
Exogenous Glomalin-Related Soil Proteins Differentially Regulate Soil Properties in Trifoliate Orange

 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Culture
2.2. Preparation and Application of GRSP and Experimental Design
2.3. Variable Determinations
2.4. Statistical Analysis
3. Results
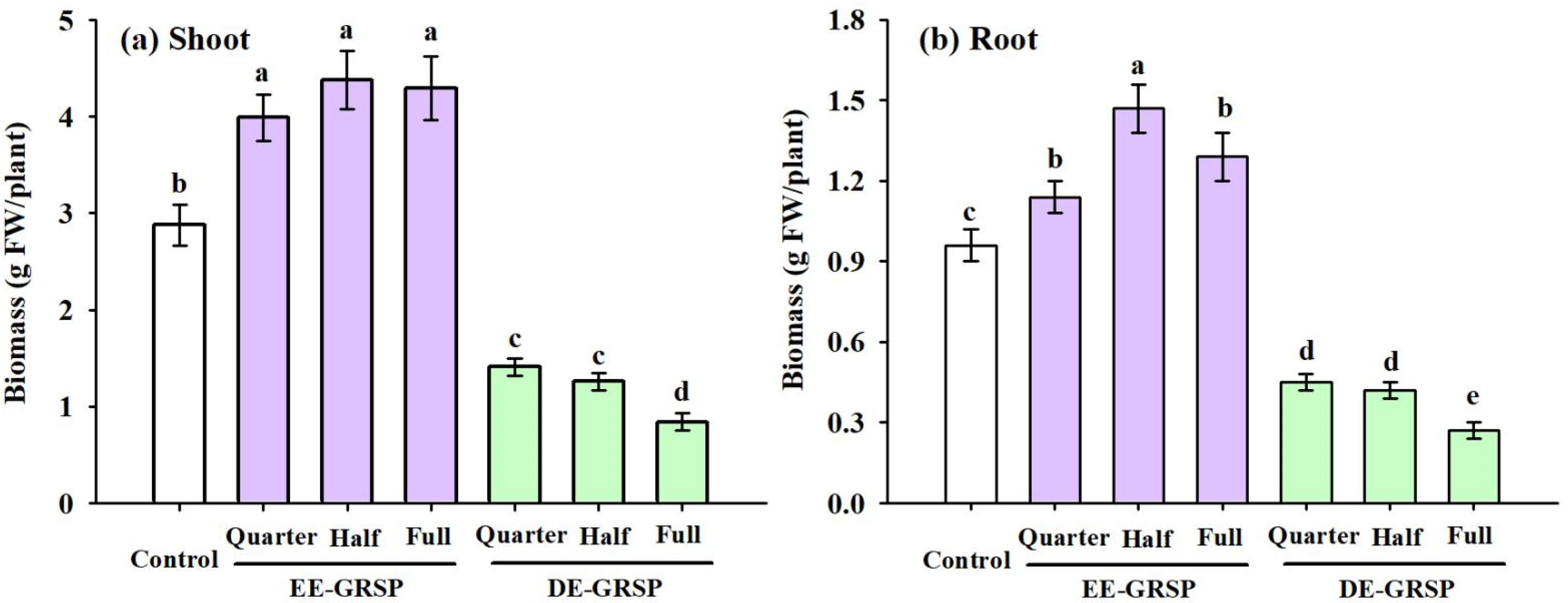
3.1. Changes in Biomass Production
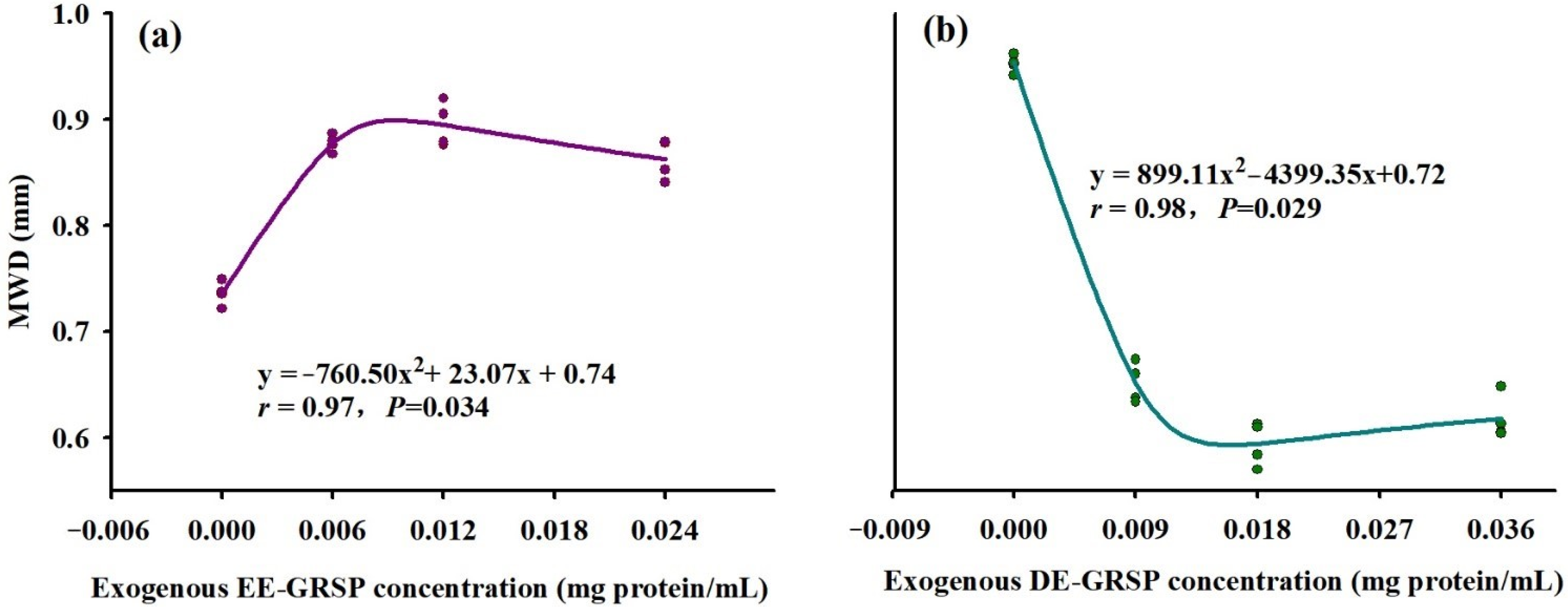
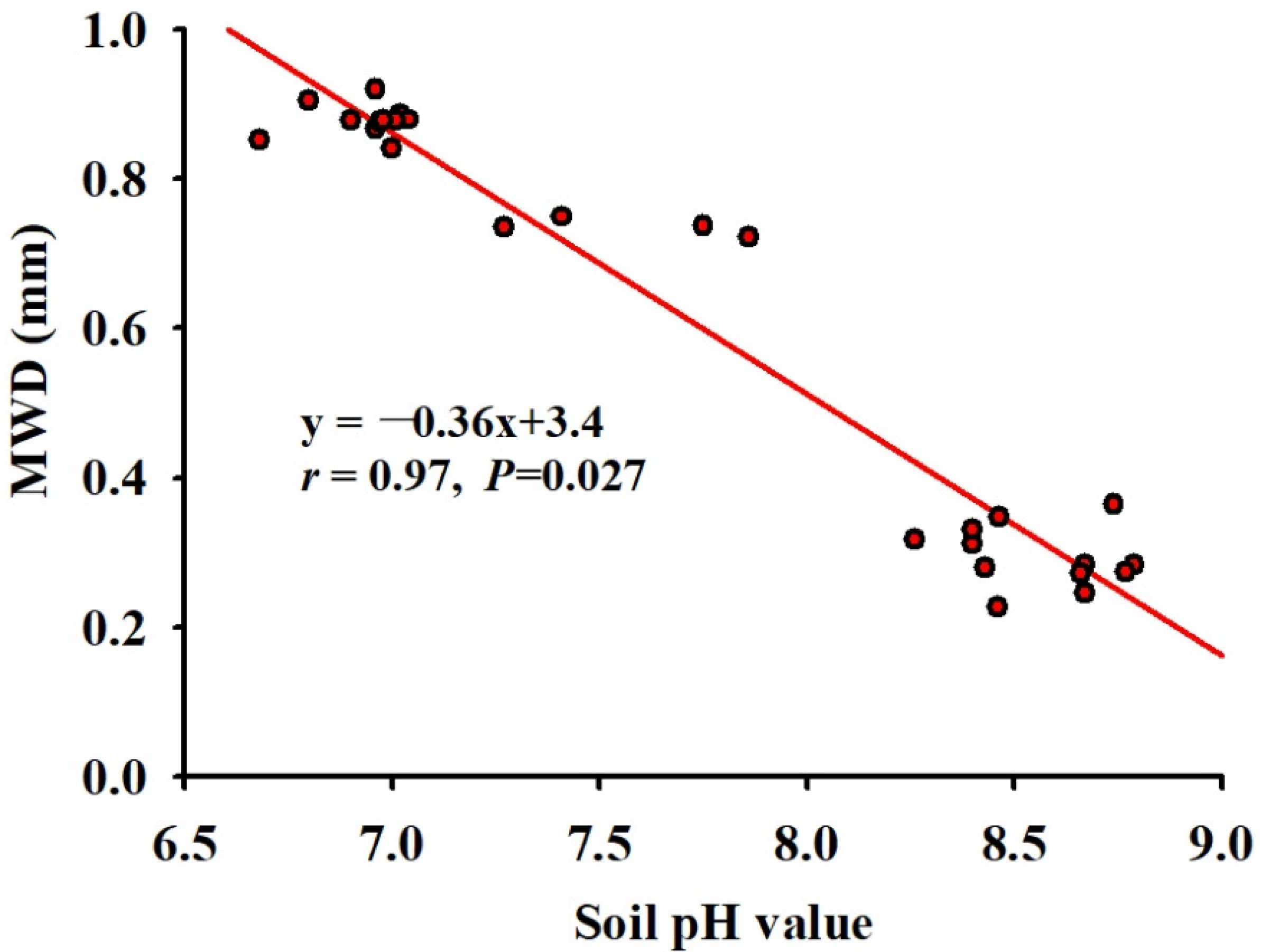
3.2. Changes in Soil WSA Distribution and Stability
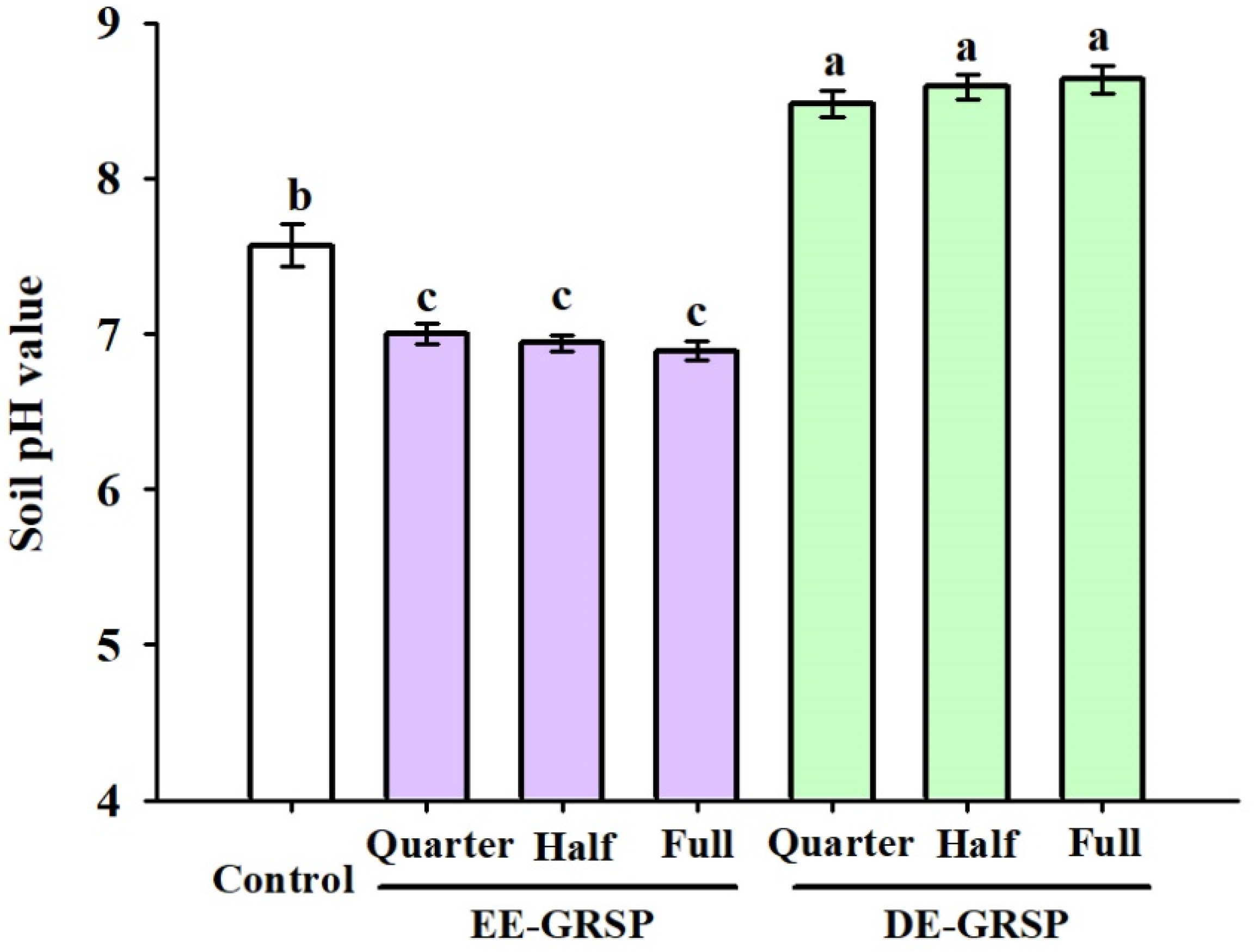
3.3. Changes in Soil pH Value
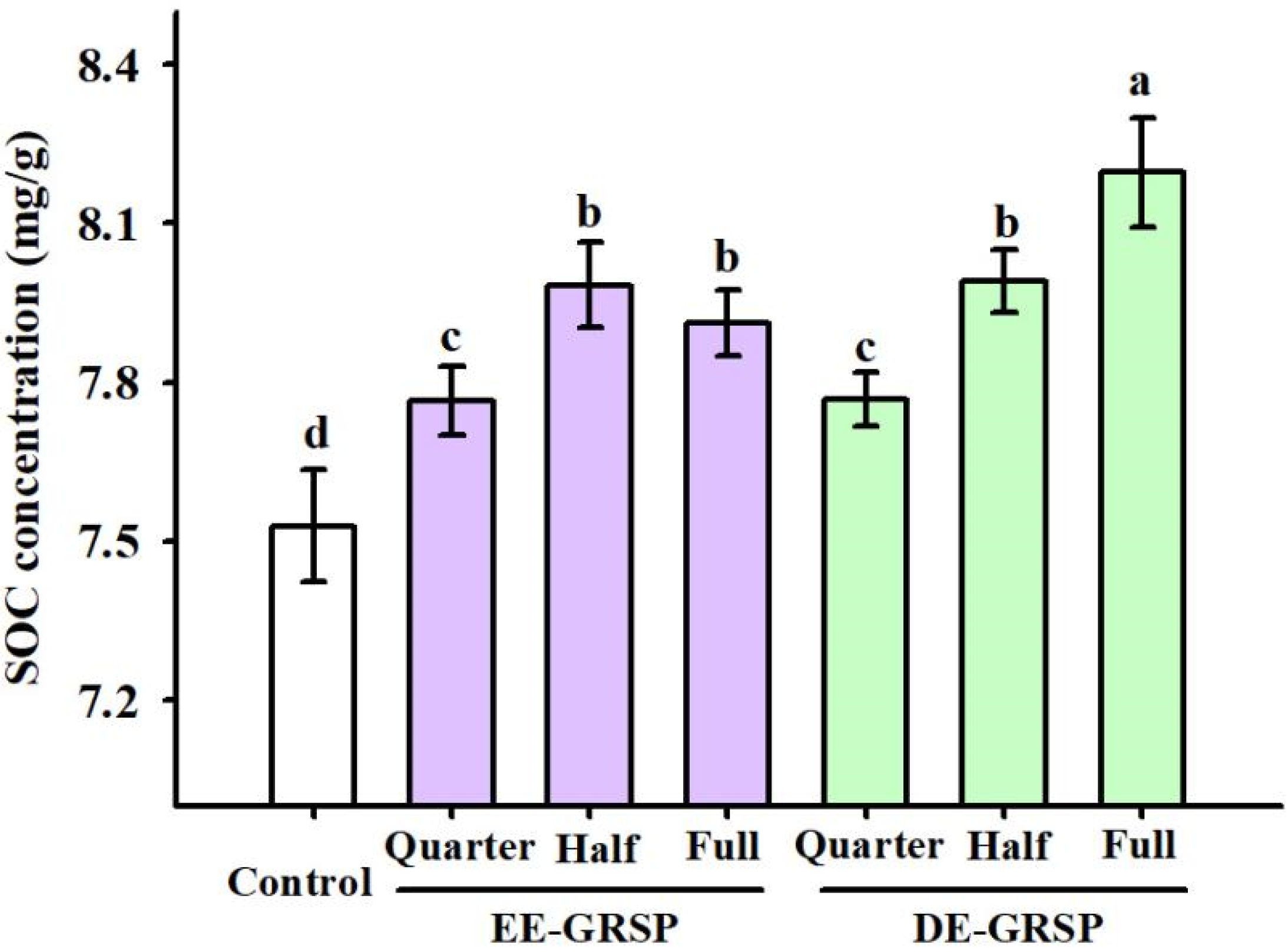
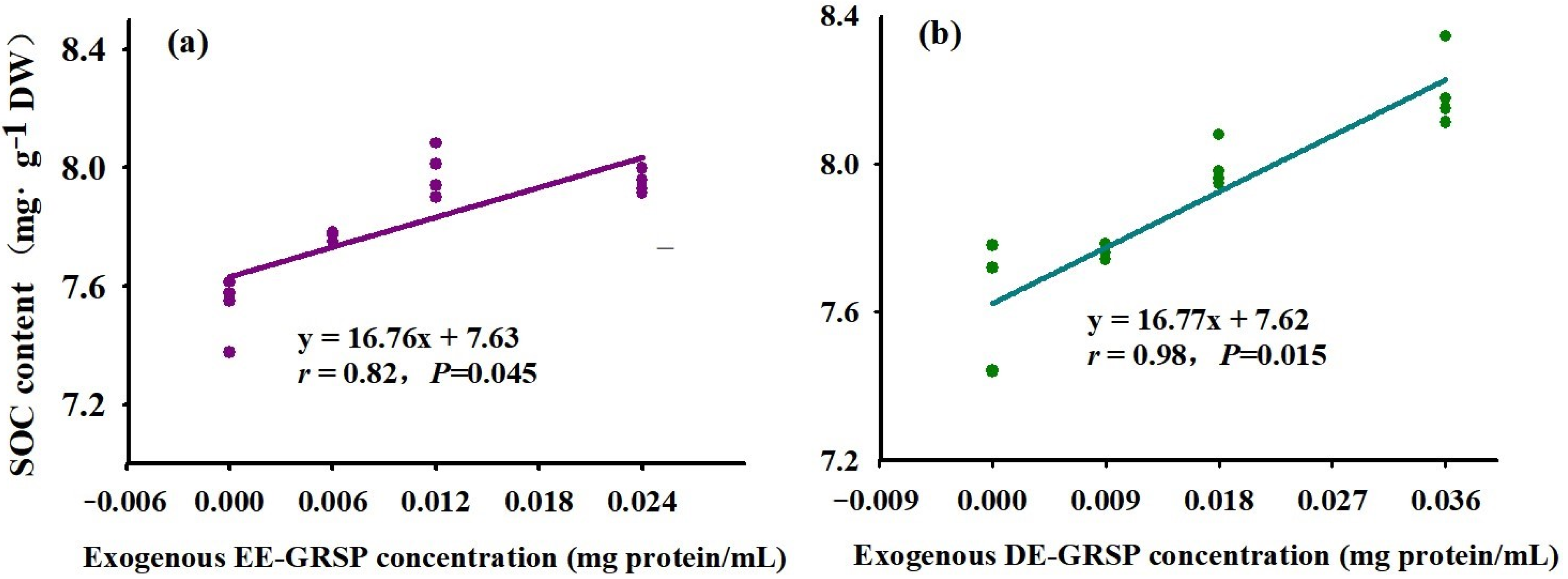
3.4. Changes in SOC Content
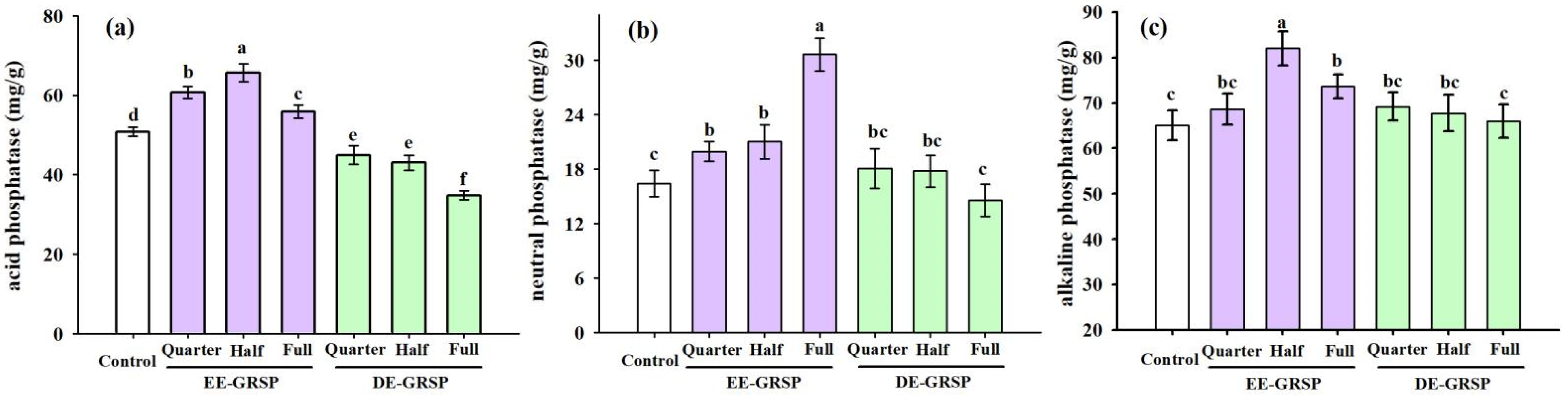
3.5. Changes in Soil Phosphatase Activity
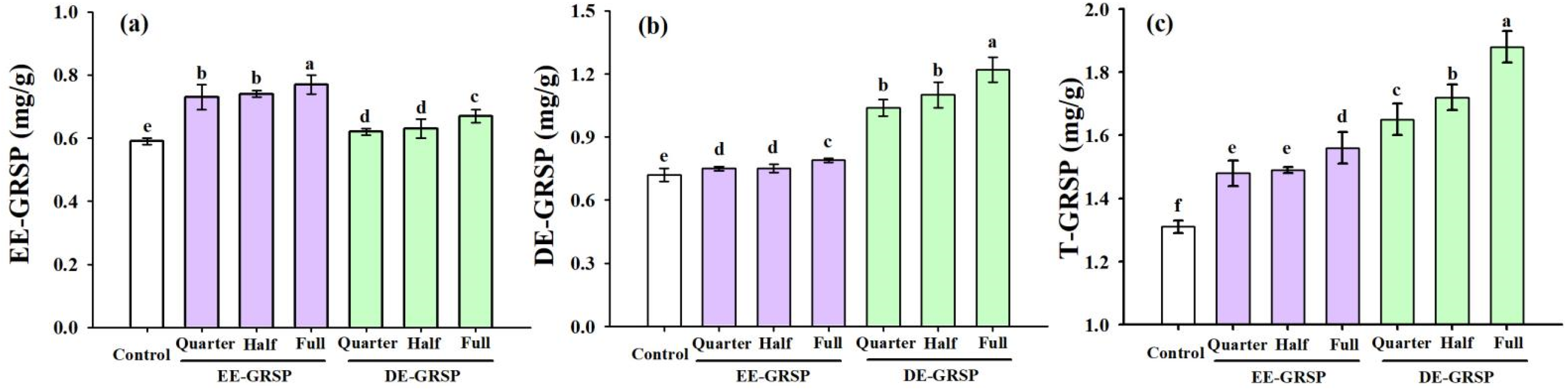
3.6. Changes in Soil GRSP Concentrations
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Walder, F.; Heijden, M.G.V.A. Regulation of resource exchange in the arbuscular mycorrhizal symbiosis. Nat. Plants 2015, 1, 15159. [Google Scholar] [CrossRef]
- Wu, Q.S.; He, J.D.; Srivastava, A.K.; Zhang, F.; Zou, Y.N. Development of propagation technique of indigenous AMF and their inoculation response in citrus. Ind. J. Agric. Sci. 2019, 89, 1190–1194. [Google Scholar]
- Gao, W.Q.; Wang, P.; Wu, Q.S. Functions and application of glomalin-related soil proteins: A review. Sains Malays. 2019, 48, 111–119. [Google Scholar] [CrossRef]
- He, J.D.; Chi, G.G.; Zou, Y.N.; Shu, B.; Wu, Q.S.; Srivastava, A.K.; Kuča, K. Contribution of glomalin-related soil proteins to soil organic carbon in trifoliate orange. Appl. Soil Ecol. 2020, 154, 103592. [Google Scholar] [CrossRef]
- Wu, Q.S.; Gao, W.Q.; Srivastava, A.K.; Zhang, F.; Zou, Y.N. Nutrient acquisition and fruit quality of Ponkan mandarin in response to AMF inoculation. Ind. J. Agric. Sci. 2020, 90, 1563–1567. [Google Scholar]
- Yang, L.; Zou, Y.N.; Tian, Z.H.; Wu, Q.S.; Kuča, K. Effects of beneficial endophytic fungal inoculants on plant growth and nutrient absorption of trifoliate orange seedlings. Sci. Hortic. 2021, 277, 109815. [Google Scholar] [CrossRef]
- Cheng, H.Q.; Giri, B.; Wu, Q.S.; Zou, Y.N.; Kuča, K. Arbuscular mycorrhizal fungi mitigate drought stress in citrus by modulating root microenvironment. Arch. Agron. Soil Sci. 2021. [Google Scholar] [CrossRef]
- Gałązka, A.; Niedźwiecki, J.; Grządziel, J.; Gawryjołek, K. Evaluation of changes in glomalin-related soil proteins (GRSP) content, microbial diversity and physical properties depending on the type of soil as the important biotic determinants of soil quality. Agronomy 2020, 10, 1279. [Google Scholar] [CrossRef]
- Ortas, I.; Çagdas, A.; Ahmet, D. Effect of mycorrhizal species on growth and nutrient uptake by seedlings of citrus (Citrus sinensis) under three soil growth conditions. Curr. Hortic. 2015, 3, 61–64. [Google Scholar]
- Ortas, I. Effect of mycorrhizal inoculation on P and Zn uptake in citrus. Curr. Hortic. 2017, 5, 59–61. [Google Scholar]
- Rillig, M.C. Arbuscular mycorrhizae, glomalin, and soil aggregation. Can. J. Soil Sci. 2004, 84, 355–363. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Wu, Q.S.; Cao, M.Q.; Zou, Y.N.; He, X.H. Direct and indirect effects of glomalin, mycorrhizal hyphae, and roots on aggregate stability in rhizosphere of trifoliate orange. Sci. Rep. 2014, 4, 5823. [Google Scholar] [CrossRef]
- Meng, L.L.; He, J.D.; Zou, Y.N.; Wu, Q.S.; Kuča, K. Mycorrhiza-released glomalin-related soil protein fractions contribute to soil total nitrogen in trifoliate orange. Plant Soil Environ. 2020, 66, 183–189. [Google Scholar] [CrossRef]
- Barna, G.; Makó, A.; Takács, T.; Füzy, A.; Horel, A.; Skic, K. Biochar alters soil physical characteristics, arbuscular mycorrhizal fungi colonization, and glomalin production. Agronomy 2020, 10, 1933. [Google Scholar] [CrossRef]
- Bertagnoli, B.; Oliveira, J.F.; Barbosa, G.M.; Filho, A.C. Poultry litter and liquid swine slurry applications stimulate glomalin, extraradicular mycelium production, and aggregation in soils. Soil Till. Res. 2020, 202, 104657. [Google Scholar] [CrossRef]
- Rillig, M.C.; Wright, S.F.; Allen, M.F.; Field, C.B. Rise in carbon dioxide changes soil structure. Nature 1999, 400, 628–629. [Google Scholar] [CrossRef]
- Wang, W.J.; Zhong, Z.L.; Wang, Q.; Wang, H.M.; Fu, Y.J.; He, X.Y. Glomalin contributed more to carbon, nutrients in deeper soils, and differently associated with climates and soil properties in vertical profiles. Sci. Rep. 2017, 7, 13003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, H.; Liu, X.L.; Gai, J.P.; Zhang, J.L.; Li, X.L. Review of glomalin-related soil protein and its function. Chin. J. Soil Sci. 2009, 40, 1215–1220. [Google Scholar]
- Gadkar, V.; Rillig, M.C. The arbuscular mycorrhizal fungal protein glomalin is a putative homolog of heat shock protein 60. FEMS Microbiol. Lett. 2006, 263, 93–101. [Google Scholar] [CrossRef] [Green Version]
- Koide, R.T.; Peoples, M.S. Behavior of Bradford-reactive substances is consistent with predictions for glomalin. Appl. Soil Ecol. 2013, 63, 8–14. [Google Scholar] [CrossRef]
- Wu, Q.S.; Li, Y.; Zou, Y.N.; He, X.H. Arbuscular mycorrhiza mediates glomalin-related soil protein production and soil enzyme activities in the rhizosphere of trifoliate orange grown under different P levels. Mycorrhiza 2015, 25, 121–130. [Google Scholar] [CrossRef]
- Wu, Q.S.; Srivastava, A.K.; Wang, S.; Zeng, J.X. Exogenous application of EE-GRSP and changes in citrus rhizosphere properties. Ind. J. Agric. Sci. 2015, 85, 58–62. [Google Scholar]
- Cheng, H.Q.; Zou, Y.N.; Wu, Q.S.; Kuča, K. Arbuscular mycorrhizal fungi alleviate drought stress in trifoliate orange by regulating H+-ATPase activity and gene expression. Front. Plant Sci. 2021, 12, 659694. [Google Scholar] [CrossRef] [PubMed]
- Kemper, W.D.; Rosenau, R.C. Aggregate Stability and Size Distribution. In Methods of Soil Analysis. Part 1. Physical and Mineralogical Methods, 2nd ed.; Klute, A., Ed.; Soil Science Society of America Agronomy Monograph No. 9; Soil Science Society of America: Madison, WI, USA, 1986; pp. 425–442. [Google Scholar]
- Rowell, D.L. Soil Science: Methods and Applications; Routledge: New York, NY, USA, 2014. [Google Scholar]
- Zhao, L.P.; Jiang, Y. Research of determination of soil phosphatase activity. Chin. J. Soil Sci. 1986, 17, 138–141. [Google Scholar]
- Wang, S.; Wu, Q.S.; He, X.H. Exogenous easily extractable glomalin-related soil protein promotes soil aggregation, relevant soil enzyme activities and plant growth in trifoliate orange. Plant Soil Environ. 2015, 2, 66–71. [Google Scholar] [CrossRef] [Green Version]
- Chi, G.G.; Srivastava, A.K.; Wu, Q.S. Exogenous easily extractable glomalin-related soil protein improves drought tolerance of trifoliate orange. Arch. Agron. Soil Sci. 2018, 64, 1341–1350. [Google Scholar] [CrossRef]
- Gillespie, A.W.; Farrell, R.E.; Walley, F.L.; Ross, A.R.S.; Leinweber, P.; Eckhardt, K.U. Glomalin-related soil protein contains non-mycorrhizal related heat-stable proteins, lipids and humic materials. Soil Biol. Biochem. 2011, 43, 766–777. [Google Scholar] [CrossRef]
- Liu, R.C.; Gao, W.Q.; Srivastava, A.K.; Zou, Y.N.; Kuča, K.; Hashem, A.; Abd_Allah, E.F.; Wu, Q.S. Differential effects of exogenous glomalin-related soil proteins on plant growth of trifoliate orange through regulating auxin changes. Front. Plant Sci. 2021, 12, 745402. [Google Scholar] [CrossRef]
- Zhang, Z.L.; Wang, W.J.; Wang, Q.; Wu, Y.; Wang, H.M.; Pei, Z.X. Glomalin amount and compositional variation, and their associations with soil properties in farmland, northeastern China. J. Plant Nutr. Soil Sci. 2017, 180, 563–575. [Google Scholar] [CrossRef]
- Schindler, F.V.; Mercer, E.J.; Rice, J.A. Chemical characteristics of glomalin-related soil protein (GRSP) extracted from soils of varying organic matter content. Soil Biol. Biochem. 2007, 39, 320–329. [Google Scholar] [CrossRef]
- Bronick, C.J.; Lal, R. Soil structure and management: A review. Geoderma 2005, 124, 3–22. [Google Scholar] [CrossRef]
- Rillig, M.C.; Wright, S.F.; Eviner, V.T. The role of arbuscular mycorrhizal fungi and glomalin in soil aggregation: Comparing effects of five plant species. Plant Soil 2002, 238, 325–333. [Google Scholar] [CrossRef]
- Zou, Y.N.; Srivastava, A.K.; Wu, Q.S. Glomalin: A potential soil conditioner for perennial fruits. Int. J. Agric. Biol. 2015, 18, 293–297. [Google Scholar] [CrossRef]
- Wu, Q.S.; Srivastava, A.K.; Cao, M.Q.; Wang, J. Mycorrhizal function on soil aggregate stability in root zone and root-free hyphae zone of trifoliate orange. Arch. Agron. Soil Sci. 2014, 61, 813–825. [Google Scholar] [CrossRef]
- Wu, Q.S.; Wang, S.; Srivastava, A.K. Mycorrhizal hyphal disruption induces changes in plant growth, glomalin-related soil protein and soil aggregation of trifoliate orange in a core system. Soil Till. Res. 2016, 160, 82–91. [Google Scholar] [CrossRef]
- Chi, C.M.; Zhao, C.W.; Sun, X.J.; Wang, Z.C. Estimating exchangeable sodium percentage from sodium adsorption ratio of salt-affected soil in the Songnen plain of northeast China. Pedosphere 2011, 21, 271–276. [Google Scholar] [CrossRef]
- Bhogal, A.; Nicholson, F.A.; Chambers, B.J. Organic carbon additions: Effects on soil bio-physical and physico-chemical properties. Eur. J. Soil Sci. 2010, 60, 276–286. [Google Scholar] [CrossRef]
- Cao, X.Y.; Li, J.P. Contents of soil organic carbon and its relations with soil physicochemical properties in Chinese fir plantations. For. Res. Manag. 2014, 6, 104–109. [Google Scholar]
- Liu, S.J.; Xia, X.; Chen, G.M.; Mao, D.; Che, S.G.; Li, Y.X. Study progress on functions and affecting factors of soil enzymes. Chin. Agric. Sci. Bull. 2011, 27, 1–7. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pH | Soil Organic Carbon (g/kg) | Alkaline N (mg/kg) | Olsen-P (mg/kg) | Available K (mg/kg) |
|---|---|---|---|---|
| 6.4 | 9.6 | 11.1 | 15.7 | 21.3 |
| Treatments | Percentage of WSA (%) | MWD (mm) | |||
|---|---|---|---|---|---|
| 2–4 mm | 1–2 mm | 0.5–1 mm | 0.25–0.5 mm | ||
| Control | 28.95 ± 0.63d | 4.28 ± 0.26c | 10.96 ± 0.59b | 17.66 ± 0.88b | 0.74 ± 0.01b |
| Quarter-strength EE-GRSP | 35.76 ± 0.16b | 3.35 ± 0.33d | 16.55 ± 1.60a | 11.36 ± 0.32c | 0.88 ± 0.01a |
| Half-strength EE-GRSP | 37.51 ± 1.02a | 7.44 ± 0.25a | 15.01 ± 1.38a | 22.57 ± 1.60a | 0.89 ± 0.02a |
| Full-strength EE-GRSP | 33.44 ± 1.03c | 5.09 ± 0.07b | 14.29 ± 0.81a | 17.04 ± 0.78b | 0.86 ± 0.02a |
| Quarter-strength DE-GRSP | 13.00 ± 1.28f | 2.38 ± 0.14e | 5.58 ± 0.23d | 9.33 ± 0.29d | 0.33 ± 0.03c |
| Half-strength DE-GRSP | 8.52 ± 1.23g | 3.33 ± 0.24d | 5.51 ± 0.74d | 10.94 ± 0.96c | 0.26 ± 0.03d |
| Full-strength DE-GRSP | 15.11 ± 0.73e | 3.07 ± 0.19d | 7.25 ± 0.64c | 10.58 ± 0.64cd | 0.29 ± 0.03cd |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, R.-C.; Zou, Y.-N.; Kuča, K.; Hashem, A.; Abd_Allah, E.F.; Wu, Q.-S. Exogenous Glomalin-Related Soil Proteins Differentially Regulate Soil Properties in Trifoliate Orange. Agronomy 2021, 11, 1896. https://doi.org/10.3390/agronomy11101896
Liu R-C, Zou Y-N, Kuča K, Hashem A, Abd_Allah EF, Wu Q-S. Exogenous Glomalin-Related Soil Proteins Differentially Regulate Soil Properties in Trifoliate Orange. Agronomy. 2021; 11(10):1896. https://doi.org/10.3390/agronomy11101896
Chicago/Turabian StyleLiu, Rui-Cheng, Ying-Ning Zou, Kamil Kuča, Abeer Hashem, Elsayed Fathi Abd_Allah, and Qiang-Sheng Wu. 2021. "Exogenous Glomalin-Related Soil Proteins Differentially Regulate Soil Properties in Trifoliate Orange" Agronomy 11, no. 10: 1896. https://doi.org/10.3390/agronomy11101896