Nitrogen Fixation and Resource Partitioning in Alfalfa (Medicago sativa L.), Cicer Milkvetch (Astragalus cicer L.) and Sainfoin (Onobrychis viciifolia Scop.) Using 15N Enrichment under Controlled Environment Conditions



Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design and Treatments
2.2. Plant Sampling and Analysis
2.3. Statistical Analysis
3. Results
3.1. Nitrogen Derived from the Atmosphere (% Ndfa)
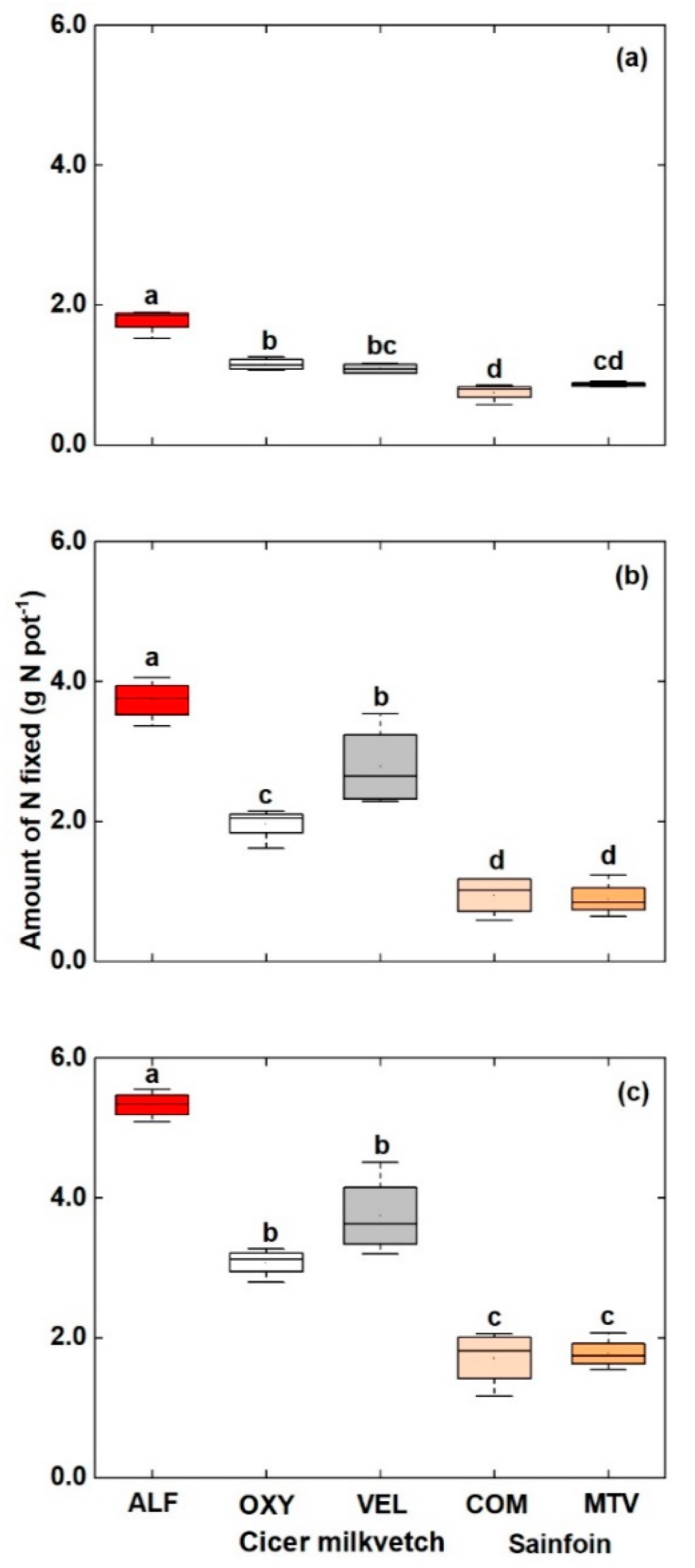
3.2. Amount of N fixed
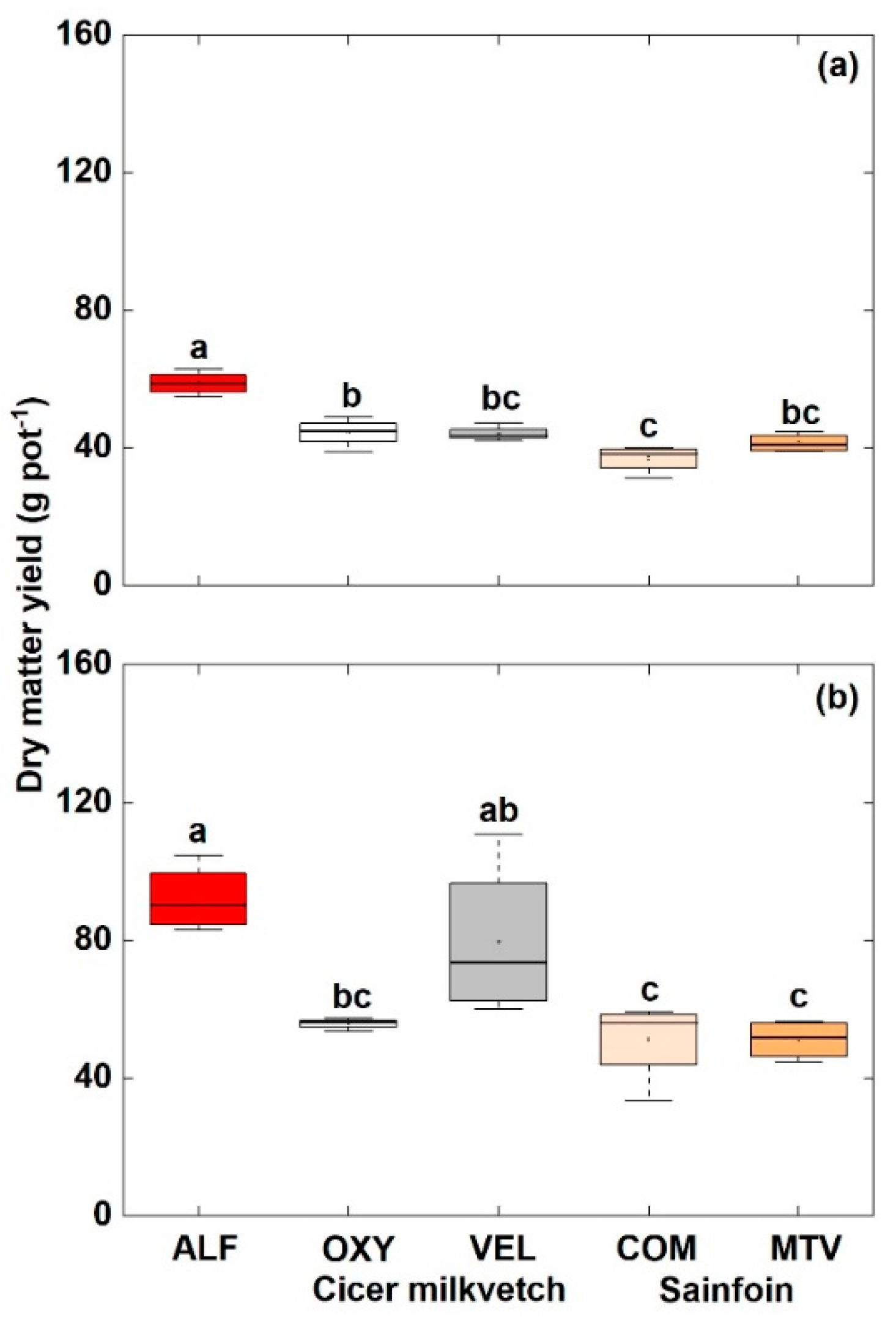
3.3. Shoot and Root Dry Matter Yield
4. Discussion
4.1. Percentage of Nitrogen Derived from the Atmosphere (% Ndfa) and the Amount of N Fixed
4.2. Shoot and Root Dry Matter Yield
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hauggaard-Nielsen, H.; Holdensen, L.; Wulfsohn, D.; Jensen, E.S. Spatial variation of N2-fixation in field pea (Pisum sativum L.) at the field scale determined by the 15N natural abundance method. Plant Soil 2010, 327, 167–184. [Google Scholar] [CrossRef]
- Ledgard, S.F.; Steele, K.W. Biological nitrogen fixation in mixed legume/grass pastures. Plant Soil 1992, 141, 137–153. [Google Scholar] [CrossRef]
- Pirhofer-Walzl, K.; Rasmussen, J.; Høgh-Jensen, H.; Eriksen, J.; Søegaard, K.; Rasmussen, J. Nitrogen transfer from forage legumes to nine neighboring plants in a multi-species grassland. Plant Soil 2012, 350, 71–84. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; McCaughey, W.P.; Grant, C.A. Pasture type and fertilization effects on N2 fixation, N budgets and external energy inputs in western Canada. Soil Biol. Biochem. 2004, 36, 1205–1212. [Google Scholar] [CrossRef]
- Oliveira, W.S.de.; Oliveira, P.P.A.; Corsi, M.; Duarte, F.R.S.; Tsai, S.M. Alfalfa yield and quality as function of nitrogen fertilization and symbiosis with Sinorhizobium meliloti. Sci. Agric. 2004, 61, 433–438. [Google Scholar] [CrossRef] [Green Version]
- Peoples, M.B.; Baldock, J.A. Nitrogen dynamics of pastures: Nitrogen fixation inputs, the impact of legumes on soil nitrogen fertility, and the contributions of fixed nitrogen to Australian farming systems. Aus. J. Exp. Agric. 2001, 41, 327–346. [Google Scholar] [CrossRef]
- McCaughey, W.P.; Chen, W. Benefits of including forage legumes in pastures and their management. In Proceedings of the Western Canadian Forage and Grazing Conference, Saskatoon, SK, Canada, 8–10 December 1999; Saskatchewan Stock Grower Association: Saskatoon, SK, Canada; pp. 97–115. [Google Scholar]
- Russelle, M.P.; Birr, A.S. Large-Scale Assessment of Symbiotic Dinitrogen Fixation by Crops: Soybean and Alfalfa in the Mississippi River Basin. Agron. J. 2004, 96, 1754–1760. [Google Scholar] [CrossRef] [Green Version]
- Burity, H.A.; Ta, T.C.; Faris, M.A.; Coulman, B.E. Estimation of nitrogen fixation and transfer from alfalfa to associated grasses in mixed swards under field conditions. Plant Soil 1989, 114, 249–255. [Google Scholar] [CrossRef]
- Kelner, D.J.; Vessey, J.K.; Entz, M.H. The nitrogen dynamics of 1-, 2- and 3-year stands of alfalfa in a cropping system. Agric. Ecosyst. Environ. 1997, 64, 1–10. [Google Scholar] [CrossRef]
- Zhu, Y.P.; Sheaffer, C.C.; Russelle, M.P.; Vance, C.P. Dinitrogen fixation of annual Medicago species. In Proceedings of the North American Alfalfa Improvement Conference, Oklahoma City, OK, USA, 30 March –2 April 1996; p. 75. [Google Scholar]
- Racca, R.; Basigalup, D.; Brenzoni, E.; Bruno, O.; Castell, C.; Collino, D.; Dardanelli, J.; Diaz Zorita, M.; Duhalde, J.; Gonzales, N.; et al. Alfalfa symbiotic dinitrogen fixation in the Argentine Pampean region. In Proceedings of the North American Alfalfa Improvement Conference, Bozeman, MT, USA, 2–6 August 1998; p. 71. [Google Scholar]
- Provorov, N.; Tickhonovich, I. Genetic Resources for improving nitrogen fixation in legume-rhizobia symbiosis. Gen. Res. Crop Evol. 2003, 50, 89–99. [Google Scholar] [CrossRef]
- Baimiev, A.K.; Baimiev, A.K.; Gubaidullin, I.I.; Kulikova, O.L.; Chemeris, A.V. Bacteria closely related to Phyllobacterium trifolii according to their 16S rRNA gene are discovered in the nodules of Hungarian sainfoin. Rus. J. Gen. 2007, 43, 587–590. [Google Scholar] [CrossRef]
- Burton, J.C.; Curley, R.L. Nodulation and nitrogen fixation in sainfoin (Onobrychis sativa LAM.) as influenced by strains of rhizobia. Mont. Agric. Exp. Stn. Bull. 1968, 627, 3–5. [Google Scholar]
- Prévost, D.; Bordeleau, L.M.; Caudry-Reznick, S.; Schulman, H.M.; Antoun, H. Characteristics of rhizobia isolated from three legumes indigenous to the Canadian high arctic: Astragalus alpinus, Oxytropis maydelliana, and Oxytropis arctobia. Plant Soil 1987, 98, 313–324. [Google Scholar] [CrossRef]
- Townsend, C.E. Breeding, physiology, culture, and utilization of cicer milkvetch (Astragalus cicer L.). Adv. Agron. 1993, 49, 253–308. [Google Scholar] [CrossRef]
- Li, Q.; Song, Y.; Li, G.; Yu, P. Grass-legume mixtures impact soil N, species recruitment, and productivity in temperate steppe grassland. Plant Soil 2015, 394, 271–285. [Google Scholar] [CrossRef]
- Nyfeler, D.; Huguenin-elie, O.; Suter, M.; Frossard, E.; Lüscher, A. Grass-legume mixtures can yield more nitrogen than legume pure stands due to mutual stimulation of nitrogen uptake from symbiotic and non-symbiotic sources. Agric. Ecosyst. Environ. 2011, 140, 155–163. [Google Scholar] [CrossRef]
- Schipanski, M.E.; Drinkwater, L.E. Nitrogen fixation in annual and perennial legume-grass mixtures across a fertility gradient. Plant Soil 2012, 357, 147–159. [Google Scholar] [CrossRef]
- Gregorich, E.G.; Drury, C.F.; Baldock, J.A. Changes in soil carbon under long-term maize in monoculture and legume-based rotation. Can. J. Soil Sci. 2001, 81, 21–31. [Google Scholar] [CrossRef] [Green Version]
- Gan, Y.; Liang, C.; Wang, X.; McConkey, B. Lowering carbon footprint of durum wheat by diversifying cropping systems. Field Crops Res. 2011, 122, 199–206. [Google Scholar] [CrossRef]
- Ledgard, S.F.; Brier, G.J.; Upsdell, M.P. Effects of clover cultivar on production and nitrogen fixation in clover-ryegrass swards under dairy cow grazing. N. Z. J. Agric. Res. 1990, 33, 243–249. [Google Scholar] [CrossRef]
- Acharya, S.N.; Kastelic, J.P.; Beauchemin, K.A.; Messenger, D.F. A review of research progress on cicer milkvetch (Astragalus cicer L.). Can. J. Plant Sci. 2006, 86, 49–62. [Google Scholar] [CrossRef]
- Acharya, S.N. A.A.C. Mountainview sainfoin (Onobrychis viciifolia subsp. Viciifolia). Can. J. Plant Sci. 2015, 95, 603–607. [Google Scholar] [CrossRef]
- Coulman, B.; Goplen, B.; Majak, W.; McAllister, T.; Cheng, K.J.; Berg, B.; Hall, J.; McCartney, D.; Acharya, S. A review of the development of a bloat-reduced alfalfa cultivar. Can. J. Plant Sci. 2000, 80, 487–491. [Google Scholar] [CrossRef] [Green Version]
- Rennie, R.J.; Dubetz, S. Nitrogen-15-determined nitrogen fixation in field-grown chickpea, lentil, faba bean, and field pea. Agron. J. 1986, 78, 654–660. [Google Scholar] [CrossRef]
- Ball, D.F. Loss-on-ignition as an estimate of organic matter and organic carbon in non-calcareous soils. Eur. J. Soil Sci. 1964, 15, 84–92. [Google Scholar] [CrossRef]
- Janzen, H.H. Soluble salts. In Soil Sampling and Methods of Analysis; Carter, M.R., Ed.; Lewis Publishers: Boca Raton, FL, USA, 1993; pp. 161–166. [Google Scholar]
- Qian, P.; Schoenau, J.J.; Karamanos, R.E. Simultaneous extraction of available phosphorus and potassium with a new soil test: A modification of Kelowna extraction. Commun. Soil Sci. Plant Anal. 1994, 25, 627–635. [Google Scholar] [CrossRef]
- Hu, Z.Y.; Zhao, F.J.; McGrath, S.P. Sulphur fractionation in calcareous soils and bioavailability to plants. Plant Soil 2005, 268, 103–109. [Google Scholar] [CrossRef]
- Burt, R. Soil Survey Field and Laboratory Methods Manual. Soil Survey Investigations Report No. 5. Method 3.2.1.2.2; United States Department of Agriculture Natural Resources Conservation Service: Lincoln, NB, USA, 2009.
- Acharya, S.; (Lethbridge Research and Development Centre, Agriculture and Agri-Food Canada (AAFC), Lethbridge, AB, Canada). Personal communication, 2020.
- Peoples, M.B.; Brockwell, J.; Herridge, D.F.; Rochester, I.J.; Alves, B.J.R.; Urquiaga, S.; Boddey, R.M.; Dakora, F.D.; Bhattarai, S.; Maskey, S.L.; et al. The contributions of nitrogen-fixing crop legumes to the productivity of agricultural systems. Symbiosis 2009, 48, 1–17. [Google Scholar] [CrossRef]
- Robinson, D.; Handley, L.L.; Scrimgeour, C.M.; Gordon, D.C.; Forster, B.P.; Ellis, R.P. Using stable isotope natural abundance (δ15N and δ 13C) to integrate the stress response of wild barley (Hordeum spontaneum C. Kock.) genotypes. J. Exp. Bot. 2000, 51, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Saxton, A.M. A macro for converting mean separation output to letter groupings in Proc Mixed. In Proceedings of the 23rd S.A.S. Users Group International, Cary, NC, USA, 22–25 March 1998. [Google Scholar]
- Malisch, C.S.; Suter, D.; Studer, B.; Lüscher, A. Multifunctional benefits of sainfoin mixtures: Effects of partner species, sowing density and cutting regime. Grass Forage Sci. 2017, 72, 794–805. [Google Scholar] [CrossRef]
- Cooper, C.S. Establishment, hay yield, and persistence of two sainfoin growth types seeded alone and with low growing grasses and legumes. Agron. J. 1972, 64, 379–381. [Google Scholar] [CrossRef]
- Sheehy, J.E.; Popple, S.C. Photosynthesis, water relations, temperature and canopy structure as factors influencing the growth of sainfoin (Onobrychis viciifolia. Scop.) and Lucerne (Medicago sativa L.). Ann. Bot. 1981, 48, 113–128. [Google Scholar] [CrossRef]
- Hayot-Carbonero, C. Sainfoin (Onobrychis viciifolia), a Forage Legume with Great Potential for Sustainable Agriculture, an Insight on its Morphological, Agronomical, Cytological and Genetic Characterization. Ph.D. Thesis, University of Manchester, Manchester, UK, 2011. [Google Scholar]
- Lardner, H.; Leah, P.; Damiran, D. Evaluation of cicer milkvetch and alfalfa varieties for nutritive value, anti-quality factors and animal preference. Sust. Agric. Res. 2019, 8, 1–10. Available online: http://sar.ccsenet.org (accessed on 8 December 2019).
- Ledgard, S.F. Nitrogen cycling in low input legume-based agriculture, with emphasis on legume/grass pastures. Plant Soil 2001, 228, 43–59. [Google Scholar] [CrossRef]
- Bolinder, M.A.; Janzen, H.H.; Gregorich, E.G.; Angers, D.A.; VandenBygaart, A.J. An approach for estimating net primary productivity and annual carbon inputs to soil for common agricultural crops in Canada. Agric. Ecosyst. Environ. 2007, 118, 29–42. [Google Scholar] [CrossRef]
- Fornara, D.A.; Tilman, D. Plant functional composition influences rates of soil carbon and nitrogen accumulation. J. Ecol. 2008, 96, 314–322. [Google Scholar] [CrossRef]
- Kielly, G.A.; Jefferson, P.G.; Lawrence, T.; Irvine, R.B. Evaluation of sainfoin-alfalfa mixtures for forage production and composition at a semi-arid location in southern Saskatchewan. Can. J. Plant Sci. 1994, 74, 785–791. [Google Scholar] [CrossRef]
- Aasen, A.; Bjorge, M. Alberta Forage Manual, 2nd ed.; Alberta Agriculture and Rural Development Information Management Services: Alberta, AB, Canada, 2009. Available online: https://www1.agric.gov.ab.ca/$Department/deptdocs.nsf/all/agdex16/$FILE/120_20-1_2009.pdf (accessed on 14 February 2018).
- Bhattarai, S.; Coulman, B.; Biligetu, B. Sainfoin (Onobrychis viciifolia Scop.): Renewed interest as a forage legume for western Canada. Can. J. Plant Sci. 2016, 96, 748–756. [Google Scholar] [CrossRef] [Green Version]
- Smoliak, S.; Hanna, M.R. Productivity of alfalfa, sainfoin and cicer milkvetch on sub-irrigated land when grazed by sheep. Can. J. Plant Sci. 1974, 55, 415–420. [Google Scholar] [CrossRef] [Green Version]
- Johnston, A.; Smoliak, S.; Hironaka, R.; Hanna, M.R. Oxley cicer milkvetch. Can. J. Plant Sci. 1971, 51, 428–429. [Google Scholar] [CrossRef] [Green Version]
- Barnes, R.F.; Miller, D.A.; Nelson, C.J. Forages. Volume 1: An Introduction to Grassland Agriculture, 5th ed.; Iowa State University Press: Ames, IA, USA, 1995. [Google Scholar]
- Popp, J.D. Beef Production on Alfalfa-Grass Pastures. Ph.D. Thesis, University of Saskatchewan, Saskatoon, SK, Canada, 1995. [Google Scholar]
- Popp, J.D.; McCaughey, W.P.; Cohen, R.D.H.; McAllister, T.A.; Majak, W. Enhancing pasture productivity with alfalfa: A review. Can. J. Plant. Sci. 2000, 80, 513–519. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Property | Value |
|---|---|
| pH (saturated paste) | 7.3 |
| Electrical conductivity (saturated paste) (dSm−1) | 1.6 |
| Organic matter (%) | 6.4 |
| Available phosphate-P (mg kg−1) | 59 |
| Available potassium (mg kg−1) | 570 |
| Available nitrate-N (mg kg−1) | 45 |
| Available sulphate-S (mg kg−1) | 15 |
| Cations in saturated paste extract | |
| Potassium (K) (mg L−1) | 86 |
| Calcium (Ca) (mg L−1) | 188 |
| Magnesium (Mg) (mg L−1) | 58 |
| Sodium (Na) (mg L−1) | 6 |
| Particle Size Analysis: Mini-Pipette Method | |
| % Sand (2.0 mm–0.05 mm) | 48.6 |
| % Silt (0.05 mm–2 µm) | 40.0 |
| % Clay (<2 µm) | 11.4 |
| Texture | Sandy Loam |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Issah, G.; Schoenau, J.J.; Lardner, H.A.; Knight, J.D. Nitrogen Fixation and Resource Partitioning in Alfalfa (Medicago sativa L.), Cicer Milkvetch (Astragalus cicer L.) and Sainfoin (Onobrychis viciifolia Scop.) Using 15N Enrichment under Controlled Environment Conditions. Agronomy 2020, 10, 1438. https://doi.org/10.3390/agronomy10091438
Issah G, Schoenau JJ, Lardner HA, Knight JD. Nitrogen Fixation and Resource Partitioning in Alfalfa (Medicago sativa L.), Cicer Milkvetch (Astragalus cicer L.) and Sainfoin (Onobrychis viciifolia Scop.) Using 15N Enrichment under Controlled Environment Conditions. Agronomy. 2020; 10(9):1438. https://doi.org/10.3390/agronomy10091438
Chicago/Turabian StyleIssah, Gazali, Jeff J. Schoenau, Herbert A. Lardner, and J. Diane Knight. 2020. "Nitrogen Fixation and Resource Partitioning in Alfalfa (Medicago sativa L.), Cicer Milkvetch (Astragalus cicer L.) and Sainfoin (Onobrychis viciifolia Scop.) Using 15N Enrichment under Controlled Environment Conditions" Agronomy 10, no. 9: 1438. https://doi.org/10.3390/agronomy10091438