 7098–7110. [Google Scholar] [CrossRef] [Green Version]
- Baldo, L.; Werren, J.H. Revisiting Wolbachia supergroup typing based on WSP: Spurious lineages and discordance with MLST. Curr. Microbiol. 2007, 55, 81–87. [Google Scholar] [CrossRef]
- Wang, X.; Xiong, X.; Cao, W.; Zhang, C.; Werren, J.H.; Wang, X. Phylogenomic Analysis of Wolbachia Strains Reveals Patterns of Genome Evolution and Recombination. Genome Biol. Evol. 2020, 12, 2508–2520. [Google Scholar] [CrossRef]
- Saha, S.; Hunter, W.B.; Reese, J.; Morgan, J.K.; Marutani-Hert, M.; Huang, H.; Lindeberg, M. Survey of Endosymbionts in the Diaphorina citri Metagenome and Assembly of a Wolbachia wDi Draft Genome. PLoS ONE 2012, 7, e50067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Y.X.; Li, W.H. Statistical tests of neutrality of mutations. Genetics 1993, 133, 693–709. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Primer Sequence (5′-3′) | (Annealing Temp °C) | Expected Product Size (bp) |
|---|---|---|---|
| ITS1- 18s F | GTGAACCTGCGAAGGA | ||
| ITS1- 5.8s R | GTTCATGTCCTGCAGTTCACA | 55 °C for 30 sec | 503–515 |
| ITS2- 5.8s F | TGTGAACTGCAGGACACATGAAC | ||
| ITS2- 28s R | ATGCTTAAATTTAGGGGGTA | 50 °C for 30 sec | 489–490 |
| C1-J-1718 F | GGAGGATTTGGAAATTGATTAGTTCC | ||
| C1-n-2191R | CCCGGTAAAATTAAAATATAAACTTC | 50 °C for 20 sec | 518–574 |
| wsp 81F | TGGTCCAATAAGTGATGAAGAAAC | ||
| wsp 691R | AAAAATTAAACGCTACTCCA | 55 °C for 60 sec | 567 |
| Population | Parasitism (%) | Emergence (%) | Sex ratio |
|---|---|---|---|
| Fujian | 68.20 ± 3.24a | 86.61 ± 2.29a | 62.81% |
| Guangdong | 64.26 ± 2.86a | 85.74 ± 2.16a | 61.60% |
| Jiangxi | 63.54 ± 2.94b | 85.33 ± 2.43a | 62.14% |
| Guayas | 57.28 ± 5.00c | 82.95 ± 3.07a | 58.40% |
| Population | Samples (n) | Min. Dist. | Max. Dist. | |
|---|---|---|---|---|
| ITS1 | Fujian | 4 | 0.0020 | 0.0028 |
| Jiangxi | 4 | 0.0020 | 0.0028 | |
| Guangdong | 4 | 0.0020 | 0.0028 | |
| Overall | 12 | 0.0020 | 0.0048 | |
| ITS2 | Fujian | 4 | 0.0020 | 0.0047 |
| Jiangxi | 4 | 0.0021 | 0.0067 | |
| Guangdong | 4 | 0.0020 | 0.0043 | |
| Overall | 12 | 0.0020 | 0.0243 | |
| COI | Fujian | 4 | 0.0001 | 0.0053 |
| Jiangxi | 4 | 0.0001 | 0.0040 | |
| Guangdong | 4 | 0.0020 | 0.0040 | |
| Overall | 12 | 0.0021 | 3.0552 | |
| wsp | Fujian | 4 | 0.0022 | 0.0072 |
| Jiangxi | 4 | 0.0022 | 0.0046 | |
| Guangdong | 4 | 0.0011 | 0.0038 | |
| Overall | 12 | 0.0022 | 0.0099 |
| Gene | Nucleotide Diversity | Neutrality Tests | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Population | n | S | k | Eta | Hd | Θ | π | Fu and Li’ s D* | p-Value | Tajima’ s D | p-Value | |
| COI | Fujian | 4 | 3 | 1.50000 | 3 | 0.500 | 0.00290 | 0.00265 | −0.7544 | 0.41 | −0.7544 | 0.10 |
| Jiangxi | 4 | 2 | 1.00000 | 2 | 0.500 | 0.00211 | 0.00193 | −0.7099 | 0.10 | −0.7099 | 0.10 | |
| Guangdong | 4 | 2 | 1.16667 | 2 | 0.833 | 0.00211 | 0.00255 | 0.5915 | 0.10 | 0.5915 | 0.10 | |
| Overall | 12 | 243 | 117.848 | 243 | 0.788 | 0.16625 | 0.24349 | 1.6893 | 0.02 * | 2.1857 | 0.05 * | |
| wsp | Fujian | 4 | 3 | 1.50000 | 3 | 0.833 | 0.00353 | 0.00324 | −0.7544 | 0.10 | −0.7544 | 0.10 |
| Jiangxi | 4 | 2 | 1.16667 | 2 | 0.833 | 0.00236 | 0.00252 | 0.59158 | 0.10 | 0.5915 | 0.10 | |
| Guangdong | 4 | 3 | 1.50000 | 3 | 0.833 | 0.00353 | 0.00324 | −0.7544 | 0.10 | −0.7544 | 0.10 | |
| Overall | 12 | 6 | 1.37879 | 6 | 0.833 | 0.00429 | 0.00298 | −1.1084 | 0.10 | −1.1671 | 0.10 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ashraf, H.J.; Akutse, K.S.; Mukhtar, I.; Ramos Aguila, L.C.; Qasim, M.; Wang, W.; Bamisile, B.S.; Wang, L. Genetic Diversity of Tamarixia radiata Populations and Their Associated Endosymbiont Wolbachia Species from China. Agronomy 2021, 11, 2018. https://doi.org/10.3390/agronomy11102018
Ashraf HJ, Akutse KS, Mukhtar I, Ramos Aguila LC, Qasim M, Wang W, Bamisile BS, Wang L. Genetic Diversity of Tamarixia radiata Populations and Their Associated Endosymbiont Wolbachia Species from China. Agronomy. 2021; 11(10):2018. https://doi.org/10.3390/agronomy11102018
Chicago/Turabian StyleAshraf, Hafiza Javaria, Komivi Senyo Akutse, Irum Mukhtar, Luis Carlos Ramos Aguila, Muhammad Qasim, Wenjie Wang, Bamisope Steve Bamisile, and Liande Wang. 2021. "Genetic Diversity of Tamarixia radiata Populations and Their Associated Endosymbiont Wolbachia Species from China" Agronomy 11, no. 10: 2018. https://doi.org/10.3390/agronomy11102018