Ranking Species for Veld Restoration in Semi-Arid Regions Using Agronomic, Morphological and Chemical Parameters of Selected Grass Species at Different Developmental Stages under Controlled Environment


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sourcing of Seeds and Planting
2.3. Data Collection
2.4. Chemical Analysis
2.5. Statistical Analysis
3. Results
4. Discussion
4.1. Plant Height and Leaf Width
4.2. Number of Tillers and Leaves per Shoot
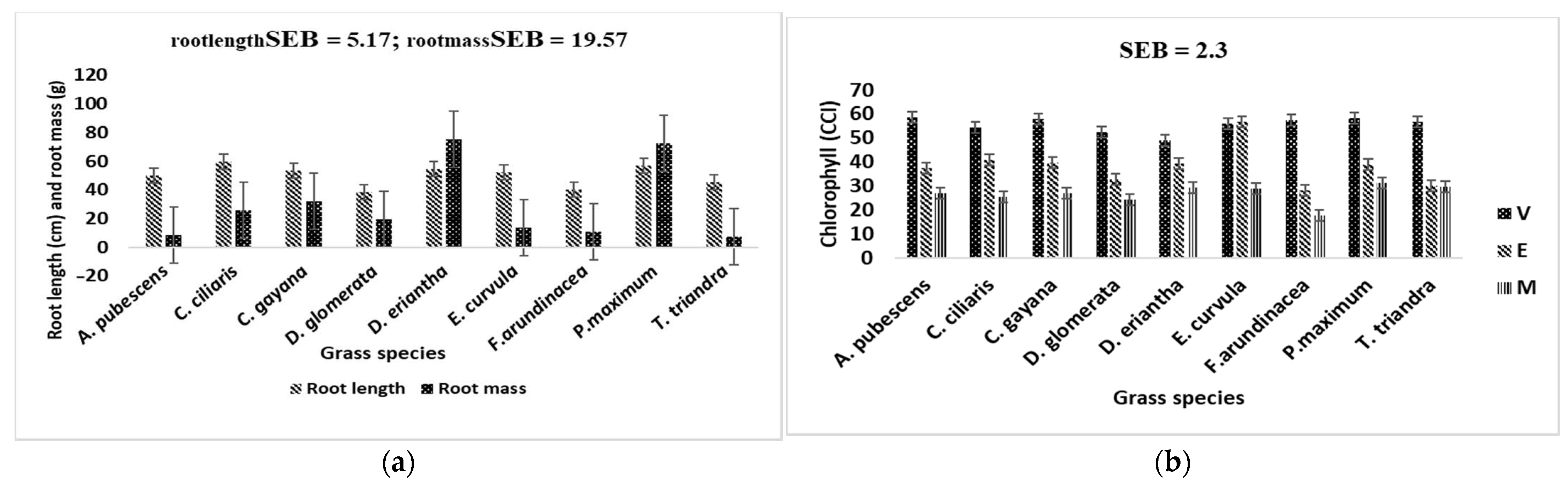
4.3. Chlorophyll
4.4. Root and Biomass Yield
4.5. Chemical Composition
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ansah, T.; Osafo, E.L.K.; Hansen, H.H. Herbage yield and chemical composition of four varieties of Napier (Pennisetum purpureum) grass harvested at three different days after planting. Agric. Biol. J. N. Am. 2010, 15, 923–929. [Google Scholar] [CrossRef]
- Beyene, S.T.; Mlambo, V. Botanical and chemical composition of common grass species around dip-tank areas in semi-arid communal rangelands of Swaziland. Trop. Subtrop. Agroecosyst. 2012, 15, 143–152. [Google Scholar]
- Deyoung, R.W.; Hellgren, E.C.; Fulbright, T.E.; Robbins, W.F.; Humphreys, I.D. Modeling nutritional carrying capacity for translocated desert Bighorn sheep in western Texas. Restor. Ecol. 2000, 8, 57–65. [Google Scholar] [CrossRef] [Green Version]
- Kassahun, A.; Tegegne, A.; Aberra, D. Impacts of rangeland degradation on soil physical, chemical and seed bank properties along a gradient in three rangeland vegetation types in Somali Region, Eastern Ethiopia. Ethiop. J. Agric. Sci. 2012, 22, 84–101. [Google Scholar]
- Selemani, I.S.; Eik, L.O.; Holand, Ø.; Ådnøy, T.; Mtengeti, E.; Mushi, D. The effects of a deferred grazing system on rangeland vegetation in a north-western, semi-arid region of Tanzania. Afr. J. Range Forage Sci. 2013, 30, 141–148. [Google Scholar] [CrossRef]
- Lascano, C.E.; Schmidt, A.; Barahona, R. Forage quality and the environment. Int. Grassl. Congr. 2001, 19, 1–19. [Google Scholar]
- Zimmermann, J.; Higgins, S.I.; Grimm, V.; Hoffmann, J.; Münkemüller, T.; Linstädter, A. Recruitment filters in a perennial grassland: The interactive roles of fire, competitors, moisture and seed availability. J. Ecol. 2008, 96, 1033–1044. [Google Scholar] [CrossRef]
- Koukolová, V.; Homolka, P.; Koukol, O.; Jančík, F. Nutritive value of Trifolium pratense L. for ruminants estimated from in situ ruminal degradation of neutral detergent fibre and in vivo digestibility of organic matter and energy. Czech J. Anim. Sci. 2010, 55, 372–381. [Google Scholar] [CrossRef] [Green Version]
- Gustavsson, A.M. A developmental scale for perennial forage grasses based on the decimal code framework. Grass Forage Sci. 2011, 66, 93–108. [Google Scholar] [CrossRef]
- Ravhuhali, K.E.; Motsepe, K.; Mnisi, C.M.; Sebolai, T. Chemical composition and in vitro ruminal DM degradability of native pasture grasses and their plant parts. Range Manag. Agrofor. 2018, 39, 243–250. [Google Scholar]
- Arzani, H.; Basiri, M.; Khatibi, F.; Ghorbani, G. Nutritive value of some Zagros Mountain rangeland species. Small Rumin. Res. 2006, 65, 128–135. [Google Scholar] [CrossRef]
- Elberse, W.T.; Berendse, F. A comparative study of the growth and morphology of eight grass species from habitats with different nutrient availabilities. Funct. Ecol. 1993, 7, 223–229. [Google Scholar] [CrossRef]
- Tefera, S. Local knowledge of grasses in semi-arid South Africa: Comparison of forage traits, status and trends, and similarities with field studies. In Proceedings of the Grassland Society of Southern Africa 49th Annual Congress, Wilderness, Western Cape, South Africa, 3–8 July 2016. [Google Scholar]
- Ravhuhali, K.E.; Mlambo, V.; Beyene, T.S.; Palamuleni, L.G. A comparative analysis of the morphology and nutritive value of five South African native grass species grown under controlled conditions. Afr. J. Range Forage Sci. 2019, 36, 67–70. [Google Scholar] [CrossRef]
- Aamlid, T.S.; Kvalbein, A.; Waalen, W. Handbook Turf Grass Winter Survival: Grass Species and Varieties for Severe Winter Climates; NIBIO Turfgrass Research Group, Norwegian Institute of Bioeconomy Research: Oslo, Norway, 2020. [Google Scholar]
- AOAC. Official Methods of Analysis of AOAC International, 18th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 2012. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- SAS. Statistical Analysis System. Statistics Software. Release 10; SAS Institute Inc.: Cary, NC, USA, 2010. [Google Scholar]
- Ozkose, A.; Tamkoc, A. Morphological and agronomic characteristics of perennial rye grass (Lolium perenne L.) genotype. Turk. J. Field Crops 2014, 19, 231–237. [Google Scholar] [CrossRef] [Green Version]
- Falster, D.S.; Westoby, M. Plant height and evolutionary games. Trends Ecol. Evol. 2003, 18, 337–343. [Google Scholar] [CrossRef]
- Moles, A.T.; Warton, D.I.; Warman, L.; Swenson, N.G.; Laffan, S.W.; Zanne, A.E.; Pitman, A.; Hemmings, F.A.; Leishman, M.R. Global patterns in plant height. J. Ecol. 2009, 97, 923–932. [Google Scholar] [CrossRef]
- Zhouri, L.; Kallida, R.; Shaimi, N.; Barre, P.; Volaire, F.; Gaboun, F.; Fakiri, M. Evaluation of cocksfoot (Dactylis glomerata L.) population for drought survival and behaviour. Saudi J. Biol. Sci. 2019, 26, 49–56. [Google Scholar] [CrossRef]
- Vasileva, V.; Kocheva, K.; Mincheva, J.; Georgiev, G.; Ilieva, A.; Porqueddu, C. Physiological analysis of growth and nitrogen metabolism of intercropped pasture species subterranean clover (Trifolium subterraneum L.) and cocksfoot (Dactylis glomerata L.) supplemented with different forms of inorganic nitrogen. J. Plant Nutr. 2017, 40, 2116–2126. [Google Scholar] [CrossRef]
- Van Oudtshoorn, F. Guide to Grasses of Southern Africa; Briza Publications: Pretoria, South Africa, 2014. [Google Scholar]
- Ojo, V.O.A.; Dele, T.A.; Amole, U.Y.; Adeoye, S.A.; Hassan, J.A.O.; Idowu, O.J. Effect of intercropping Panicum Maximum var. Ntchisi and Lablab purpureus on the growth, yield and chemical composition of Panicum maximum var. Ntchisi at different harvesting times. Pak. J. Biol. Sci. 2013, 10, 1–4. [Google Scholar] [CrossRef]
- Yigzaw, G.W. Effect of harvesting stage on yield and nutritive value of buffel grass (Cenchrus ciliaris linn) under irrigation at Gewane district, north eastern Ethiopia. J. Sci. Innov. Res. 2019, 8, 7–12. [Google Scholar]
- Werner, J.; Umstatter, C.; Leso, L.; Kennedy, E.; Geoghegan, A.; Shalloo, L.; Schick, M.; O’Brien, B. Evaluation and application potential of an accelerometer-based collar device for measuring grazing behaviour of dairy cows. Animal 2019, 13, 2070–2079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urrea, J.L. Grass Height in Cow Nutrition: Size does Matter; CIAT Blog: Medellín, Colobmbia, 2019. [Google Scholar]
- Olson, A.B.E.; Richards, J.H. Tussock regrowth after grazing intercalary: Meristem and axillary bud activity of tillers of Agropyron desertorum. Oikos 1988, 51, 374–382. [Google Scholar] [CrossRef]
- Manske, L.L. General Description of Grass Growth and Development and Defoliation Resistance Mechanisms. NDSU Dickson Research Extension Center. Range Management Report DREC 98-1022; NDSU Dickinson Research Extension Center: Dickinson, ND, USA, 1998. [Google Scholar]
- Trlica, M.J. Grass Growth and Response to Grazing. Service Action; No. 6.108; Colorado State University: Fort Collins, CO, USA, 1992. [Google Scholar]
- Bhattarai, S.P.; Fox, J.; Gyasi-Agyei, Y. Enhancing buffel grass seed germination by acid treatment for rapid vegetation establishment on railway batters. J. Arid Environ. 2008, 72, 255–262. [Google Scholar] [CrossRef]
- Laidlaw, A.S. The relationship between tiller appearance in spring and contribution to dry-matter yield in perennial ryegrass (Lolium perenne L.) cultivars differing in heading date. Grass Forage Sci. 2005, 60, 200–209. [Google Scholar] [CrossRef]
- Mganga, K.Z.; Musimba, N.K.; Nyariki, D.M.; Nyangito, M.; Mwang’omb, A.; Ekaya, W.; Mwang’ombe, A.; Muiru, W.; Books, R.; Oer, R.; et al. Influence of forage value on the choice of grass species to combat desertification in semi-arid regions of Kenya. In Proceedings of the Fifth African Higher Education week and RUFORUM Biennial Conference, “Linking Agricultural Universities with Civil Society, the Private Sector, Governments and Other Stakeholders in Support of Agricultural Development in Africa”, Cape Town, South Africa, 17–21 October 2016; pp. 537–541. [Google Scholar]
- Montagner, D.B.; Nascimento Júnior, D.D.; Vilela, H.H.; Sousa, B.M.D.L.; Euclides, V.P.B.; Silva, S.C.D.; Carloto, M.N. Tillering dynamics in pastures of guinea grass subjected to grazing severities under intermittent stocking. Rev. Bras. Zootec. 2012, 41, 544–549. [Google Scholar] [CrossRef] [Green Version]
- Nguku, S.A. An Evaluation of Brachiaria Grass Cultivars Productivity in Semi-Arid Kenya. Master’s Thesis, South Eastern Kenya University, Kitui, Kenya, 2015. [Google Scholar]
- De Lima Veras, E.L.; Difante, G.D.S.; Chaves Gurgel, A.L.; Gracianoda Costa, A.B.; Gomes Rodrigues, J.; Marques Costa, C.; Emerenciano Neto, J.V.; Gusmão Pereira, M.D.; Ramon Costa, P. Tillering and structural characteristics of panicum cultivars in the Brazilian semiarid region. Sustainability 2020, 12, 3849. [Google Scholar] [CrossRef]
- Moolman, A.C.; Van Rooyen, N.; Van Rooyen, M.W. The effect of drought stress on the dry matter production, growth rate and biomass allocation of Anthephora pubescens Nees. S. Afr. J. Bot. 1996, 62, 41–45. [Google Scholar] [CrossRef] [Green Version]
- Opiyo, F.E.O.; Ekaya, W.N.; Nyariki, D.M.; Mureithi, S.M. Seedbed preparation influence on morphometric characteristics of perennial grasses of a semi-arid rangeland in Kenya. Afr. J. Plant Sci. 2011, 5, 460–468. [Google Scholar]
- Mnene, W.N. Strategies to Increase Success Rates in Natural Pasture Improvement through Re-Seeding Degraded Semi-Arid Rangelands of Kenya. Ph.D. Thesis, University of Nairobi, Nairobi, Kenya, 2006. [Google Scholar]
- Arzani, H.; Zohdi, M.; Fish, E.; Amiri, G.H.Z.; Nikkhah, A.; Wester, D. Phenological effects on forage quality of five grass species. J Range Manag. 2004, 57, 624–629. [Google Scholar] [CrossRef]
- Hughes, M.P.; Mlambo, V.; Lallo, C.H.O.; Basha, N.A.D.; Nsahlai, I.V.; Jennings, P.G.A. Accuracy of two optical chlorophyll meters in predicting chemical composition and in vitro ruminal organic matter degradability of Brachiaria hybrid, Megathyrsus maximus, and Paspalum atratum. Anim. Nutr. 2017, 3, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.S.; Mishra, K.N.; Mahananda, M.R. Chlorophyll content studies from inception of leaf buds to leaf-fall stages of Teak (Tectona Grandis) of Kapilash forest division, Dhenkanal, Odisha. J. Glob. Biosci. 2013, 2, 26–30. [Google Scholar]
- Filimon, R.V.; Rotaru, L.; Filimon, R.M. Quantitative investigation of leaf photosynthetic pigments during annual biological cycle of Vitis vinifera L. table grape cultivars. S. Afr. J. Enol. Vitic. 2016, 37, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Biber, P.D. Evaluating a chlorophyll content meter on three coastal wetland plant species. J. Agric. Food Environ. Sci. 2007, 1, 1–11. [Google Scholar]
- Brown, R. The water relations of range plants: Adaptations to water deficits. Wildl. Plants Physiol. Ecol. Dev. Morphol. 1995, 47, 291–413. [Google Scholar]
- Adjolohoun, S.; Bindelle, J.; Adandedjan, C.; Colinet, G.; Buldgen, A. Soil chemical changes following 3-year legume or grass leys in west Africa. Trop. Grasslands 2010, 44, 115–122. [Google Scholar]
- Demlew, M.; Alemu, B.; Awuk, A. Evaluation of agronomic performance and biomass yield of buffel grass and silver leaf Desmodium grown in pure stands and in mixture at different harvesting times in Gozamen district, east Gojjam Zone, Ethiopia. Greener J. Agric. Sci. 2019, 9, 396–404. [Google Scholar]
- Ndathi, A.; Bosma, L.; Kioko, T.; Kadenyi, N.; Wambua, S.; Van Steenbergen, F.; Musimba, N. Morpho-ecological characteristics of forage grasses used to rehabilitate degraded African rangelands. Rangel. J. 2019, 1–19. [Google Scholar]
- Marshall, V.M.; Lewis, M.M.; Ostendorf, B. Buffel grass (Cenchrus ciliaris) as an invader and threat to biodiversity in arid environments: A review. J. Arid Environ. 2012, 78, 1–12. [Google Scholar] [CrossRef]
- Tedder, M.J.; Morris, C.D.; Fynn, R.W.S.; Kirkman, K.P. Grass-on-grass competition along a catenal gradient in mesic grassland, South Africa. Afr. J. Range Forage Sci. 2011, 28, 79–85. [Google Scholar] [CrossRef]
- Rambau, M.D.; Fushai, F.; Baloyi, J.J. Productivity, chemical composition and ruminal degradability of irrigated Napier grass leaves harvested at three stages of maturity. S. Afr. J. Anim. Sci. 2016, 46, 388–408. [Google Scholar] [CrossRef] [Green Version]
- Odedire, J.A.; Babayemi, O.J. Comparative studies on the yield and chemical composition of Panicum maximum and Andropogon gayanus as influenced by Tephrosia candida and Leucaena leucocephala. Livest. Res. Rural Dev. 2008, 20, 1–8. [Google Scholar]
- Wassie, W.A.; Tsegay, B.A.; Wolde, A.T.; Limeneh, B.A. Evaluation of morphological characteristics, yield and nutritive value of Brachiaria grass ecotypes in Northwestern Ethiopia. Agric. Food Secur. 2018, 7, 89. [Google Scholar] [CrossRef]
- Daba, A.W.; Qureshi, A.S.; Nisaren, B.N. Evaluation of some Rhodes grass (Chloris gayana) genotypes for their salt tolerance, biomass yield and nutrient composition. Appl. Sci. 2019, 9, 143. [Google Scholar] [CrossRef] [Green Version]
- Soliva, C.R.; Amelchanka, S.L.; Kreuzer, M. The requirements for rumen-degradable protein per unit of fermentable organic matter differ between fibrous feed sources. Front. Microbiol. 2015, 6, 1–17. [Google Scholar] [CrossRef]
- Keba, H.T.; Madakadze, I.C.; Angassa, A.; Hassen, A. Nutritive value of grasses in semi-arid rangelands of Ethiopia: Local experience-based herbage preference evaluation versus laboratory analysis. Asian Australas. J. Anim. Sci. 2013, 26, 366–377. [Google Scholar] [CrossRef] [Green Version]
- Berhane, G.; Eik, L.O.; Tolera, A. Chemical composition and in vitro gas production of vetch (Vicia sativa) and some browse and grass species in northern Ethiopia. Afr. J. Range Forage Sci. 2006, 23, 69–75. [Google Scholar] [CrossRef]
- Erasmus, L.J.; Botha, P.M.; Cruywagen, C.W.; Meissner, H.H. Amino acid profile and intestinal digestibility in dairy cows of rumen-undegradable protein from various feedstuffs. J. Dairy Sci. 1994, 77, 541–551. [Google Scholar] [CrossRef]
- Adejoro, F.A.; Hassen, A. Effect of supplementing or treating Eragrostis curvula hay with urea or nitrate on its digestibility and in vitro fermentation. S. Afr. J. Anim. Sci. 2017, 47, 168–177. [Google Scholar] [CrossRef] [Green Version]
- Omotoso, O.B.; Ogunlusi, O.K.; Fajemisin, A.N.; Alokan, J.A. Growth response of west African dwarf sheep fed guinea grass substituted with Mulberry. In Proceedings of the 10th International Rangeland Congress, Saskatoon, SK, Canada, 16–22 July 2016; pp. 341–343. [Google Scholar]
- National Research Council (NRC). Nutrient Requirements of Laboratory Animals: 1995; National Academies Press: Washington, DC, USA, 1995; pp. 38–53. [Google Scholar]
- National Research Council (NRC). Nutrient Requirements of Beef Cattle, 7th ed.; NRC: Washington, DC, USA, 2000. [Google Scholar]
- Amary, N.M. Assessing the Quality of Forage for Livestock in a Semi-Arid Pastoral System in South Africa. Master’s Thesis, University of the Western Cape, Tigerberg City, South Africa, 2016. [Google Scholar]
- Amiri, F.; Mohamed Shariff, A.R. Comparison of nutritive values of grasses and legume species using forage quality index. Songklanakarin J. Sci. Technol. 2012, 34, 577–586. [Google Scholar]
- Lima, L.G.D.; Nussio, L.G.; Gonçalves, J.R.S.; Simas, J.M.C.D.; Pires, A.V.; Santos, F.A.P. Frontes de amido e proteína em dietas para vacas leiteiras em dietas à base de capim elefante. Sci. Agric. 2002, 59, 19–27. [Google Scholar] [CrossRef] [Green Version]
- Schroeder, K. Feeding Cull Potatoes to Dairy and Beef Cattle; University of Wisconsin Extension: Portage Country, WI, USA, 2012; Available online: http://citeseerx.ist.psu.edu/viewdoc/download?doi=10.1.1.278.5494&rep=rep1&type=pdf (accessed on 28 December 2020).
- Matlebyane, M.M.; Ng’ambi, J.W.W.; Aregheore, E.M. Relationships between chemical composition and in vitro digestibility of some common forage species used for ruminant livestock production in three chief areas of Capricorn region, Limpopo Province, South Africa. Res. J. Agric. Biol. Sci. 2009, 5, 138–149. [Google Scholar]
- Van Niekerk, W.A.; Hassen, A. Qualitative evaluation of four subtropical grasses as standing hay: Diet selection, rumen fermentation and partial digestibility by sheep. Afr. J. Range Forage Sci. 2009, 26, 69–74. [Google Scholar] [CrossRef]
- Coêlho, J.J.; de Mello, A.C.L.; dos Santos, M.V.F.; Dubeux, J.C.B., Jr.; da Cunha, M.V.; Lira, M.A. Prediction of the nutritional value of grass species in the semiarid region by repeatability analysis. Pesqui. Agropecu. Bras. 2018, 53, 378–385. [Google Scholar] [CrossRef] [Green Version]
- Galindo, F.S.; Buzetti, S.; Filho, M.C.M.T.; Dupas, E. Rates and sources of nitrogen fertilizer application on yield and quality of Panicum maximum cv. Mombasa. Idesia 2019, 37, 67–73. [Google Scholar] [CrossRef]
- Abdullah, M.; Rafay, M.; Hussain, T.; Ahmad, H.; Tahir, U.; Rasheed, F.; Ruby, T.; Khalil, S. Nutritive potential and palatability preference of browse foliage by livestock in arid rangelands of Cholistan desert (Pakistan). J. Anim. Plant Sci. 2017, 27, 1656–1664. [Google Scholar]
- Aung, M.; Thein, S.M.; Phaw, N.C.; Phyo, T.H.; Phyo, Y.P.; Phyo, N.Y.; Van Bik, L.; Bo, A.B.; Maw, H.; Maw, A.M.; et al. In vitro fermentation of grass-based diet supplemented with two different tree legume forages in ruminant. Adv. Anim. Vet. Sci. 2019, 10, 272–279. [Google Scholar] [CrossRef] [Green Version]
- Ndozi, H.M. Effects of Chemical Treatment on Nutrient Content and Palatability of Senegalia mellifera and Catophtactes alexandrii Bush-Based Feeds. Master’s Thesis, University of Namibia, Windhoek, Namibia, 2019. [Google Scholar]
- Limenih, B.A. Evaluation of the Agronomic, Utilization, Nutritive and Feeding Value of desho Grass (Pennisetum pedicellatum). Ph.D. Thesis, Jimma University, Jimma, Ethiopia, 2016. [Google Scholar]
- Agza, B.; Kassa, B.; Zewdu, S.; Aklilu, E.; Alemu, F. Forage yield and nutritive value of natural pastures at varying levels of maturity in North West Lowlands of Ethiopia. World J. Agric. Sci. 2013, 1, 106–112. [Google Scholar]
- Ramírez, G.R.; Aguilera-Gonzalez, J.C.; Garcia-Diaz, G.; Núñez-González, A.M. Effect of urea treatment on chemical composition and digestion of Cenchrus ciliaris and Cynodon dactylon hays and Zea mays residues. Cellulose 2007, 38, 1036–1041. [Google Scholar]
- Aganga, A.A.; Autlwetse, M.N. Utilization of sorghum forage, millet forage, veldt grass and buffel grass by Tswana sheep and goats when fed Lablab purpureus L. as protein supplement. Asian Australas. J. Anim. Sci. 2000, 13, 1127–1132. [Google Scholar] [CrossRef]
- Bayble, T.; Melaku, S.; Prasad, N.K. Effects of cutting dates on nutritive value of Napier (Pennisetum purpureum) grass planted sole and in association with Desmodium (Desmodium intortum) or Lablab (Lablab purpureus). Livest. Res. Rural Dev. 2007, 19, 120–136. [Google Scholar]



{kind=link}
{kind=link}
| Plant Height (cm) | Leaf Width (mm) | |||||
|---|---|---|---|---|---|---|
| Grass Species | V | E | M | V | E | M |
| A. pubescens | 25.60 aC | 47.33 cB | 113.21 bA | 4.98 cC | 6.37 cB | 8.04 cdA |
| C. ciliaris | 34.52 aC | 76.37 aB | 107.28 bcA | 3.95 cdC | 7.86 bcB | 12.54 bA |
| C. gayana | 25.61 aC | 54.94 bcB | 89.77 eA | 5.55 bcC | 7.40 bcB | 8.49 cA |
| D. glomerata | 23.96 aC | 36.78 cB | 57.10 fA | 3.84 cdeC | 6.01 cB | 7.46 cdA |
| D. eriantha | 28.53 aC | 52.18 bcB | 118.00 abA | 7.04 bB | 8.97 bB | 12.55 bA |
| E. curvula | 37.91 aC | 69.96 abB | 102.99 bcdA | 1.89 eC | 3.65 dB | 4.76 eA |
| F. arundinacea | 25.07 aC | 40.91 cdB | 84.69 deA | 2.92 deC | 6.43 cB | 7.90 cdA |
| P. maximum | 27.48 aC | 51.30 cB | 80.35 eA | 12.53 aC | 15.27 aB | 17.88 aA |
| T. triandra | 29.77 aC | 70.36 abB | 135.59 aA | 4.02 cdeB | 6.11 cA | 6.27 deA |
| SE | 3.508 | 0.371 | ||||
| Average Number of Tillers (log10) | Average Number of Leaves (LN/shoot) (log10) | |||||
|---|---|---|---|---|---|---|
| Grass Species | 0–2 Months | 2–4 Month | 4–6 Months | 0–2 Months | 2–4 Months | 4–6 Months |
| A. pubescens | 0.30 abB | 0.81 abA | 0.87 cdeA | 0.32 abC | 0.60 aB | 0.92 bcdA |
| C. ciliaris | 0.25 abC | 0.65 cB | 0.80 eA | 0.28 bcC | 0.58 aB | 0.98 bA |
| C. gayana | 0.29 abC | 0.68 cB | 1.15 abA | 0.30 abcC | 0.63 aB | 1.11 aA |
| D. glomerata | 0.21 bC | 0.65 cB | 0.92 cdA | 0.37 aC | 0.62 aB | 0.92 bcdA |
| D. eriantha | 0.30 aC | 0.48 dB | 0.85 deA | 0.23 cC | 0.60 aB | 0.90 cdA |
| E. curvula | 0.24 abC | 0.48 dB | 1.07 bA | 0.300 abcC | 0.58 aB | 0.88 dA |
| F. arundinacea | 0.27 abB | 0.90 aA | 0.95 cA | 0.27 bcC | 0.59 aB | 0.95 bcA |
| P. maximum | 0.30 abC | 0.80 bB | 1.20 aA | 0.300 abcC | 0.61 aB | 0.92 bcdA |
| T. triandra | 0.27 abC | 0.56 dB | 0.79 eA | 0.27 bcC | 0.58 aB | 0.99 bA |
| SE | 0.03 | 0.35 | ||||
| Ash (g/kg DM) | OM (g/kg DM) | CP (g/kg DM) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Grass Species | V | E | M | V | E | M | V | E | M |
| A. pubescens | 116.68 dA | 107.45 eB | 78.27 fC | 836.41 bA | 835.11 cA | 849.88 bcA | 55.39 eA | 51.63 bcA | 40.68 bcB |
| C. ciliaris | 127.58 bA | 130.03 bA | 131.41 bA | 827.10 bA | 830.38 cA | 832.59 deA | 87.27 cA | 35.01 dB | 30.32 dB |
| C. gayana | 119.05 dB | 132.69 bA | 120.66 cB | 824.22 bB | 822.68 cB | 842.22 bcdA | 87.63 cA | 40.00 dB | 37.10 bcdB |
| D. glomerata | 121.88 cdA | 115.28 cdB | 123.56 cA | 822.50 bA | 835.15 cA | 836.15 cdeA | 75.95 dA | 61.38 abB | 59.18 aB |
| D. eriantha | 125.64 bcA | 89.02 fC | 113.39 dB | 821.53 bC | 855.91 bB | 877.59 aA | 64.52 eA | 42.12 cdB | 39.01 bcdB |
| E. curvula | 90.83 eA | 75.24 gC | 83.69 fB | 864.60 aB | 890.98 aA | 881.76 aA | 102.29 bA | 65.01 aB | 58.12 aB |
| F. arundinacea | 142.76 aA | 111.37 deC | 118.13 cdB | 757.12 cB | 825.52 cA | 828.11 deA | 120.14 aA | 60.31 abB | 55.19 aB |
| P. maximum | 130.60 bC | 143.72 aB | 148.92 aA | 828.09 bA | 826.80 cA | 821.24 eA | 97.07 bcA | 56.41 abB | 45.07 bC |
| T. triandra | 116.48 dA | 117.40 cA | 106.41 eB | 838.03 bB | 838.24 cB | 855.87 bA | 35.79 fA | 34.84 dA | 33.27 cdA |
| SE | 1.92 | 5.89 | 3.49 | ||||||
| NDF | ADF | ADL | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Grass Species | V | E | M | V | E | M | V | E | M |
| A. pubescens | 553.71 cB | 667.44 aA | 687.94 aA | 418.04 aB | 450.75 aA | 449.17 abA | 131.59 aB | 164.43 aA | 191.56 bA |
| C. ciliaris | 581.03 bcB | 665.57 aA | 678.43 abA | 390.62 bC | 446.99 aB | 471.09 aA | 133.98 aB | 163.06 aB | 217.21 abA |
| C. gayana | 557.14 cB | 599.89 bA | 638.85 bcA | 324.68 cB | 338.00 dB | 384.34 dA | 168.07 aB | 185.62 aA | 206.06 abA |
| D. glomerata | 470.27 dA | 483.37 dA | 493.82 dA | 325.41 cB | 363.17 cA | 375.63 dA | 167.66 aA | 177.00 aA | 201.35 bA |
| D. eriantha | 620.97 abB | 644.78 aA | 665.05 abcA | 406.54 abB | 415.09 bA | 436.90 bA | 162.27 aB | 194.91 aB | 240.45 aA |
| E. curvula | 627.01 aA | 643.02 aA | 667.10 abcA | 341.06 cC | 365.68 cB | 426.58 bcA | 163.69 aB | 194.19 aA | 213.42 abA |
| F. arundinacea | 410.70 eB | 460.71 dA | 487.79 dA | 293.48 dC | 339.06 dB | 374.45 dA | 148.73 aB | 166.91 aB | 216.50 abA |
| P. maximum | 507.95 dC | 556.01 cB | 629.01 cA | 292.79 dC | 318.02 dB | 374.09 dA | 72.87 bB | 91.78 bA | 113.38 cA |
| T. triandra | 613.12 abB | 648.67 aA | 688.01 aA | 397.88 abA | 405.15 bA | 413.77 cA | 79.39 bB | 95.18 bA | 130.89 cA |
| SE | 15.01 | 8.04 | 13.09 | ||||||
| Grass Species | PH | LW | TN | NL | CCI | RL | RM | BIO | ASH | OM | NDF | ADF | ADL | CP | Average |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A. pubescens | 5 | 5 | 6 | 6 | 6 | 4 | 2 | 2 | 1 | 6 | 5 | 9 | 3 | 3 | 5 |
| C. ciliaris | 8 | 7 | 3 | 5 | 5 | 9 | 6 | 5 | 8 | 4 | 6 | 8 | 4 | 4 | 6 |
| C. gayana | 4 | 6 | 8 | 9 | 7 | 6 | 7 | 9 | 7 | 3 | 4 | 3 | 7 | 5 | 6 |
| D. glomerata | 1 | 4 | 4 | 2 | 2 | 1 | 5 | 7 | 5 | 5 | 2 | 4 | 6 | 6 | 4 |
| D. eriantha | 6 | 8 | 2 | 3 | 4 | 7 | 9 | 4 | 3 | 8 | 7 | 7 | 9 | 2 | 5 |
| E. curvula | 7 | 1 | 5 | 7 | 9 | 5 | 4 | 3 | 2 | 9 | 8 | 5 | 8 | 8 | 6 |
| F. arundinacea | 2 | 3 | 7 | 1 | 1 | 2 | 3 | 1 | 6 | 1 | 1 | 2 | 5 | 9 | 4 |
| P. maximum | 3 | 9 | 9 | 4 | 8 | 8 | 8 | 8 | 9 | 2 | 3 | 1 | 1 | 7 | 6 |
| T. triandra | 9 | 2 | 1 | 8 | 3 | 3 | 1 | 6 | 4 | 7 | 9 | 6 | 2 | 1 | 4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Msiza, N.H.; Ravhuhali, K.E.; Mokoboki, H.K.; Mavengahama, S.; Motsei, L.E. Ranking Species for Veld Restoration in Semi-Arid Regions Using Agronomic, Morphological and Chemical Parameters of Selected Grass Species at Different Developmental Stages under Controlled Environment. Agronomy 2021, 11, 52. https://doi.org/10.3390/agronomy11010052
Msiza NH, Ravhuhali KE, Mokoboki HK, Mavengahama S, Motsei LE. Ranking Species for Veld Restoration in Semi-Arid Regions Using Agronomic, Morphological and Chemical Parameters of Selected Grass Species at Different Developmental Stages under Controlled Environment. Agronomy. 2021; 11(1):52. https://doi.org/10.3390/agronomy11010052
Chicago/Turabian StyleMsiza, Ntokozo Happy, Khuliso Emmanuel Ravhuhali, Hilda Kwena Mokoboki, Sydney Mavengahama, and Lebogang Ezra Motsei. 2021. "Ranking Species for Veld Restoration in Semi-Arid Regions Using Agronomic, Morphological and Chemical Parameters of Selected Grass Species at Different Developmental Stages under Controlled Environment" Agronomy 11, no. 1: 52. https://doi.org/10.3390/agronomy11010052