Antioxidant Purple Corn Protein Concentrate from Germinated Andean Purple Corn Seeds

 ,
,  ,
, 
 ,
, 
Abstract
:1. Introduction
2. Material and Methods
2.1. Chemicals Reactive
2.2. Plant Material
2.3. Proximal Analysis of Purple Corn Flour
2.4. Germination of Purple Corn Seeds
2.5. Purple Corn Protein-Phenolic Concentrate (PCPC) from Germinated Seeds
2.6. PCPC Electrophoresis Analysis
2.7. PCPC Quantification of Total Phenolic Content (TPC)
2.8. Assay of Antioxidant Activity by ABTS Method
2.9. Assay of Antioxidant Activity by the Ferric-Reducing Antioxidant Power (FRAP) Method
2.10. Statistical Analysis
3. Results and Discussion
3.1. Purple Corn Flour Proximal Analysis
3.2. Germination (%) of Purple Corn Seeds
3.3. PCPCs Protein Profile
3.4. PCPCs Protein Content Quantification
3.5. TPC Content of Germinated PCPCs
3.6. Germinated PCPCs Antioxidant Activity by ABTS and FRAP Methods
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAOSTAT 2018 Food and Agriculture Organization Corporate Statistical Database. Available online: http://faostat.fao.org (accessed on 1 May 2020).
- Cuevas-Rodríguez, E.O.; Verdugo-Montoya, N.M.; Angulo-Bejarano, P.I.; Milan-Carrillo, J.; Mora-Escobedo, R.; Bello-Pérez, L.A.; Garzón-Tiznado, J.A.; Reyes-Moreno, C. Nutritional properties of tempeh flour from quality protein maize (Zea mays L.). LWT Food Sci. Technol. 2006, 39, 1072–1079. [Google Scholar] [CrossRef]
- Ul-Allah, S.; Ijaz, M.; Nawaz, A.; Sattar, A.; Sher, A.; Naeem, M.; Shahzad, U.; Nawaz, F.; Mahmood, K. Potassium Application Improves Grain Yield and Alleviates Drought Susceptibility in Diverse Maize Hybrids. Plants 2020, 9, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.; Zhai, W. Identification and antioxidant activity of anthocyanins extracted from the seed and cob of purple corn (Zea mays L.). Innov. Food Sci. Emerg. Technol. 2010, 11, 169–176. [Google Scholar] [CrossRef]
- Mrad, R.; Debs, E.; Saliba, R.; Maroun, R.G.; Louka, N. Multiple optimization of chemical and textural properties of roasted expanded purple maize using response surface methodology. J. Cereal Sci. 2014, 60, 397–405. [Google Scholar] [CrossRef]
- Kabir, S.H.; Das, A.K.; Rahman, M.S.; Singh, S.K.; Morshed, M.; Marma, A.S.H. Effect of genotype on proximate composition and biological yield of maize (Zea mays L.). AAES 2019, 4, 185–189. [Google Scholar] [CrossRef]
- Moreira, R.; Chenlo, F.; Arufe, S.; Rubinos, S.N. Physicochemical characterization of white, yellow and purple maize flours and rheological characterization of their doughs. J. Food Sci. Technol. 2015, 52, 7954–7963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Govender, V.; Aveling, T.A.S.; Kritzinger, Q. The effect of traditional storage methods on germination and vigour of maize (Zea mays L.) from northern KwaZulu-Natal and southern Mozambique. S. Afr. J. Bot. 2008, 74, 190–196. [Google Scholar] [CrossRef] [Green Version]
- Cuevas-Rodríguez, E.O.; MiIán-Carrillo, J.; Mora-Escobedo, R.; Cárdenas-Valenzuela, O.G.; Reyes-Moreno, C. Quality protein maize (Zea mays L.) tempeh flour through solid state fermentation process. LWT-Food Sci. Technol. 2004, 37, 59–67. [Google Scholar] [CrossRef]
- Wang, Y.; King, J.M.; Xu, Z.; Losso, J.; Prudente, A. Lutein from ozone-treated corn retains antimutagenic properties. J. Agric. Food Chem. 2008, 56, 7942–7949. [Google Scholar] [CrossRef]
- Carrillo, W.; Carpio, C.; Morales, D.; Vilcacundo, E.; Alvarez, M.; Silva, M. Content of fatty acids in corn (Zea mays L.) oil from Ecuador. Asian J. Pharm. Clin. Res. 2017, 10, 150–153. [Google Scholar]
- Wang, X.F.; Jing, X.M.; Lin, J. Starch mobilization in ultra-dried seed of maize (Zea mays L.) during germination. J. Integr. Plant Biol. 2005, 47, 443–451. [Google Scholar] [CrossRef]
- López-Martínez, L.X.; Leyva-López, N.; Gutiérrez-Grijalva, E.P.; Heredia, J.B. Effect of cooking and germination on bioactive compounds in pulses and their health benefits. J. Funct. Foods 2017, 38, 624–634. [Google Scholar] [CrossRef]
- Han, Z.; Wang, B.; Tian, L.; Wang, S.; Zhang, J.; Guo, S.; Zhang, H.; Xu, L.; Chen, Y. Comprehensive dynamic transcriptome analysis at two seed germination stages in maize (Zea mays L.). Physiol. Plant. 2020, 168, 205–217. [Google Scholar] [CrossRef]
- Sokrab, A.M.; Ahmed, I.A.M.; Babiker, E.E. Effect of germination on antinutritional factors, total, and extractable minerals of high and low phytate corn (Zea mays L.) genotypes. J. Saudi Soc. Agric. Sci. 2012, 11, 123–128. [Google Scholar] [CrossRef] [Green Version]
- Aguilera, Y.; Díaz, M.F.; Jiménez, T.; Benítez, V.; Herrera, T.; Cuadrado, C.; Martín-Cabrejas, M.A. Changes in non-nutritional factors and antioxidant activity during germination of nonconventional legumes. J. Agric. Food Chem. 2013, 61, 8120–8125. [Google Scholar] [CrossRef] [PubMed]
- Mbithi, S.; Van Camp, J.; Rodriguez, R.; Huyghebaert, A. Effects of sprouting on nutrient and antinutrient composition of kidney beans (Phaseolus vulgaris var. Rose coco). Eur. Food Res. Technol. 2001, 212, 188–191. [Google Scholar] [CrossRef]
- Benítez, V.; Cantera, S.; Aguilera, Y.; Mollá, E.; Esteban, R.M.; Díaz, M.F.; Martín-Cabrejas, M.A. Impact of germination on starch, dietary fiber and physicochemical properties in non-conventional legumes. Food Res. Int. 2013, 50, 64–69. [Google Scholar] [CrossRef]
- Carrillo, W.; Gómez-Ruiz, J.A.; Miralles, B.; Ramos, M.; Barrio, D.; Recio, I. Identification of antioxidant peptides of hen egg-white lysozyme and evaluation of inhibition of lipid peroxidation and cytotoxicity in the Zebrafish model. Eur. Food Res. Technol. 2016, 242, 1777–1785. [Google Scholar] [CrossRef] [Green Version]
- Piñuel, L.; Vilcacundo, E.; Boeri, P.; Barrio, D.A.; Morales, D.; Pinto, A.; Morán, R.; Samaniego, I.; Carrillo, W. Extraction of protein concentrate from red bean (Phaseolus vulgaris L.): Antioxidant activity and inhibition of lipid peroxidation. J. Appl. Pharm. Sci. 2019, 9, 1–14. [Google Scholar]
- Vilcacundo, R.; Barrio, D.; Carpio, C.; García-Ruiz, A.; Rúales, J.; Hernández-Ledesma, B.; Carrillo, W. Digestibility of quinoa (Chenopodium quinoa Willd) protein concentrate and its potential to inhibit lipid peroxidation in the Zebrafish larvae model. Plant Foods Hum. Nutr. 2017, 72, 294–300. [Google Scholar] [CrossRef]
- Saito, M.; Sakagami, H.; Fujisawa, S. Cytotoxicity and apoptosis induction by butylated hydroxyanisole (BHA) and butylated hydroxytoluene (BHT). Anticancer Res. 2003, 23, 4693–4701. [Google Scholar] [PubMed]
- Nieva-Echevarría, B.; Manzanos, M.J.; Goicoechea, E.; Guillén, M.D. 2,6-Di-tert-butyl-hydroxytoluene and its metabolites in foods. Compr. Rev. Food Sci. Food Saf. 2015, 14, 67–80. [Google Scholar] [CrossRef]
- Gawlik-Dziki, U.; Świeca, M.; Sułkowski, M.; Dziki, D.; Baraniak, B.; Czyż, J. Antioxidant and anticancer activities of Chenopodium quinoa leaves extracts–in vitro study. Food Chem. Toxicol. 2013, 57, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Vilcacundo, R.; Miralles, B.; Carrillo, W. Hernández-Ledesma B in vitro chemopreventive properties of peptides released from quinoa (Chenopodium quinoa Willd) protein under simulated gastrointestinal digestion. Food Res. Int. 2018, 105, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Vilcacundo, R.; Barrio, D.A.; Piñuel, L.; Boeri, P.; Tombari, A.; Welbaum, J.; Pinto, A.; Hernández-Ledesma, B.; Carrillo, W. Inhibition of lipid peroxidation of kiwicha (Amaranthus caudatus) hydrolyzed protein using zebrafish larvae and embryos. Plants 2018, 7, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, A.K.; Singh, V. Antioxidative free and bound phenolic constituents in pericarp, germ and endosperm of Indian dent (Zea mays var. indentata) and flint (Zea mays var. indurata) maize. J. Funct. Foods 2015, 13, 363–374. [Google Scholar] [CrossRef]
- Lao, F.; Sigurdson, G.T.; Giusti, M.M. Health benefits of purple corn (Zea mays L.) phenolic compounds. Compr. Rev. Food Sci. Food Saf. 2017, 16, 234–246. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; de Mejia, E.G.; Luna-Vital, D.; Tao, T.; Chandrasekaran, S.; Chatham, L.; Kumar, D. Relationship of phenolic composition of selected purple maize (Zea mays L.) genotypes with their anti-inflammatory, anti-adipogenic and anti-diabetic potential. Food Chem. 2019, 89, 739–750. [Google Scholar] [CrossRef]
- Dueñas, M.; Martiínez-Villaluenga, C.; Limón, R.I.; Peñas, E.; Frias, J. Effect of germination and elicitation on phenolic composition and bioactivity of kidney beans. Food Res. Int. 2015, 70, 55–63. [Google Scholar] [CrossRef] [Green Version]
- González-Montoya, M.; Hernández-Ledesma, B.; Silván, J.M.; Mora-Escobedo, R.; Martínez-Villaluenga, C. Peptides derived from in vitro gastrointestinal digestion of germinated soybean proteins inhibit human colon cancer cells proliferation and inflammation. Food Chem. 2018, 242, 75–82. [Google Scholar] [CrossRef] [Green Version]
- De Souza Rocha, T.; Hernandez, L.M.R.; Mojica, L.; Johnson, M.H.; Chang, Y.K.; de Mejía, E.G. Germination of Phaseolus vulgaris and alcalase hydrolysis of its proteins produced bioactive peptides capable of improving markers related to type-2 diabetes in vitro. Food Res. Int. 2015, 76, 150–159. [Google Scholar] [CrossRef] [Green Version]
- Mamilla, R.K.; Mishra, V.K. Effect of germination on antioxidant and ACE inhibitory activities of legumes. LWT Food Sci. Technol. 2017, 75, 51–58. [Google Scholar] [CrossRef]
- López-Amorós, M.L.; Hernández, T.; Estrella, I. Effect of germination on legume phenolic compounds and their antioxidant activity. J. Food Comp. Anal. 2006, 19, 277–283. [Google Scholar] [CrossRef]
- Cornejo, F.; Novillo, G.; Villacrés, E.; Rosell, C.M. Evaluation of the physicochemical and nutritional changes in two amaranth species (Amaranthus quitensis and Amaranthus caudatus) after germination. Food Res. Int. 2019, 121, 933–939. [Google Scholar] [CrossRef]
- Piñuel, L.; Boeri, P.; Zubillaga, F.; Barrio, D.A.; Torreta, J.; Cruz, A.; Vásquez, G.; Pinto, A.; Carrillo, W. Production of White, Red and Black Quinoa (Chenopodium quinoa Willd Var. Real) Protein Isolates and Its Hydrolysates in Germinated and Non-Germinated Quinoa Samples and Antioxidant Activity Evaluation. Plants 2019, 8, 257. [Google Scholar] [CrossRef] [Green Version]
- AOAC. Association of Official Analytical Chemists AOAC, Official Methods of Analysis, 18th ed.; AOAC: Gaithersburg, MD, USA, 2012. [Google Scholar]
- Instituto Ecuatoriano de Normalización (INEN). Frutas Frescas Tomate de árbol, Requisitos; INEN 1909; Norma Técnica Ecuatoriana: Quito, Ecuador, 2013; p. 12. [Google Scholar]
- Paucar-Menacho, L.M.; Martinez-Villaluenga, C.; Dueñas, M.; Frias, J.; Peñas, E. Optimization of germination time and temperature to maximize the content of bioactive compounds and the antioxidant activity of purple corn (Zea mays L.) by response surface methodology. LWT Food Sci. Technol. 2017, 76, 236–244. [Google Scholar] [CrossRef]
- Acosta, C.; Carpio, C.; Vilcacundo, R.; Carrillo, W. Identification of proteins isolate from amaranth (Amaranthus caudatus) by sodium dodecyl sulfate-polyacrylamide gel electrophoresis with water and NaCl 0.1 M solvents. Asian J. Pharm. Clin. Res. 2016, 9, 331–333. [Google Scholar]
- Serrano, S.; Rincón, F.; García-Olmo, J. Cereal protein analysis via Dumas method: Standardization of a micro-method using the Euro vector elemental analyzer. J. Cereal Sci. 2013, 58, 31–36. [Google Scholar] [CrossRef]
- Poveda, T.; Vilcacundo, R.; Carpio, C.; Carrillo, W. Analysis of sesame proteins isolate (Sesamum indicum L.) with water and salt treatment. Asian J. Pharm. Clin. Res. 2016, 9, 404–407. [Google Scholar]
- Lara, D.; Vilcacundo, E.; Carrillo, C.; Silva, M.; Alvarez, M.; Carpio, C.; Carrillo, W. Obtention of protein concentrate and polyphenols from macadamia (Macadamia integrifolia) with aqueous extraction method. Asian J. Pharm. Clin. Res. 2017, 10, 138–142. [Google Scholar]
- Boeri, P.; Piñuel, L.; Dalzotto, D.; Monasterio, R.; Fontana, A.; Sharry, S.; Barrio, D.A.; Carrillo, W. Argentine Patagonia barberry chemical composition and evaluation of its antioxidant capacity. J. Food Biochem. 2020. [Google Scholar] [CrossRef] [PubMed]
- Zambrano, M.; Vásquez, G.; Morales, D.; Vilcacundo, R.; Carrillo, W. Isolation of baby lima bean (Phaseolus lunatus) proteins fractions and evaluation of their antioxidant activity. Ital. J. Food Sci. 2020, 32. [Google Scholar] [CrossRef]
- Trehan, S.; Singh, N.; Kaur, A. Characteristics of white, yellow, purple corn accessions: Phenolic profile, textural, rheological properties and muffin making potential. J. Food Sci. Technol. 2018, 55, 2334–2343. [Google Scholar] [CrossRef] [PubMed]
- Mansilla, P.S.; Nazar, M.C.; Pérez, G.T. Flour functional properties of purple maize (Zea mays L.) from Argentina. Influence of environmental growing conditions. Int. J. Biol. Macromol. 2020, 146, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Messing, J. Proteome balancing of the maize seed for higher nutritional value. Front. Plant Sci. 2014, 5, 240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esen, A. A proposed nomenclature for the alcohol-soluble proteins (zeins) of maize (Zea mays L.). J. Cereal Sci. 1987, 5, 117–128. [Google Scholar] [CrossRef]
- Coleman, C.E.; Larkins, B.A. The prolamins of maize. In Seed Proteins; Shewry, P.R., Casey, R., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1998; pp. 109–139. [Google Scholar]
- Mertz, E.T.; Bates, L.S.; Nelson, O.E. Mutant gene that changes protein composition and increases lysine content of maize endosperm. Science 1964, 145, 279–280. [Google Scholar] [CrossRef]
- Esen, A. Separation of alcohol-soluble proteins (zeins) from maize into three fractions by differential solubility. Plant Physiol. 1986, 80, 623–627. [Google Scholar] [CrossRef] [Green Version]
- Lai, H.M.; Padua, G.W.; Wei, L.S. Properties and microstructure of zein sheets plasticized with palmitic and stearic acids. Cereal Chem. 1997, 74, 83–90. [Google Scholar] [CrossRef]
- Shukla, R.; Cheryan, M. Zein: The industrial protein from corn. Ind. Crops Prod. 2001, 13, 171–192. [Google Scholar] [CrossRef]
- Swallen, L.C. Zein. A new industrial protein. Ind. Eng. Chem. 1941, 33, 394–398. [Google Scholar] [CrossRef]
- Sharif, N.; Fabra, M.J.; López-Rubio, A. Nanostructures of zein for encapsulation of food ingredients. In Biopolymer Nanostructures for Food Encapsulation Purposes; Academic Press: New York, NY, USA, 2019; pp. 217–245. [Google Scholar]
- Zhang, M.; Reitmeier, C.A.; Hammond, E.G.; Myers, D.J. Production of textile fibers from zein and a soy protein-zein blend. Cereal Chem. 1997, 74, 594–598. [Google Scholar] [CrossRef]
- Morawsky, N.; Martino, G.T.; Guth, J.; Tsai, J.; Jeffcoat, R. Hydrolyzed Zein as Fixative in Hair Compositions. U.S. Patent 5,518,717, 21 May 1996. [Google Scholar]
- Mathiowitz, E.; Bernstein, H.; Morrel, E.; Schwaller, K. Method for Producing Protein Microspheres. U.S. Patent WO 91/06286, 16 May 1991. [Google Scholar]
- Peres, A.E.C.; Correa, M.I. Depression of iron oxides with corn starches. Miner. Eng. 1996, 9, 1227–1234. [Google Scholar] [CrossRef]
- Trezza, T.A.; Vergano, P.J. Grease resistance of corn zein coated paper. J. Food Sci. 1994, 59, 912–915. [Google Scholar] [CrossRef]
- Wang, J.Y.; Miyazawa, T.; Fujimoto, K. Inhibition of methyl linoleate peroxidation by maize zein in powder model system at high water activity. Agric. Biol. Chem. 1991, 55, 1531–1536. [Google Scholar]
- De la Parra, C.; Serna Saldivar, S.O.; Liu, R.H. Effect of processing on the phytochemical profiles and antioxidant activity of corn for production of masa, tortillas, and tortilla chips. J. Agric. Food Chem. 2007, 55, 4177–4183. [Google Scholar] [CrossRef] [PubMed]
- Mora-Rochin, S.; Gutiérrez-Uribe, J.A.; Serna-Saldivar, S.O.; Sánchez-Peña, P.; Reyes-Moreno, C.; Milán-Carrillo, J. Phenolic content and antioxidant activity of tortillas produced from pigmented maize processed by conventional nixtamalization or extrusion cooking. J. Cereal Sci. 2010, 52, 502–508. [Google Scholar] [CrossRef]
- Coco, M.G.; Vinson, J.A. Analysis of Popcorn (Zea Mays L. var. Everta) for Antioxidant Capacity and Total Phenolic Content. Antioxidants 2019, 8, 22. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Martinez, L.X.; Parkin, K.L.; Garcia, H.S. Antioxidant and quinone reductase inducing activities of ethanolic fractions from purple maize. LWT Food Sci. Technol. 2014, 59, 270–275. [Google Scholar] [CrossRef]



{kind=link}
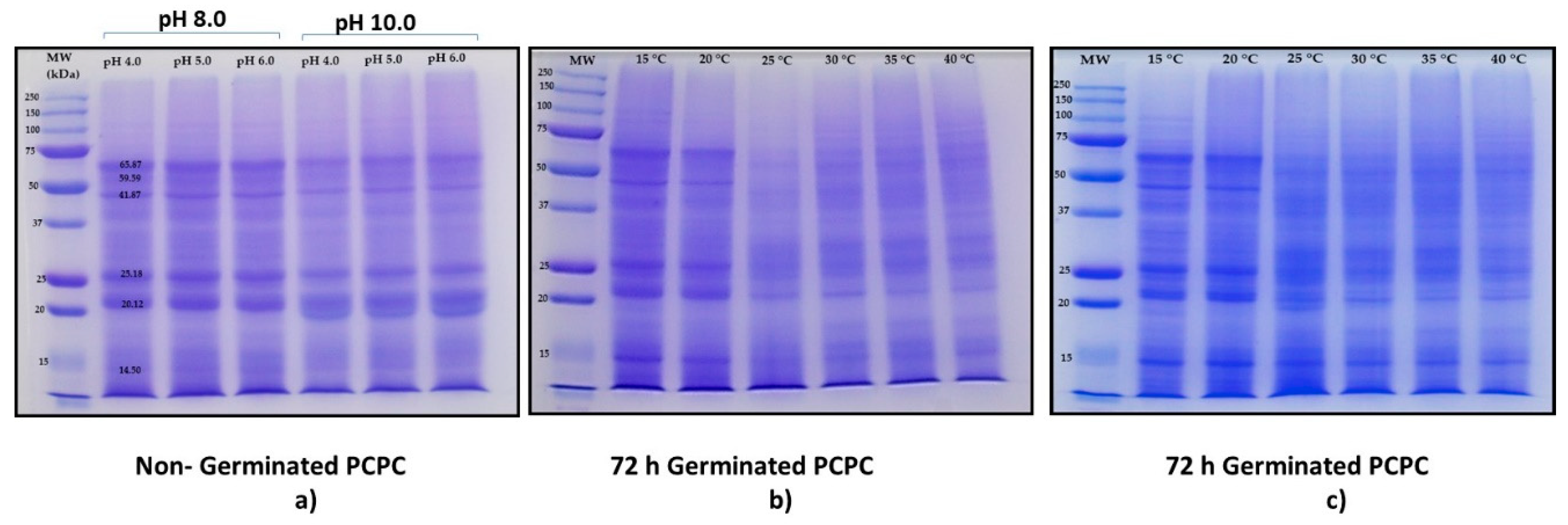{kind=link}
| % of Germination of Andean Purple Corn Seeds | ||||||
|---|---|---|---|---|---|---|
| Time (h) | 15 °C | 20 °C | 25 °C | 30 °C | 35 °C | 40 °C |
| 24 | 0.00 ± 0.00 | 0.00 ± 0.00 | 3.33 ± 0.58 a | 9.00 ± 3.56 a | 20.67 ± 4.04 a | 9.33 ± 2.08 a |
| 48 | 0.00 ± 0.00 | 3.33 ± 0.71 a | 42.00 ± 7.55 b | 33.00 ± 3.54 b | 36.67 ± 4.73 b | 22.67 ± 2.08 b |
| 72 | 0.00 ± 0.00 | 24.00 ± 6.24 b | 52.67 ± 8.74 c | 37.00 ± 3.54 b | 38.00 ± 5.29 b | 23.33 ± 2.52 b |
| 96 | 0.00 ± 0.00 | 38.67 ± 4.62 c | 54.00 ± 8.19 c | 41.00 ± 3.54 c | 38.67 ± 5.69b | 23.33 ± 2.52 b |
| 120 | 6.00 ± 0.58 a | 46.67 ± 7.43 d | 58.67 ± 8.39 d | 41.00 ± 3.54 c | 38.67 ± 5.69 b | 23.33 ± 2.52 b |
| 144 | 6.00 ± 0.58 a | 53.33 ± 4.51 e | 60.00 ± 8.66 d | 41.00 ± 3.54 c | 40.00 ± 4.58 c | 26.00 ± 2.00 c |
| 168 | 6.00 ± 0.58 a | 56.00 ± 4.58 e | 63.33 ± 7.23 e | 41.00 ± 3.54 c | 40.00 ± 4.58 c | 26.00 ± 2.00 c |
| pH (Alkaline-Ip) | 72 h | 120 h | 168 h |
|---|---|---|---|
| % Protein-germinated 15 °C | |||
| 8.0–4.0 | 25.16 ± 0.57 b | 21.81 ± 0.18 a | 24.00 ± 0.53 c |
| 8.0–5.0 | 24.09 ± 0.22 a | 21.97 ± 0.22 a | 23.06 ± 0.44 b |
| 8.0–6.0 | 24.00 ± 0.09 a | 21.94 ± 0.09 a | 21.06 ± 0.45 a |
| 10.0–4.0 | 27.16 ± 0.13 c | 24.88 ± 0.00 c | 25.31 ± 0.62 d |
| 10.0–5.0 | 26.34 ± 0.04 c | 24.34 ± 0.14 c | 23.91 ± 0.49 b |
| 10.0–6.0 | 24.34 ± 0.75 a | 22.00 ± 0.35 b | 23.06 ± 0.32 b |
| % Protein-germinated 20 °C | |||
| 8.0–4.0 | 23.16 ± 0.04 b | 18.63 ± 0.18 b | 25.72 ± 0.04 c |
| 8.0–5.0 | 22.56 ± 0.00 b | 23.44 ± 0.09 d | 16.84 ± 0.04 a |
| 8.0–6.0 | 20.31 ± 0.09 a | 16.50 ± 0.00 a | 22.08 ± 0.02 b |
| 10.0–4.0 | 23.50 ± 0.00 b | 27.06 ± 0.17 e | 28.19 ± 0.00 d |
| 10.0–5.0 | 22.97 ± 0.13 b | 24.22 ± 0.13 d | 24.48 ± 0.07 c |
| 10.0–6.0 | 20.09 ± 0.05 a | 21.88 ± 0.08 c | 22.53 ± 0.04 b |
| % Protein-germinated 25 °C | |||
| 8.0–4.0 | 17.78 ± 0.13 c | 10.69 ± 0.18 a | 22.16 ± 3.14 b |
| 8.0–5.0 | 18.69 ± 0.09 d | 9.44 ± 0.18 a | 19.09 ± 0.13 a |
| 8.0–6.0 | 18.25 ± 0.18 d | 9.38 ± 0.09 a | 19.75 ± 0.35 a |
| 10.0–4.0 | 14.88 ± 0.17 b | 27.50 ± 0.09 d | 33.56 ± 1.94 d |
| 10.0–5.0 | 13.19 ± 0.04 a | 16.88 ± 0.08 c | 29.06 ± 1.33 c |
| 10.0–6.0 | 13.00 ± 0.18 a | 12.47 ± 0.04 b | 30.00 ± 0.44 c |
| % Protein-germinated 30 °C | |||
| 8.0–4.0 | 26.56 ± 0.09 d | 26.44 ± 0.18 d | 23.75 ± 0.09 c |
| 8.0–5.0 | 25.19 ± 0.22 c | 24.38 ± 0.00 c | 21.00 ± 0.09 b |
| 8.0–6.0 | 22.66 ± 0.09 a | 21.44 ± 0.27 a | 18.38 ± 0.00 a |
| 10.0–4.0 | 26.25 ± 0.00 d | 27.28 ± 0.13 d | 29.38 ± 0.09 e |
| 10.0–5.0 | 26.75 ± 0.18 d | 26.50 ± 0.00 d | 27.53 ± 014 d |
| 10.0–6.0 | 24.19 ± 0.00 b | 23.71 ± 0.14 b | 19.31 ± 0.00 a |
| % Protein-germinated 35 °C | |||
| 8.0–4.0 | 26.60 ± 0.00 d | 26.50 ± 0.18 c | 23.80 ± 0.09 c |
| 8.0–5.0 | 25.20 ± 0.18 c | 24.34 ± 0.04 b | 21.06 ± 0.19 b |
| 8.0–6.0 | 22.60 ± 0.04 a | 21.44 ± 0.00 a | 18.40 ± 0.20 a |
| 10.0–4.0 | 26.25 ± 0.09 d | 27.25 ± 0.35 c | 29.40 ± 0.00 e |
| 10.0–5.0 | 25.53 ± 0.18 c | 26.56 ± 0.27 c | 27.60 ± 0.19 d |
| 10.0–6.0 | 24.19 ± 0.00 b | 23.75 ± 0.00 b | 19.35 ± 0.09 a |
| % of Protein-germinated 40 °C | |||
| 8.0–4.0 | 21.94 ± 0.88 b | 22.75 ± 0.09 a | 24.53 ± 0.49 c |
| 8.0–5.0 | 20.63 ± 0.09 b | 22.22 ± 0.13 a | 19.94 ± 1.06 b |
| 8.0–6.0 | 13.47 ± 2.25 a | 22.31 ± 0.09 a | 15.60 ± 2.96 a |
| 10.0–4.0 | 28.56 ± 0.35 d | 26.38 ± 0.35 b | 21.34 ± 0.04 b |
| 10.0–5.0 | 27.03 ± 1.64 d | 23.66 ± 0.75 a | 21.91 ± 0.04 b |
| 10.0–6.0 | 24.13 ± 1.77 c | 22.34 ± 2.87 a | 14.06 ± 0.27 a |
| TPC (mg GAE/g PCPC, DW) | |||||||
|---|---|---|---|---|---|---|---|
| pH | 24 h | 48 h | 72 h | 96 h | 120 h | 144 h | 168 h |
| TPC germinated at 15 °C | |||||||
| 8.0–4.0 | 910.48 ± 0.02 b | 834.29 ± 0.02 b | 1248.57 ± 0.11 d | 1170.00 ± 0.11 b | 900.95 ± 0.04 a | 1205.71 ± 0.03 c | 1289.05 ± 0.03 d |
| 8.0–5.0 | 1029.52 ± 0.01 c | 734.29 ± 0.01 a | 1158.10 ± 0.03 c | 1167.62 ± 0.04 b | 943.81 ± 0.03 b | 755.71 ± 0.01 a | 1112.86 ± 0.03 b |
| 8.0–6.0 | 710.48 ± 0.01 a | 712.86 ± 0.01 a | 812.86 ± 0.01 a | 898.57 ± 0.02 a | 1143.81 ± 0.02 d | 1243.81 ± 0.03 d | 886.67 ± 0.05 a |
| 10.0–4.0 | 948.57 ± 0.01 b | 1460.48 ± 0.01 e | 1243.81 ± 0.03 d | 1379.52 ± 0.04 c | 1172.38 ± 0.01 e | 1329.52 ± 0.03 e | 1381.90 ± 0.02 e |
| 10.0–5.0 | 1177.14 ± 0.02 d | 1074.76 ± 0.01 d | 1134.29 ± 0.06 c | 1370.00 ± 0.02 c | 1165.24 ± 0.02 e | 1458.10 ± 0.05 f | 1574.76 ± 0.05 f |
| 10.0–6.0 | 1058.10 ± 0.01 c | 984.29 ± 0.01 c | 936.67 ± 0.03 b | 1400.95 ± 0.04 d | 998.57 ± 0.06 c | 962.86 ± 0.02 b | 1196.19 ± 0.04 c |
| TPC germinated at 20 °C | |||||||
| 8.0–4.0 | 540.32 ± 0.01 b | 613.33 ± 0.00 b | 1248.57 ± 0.11 d | 1170.00 ± 0.11 b | 900.95 ± 0.04 a | 1205.71 ± 0.03 c | 1289.05 ± 0.03 d |
| 8.0–5.0 | 621.27 ± 0.01 c | 699.05 ± 0.01 c | 1158.10 ± 0.03 c | 1167.62 ± 0.04 b | 943.81 ± 0.03 b | 755.71 ± 0.01 a | 1112.86 ± 0.03 b |
| 8.0–6.0 | 416.51 ± 0.01 a | 554.60 ± 0.01 a | 812.86 ± 0.01 a | 898.57 ± 0.02 a | 1143.81 ± 0.02 c | 1243.81 ± 0.03 c | 886.67 ± 0.05 a |
| 10.0–4.0 | 694.29 ± 0.01 d | 741.90 ± 0.01 d | 1243.81 ± 0.03 d | 1379.52 ± 0.04 c | 1172.38 ± 0.01 c | 1329.52 ± 0.03 d | 1381.90 ± 0.02 e |
| 10.0–5.0 | 656.19 ± 0.01 d | 591.11 ± 0.01 a | 1134.29 ± 0.06 c | 1370.00 ± 0.02 c | 1165.24 ± 0.02 c | 1458.10 ± 0.05 e | 1574.76 ± 0.05 f |
| 10.0–6.0 | 614.92 ± 0.01 c | 762.54 ± 0.01 d | 936.67 ± 0.03 b | 1400.95 ± 0.04 d | 998.57 ± 0.06 c | 962.86 ± 0.02 b | 1196.19 ± 0.04 c |
| TPC germinated at 25 °C | |||||||
| 8.0–4.0 | 453.33 ± 0.03 b | 536.67 ± 0.02 c | 1820.00 ± 0.01 d | 1491.43 ± 0.03 e | 1100.95 ± 0.09 d | 1258.10 ± 0.01 c | 1529.52 ± 0.02 f |
| 8.0–5.0 | 350.95 ± 0.04 a | 522.38 ± 0.03 c | 1803.33 ± 0.05 d | 1217.62 ± 0.09 b | 920.00 ± 0.05 b | 1081.90 ± 0.01 a | 1250.95 ± 0.02 b |
| 8.0–6.0 | 1577.14 ± 0.04 f | 508.10 ± 0.03 b | 1746.19 ± 0.06 c | 1212.86 ± 0.17 b | 781.90 ± 0.04 a | 1136.67 ± 0.01 b | 1155.71 ± 0.03 a |
| 10.0–4.0 | 660.48 ± 0.01 d | 493.81 ± 0.02 b | 1550.95 ± 0.10 b | 1398.57 ± 0.05 d | 1541.43 ± 0.04 f | 1620.00 ± 0.00 e | 1315.24 ± 0.01 c |
| 10.0–5.0 | 617.62 ± 0.05 c | 531.90 ± 0.02 c | 1736.67 ± 0.08 c | 1284.29 ± 0.09 c | 1265.24 ± 0.01 e | 1362.86 ± 0.06 d | 1436.67 ± 0.03 e |
| 10.0–6.0 | 1172.38 ± 0.04 e | 384.29 ± 0.02 a | 1429.52 ± 0.03 a | 1003.33 ± 0.03 a | 972.38 ± 0.03 c | 1284.29 ± 0.03 c | 1355.71 ± 0.02 d |
| TPC germinated at 30 °C | |||||||
| 8.0–4.0 | 1143.81 ± 0.02 b | 993.81 ± 0.04 b | 1217.62 ± 0.02 c | 1222.38 ± 0.05 b | 1324.76 ± 0.03 c | 1277.14 ± 0.01 b | 1265.24 ± 0.06 b |
| 8.0–5.0 | 1179.52 ± 0.04 c | 1150.95 ± 0.03 e | 1229.52 ± 0.18 c | 1441.43 ± 0.02 e | 1196.19 ± 0.02 b | 1258.10 ± 0.01 b | 1343.81 ± 0.03 c |
| 8.0–6.0 | 1179.52 ± 0.04 c | 920.00 ± 0.01 a | 927.14 ± 0.02 a | 1003.33 ± 0.01 a | 1015.24 ± 0.02 a | 950.95 ± 0.01 a | 979.52 ± 0.04 a |
| 10.0–4.0 | 1486.67 ± 0.03 c | 1155.71 ± 0.03 d | 1284.29 ± 0.05 c | 1391.43 ± 0.07 d | 1472.38 ± 0.02 e | 1386.67 ± 0.01 d | 1389.05 ± 0.07 d |
| 10.0–5.0 | 1158.10 ± 0.01 b | 1008.10 ± 0.05 b | 1346.19 ± 0.06 d | 1481.90 ± 0.02 e | 1408.10 ± 0.05 d | 1358.10 ± 0.01 c | 1631.90 ± 0.03 e |
| 10.0–6.0 | 1022.38 ± 0.03 a | 1103.33 ± 0.01 c | 1191.43 ± 0.06 b | 1300.95 ± 0.01 c | 1329.52 ± 0.02 c | 1365.24 ± 0.01 c | 1393.81 ± 0.03 d |
| TPC germinated at 35 °C | |||||||
| 8.0–4.0 | 1500.95 ± 0.03 b | 1170.00 ± 0.01 c | 1355.71 ± 0.01 c | 1396.19 ± 0.04 d | 1289.05 ± 0.05 b | 1424.76 ± 0.04 e | 1448.57 ± 0.03 c |
| 8.0–5.0 | 1715.24 ± 0.06 e | 979.52 ± 0.02 a | 1227.14 ± 0.02 b | 1298.57 ± 0.04 c | 1467.62 ± 0.06 c | 1210.48 ± 0.04 b | 1436.67 ± 0.05 c |
| 8.0–6.0 | 1131.90 ± 0.03 a | 1074.76 ± 0.02 b | 1010.48 ± 0.02 a | 936.67 ± 0.03 a | 889.05 ± 0.03 a | 984.29 ± 0.04 a | 989.05 ± 0.03 a |
| 10.0–4.0 | 1708.10 ± 0.02 e | 1484.29 ± 0.02 d | 1455.71 ± 0.02 d | 1070.00 ± 0.00 b | 1446.19 ± 0.02 c | 1272.38 ± 0.02 c | 1605.71 ± 0.05 d |
| 10.0–5.0 | 1620.00 ± 0.00 d | 1648.57 ± 0.01 e | 1872.38 ± 0.01 f | 1446.19 ± 0.04 e | 1534.29 ± 0.05 d | 1289.05 ± 0.05 c | 1605.71 ± 0.02 d |
| 10.0–6.0 | 1593.81 ± 0.03 c | 1667.62 ± 0.02 e | 1767.62 ± 0.02 e | 1510.48 ± 0.01 f | 1446.19 ± 0.03 c | 1327.14 ± 0.04 d | 1174.76 ± 0.03 b |
| TPC germinated at 40 °C | |||||||
| 8.0–4.0 | 1150.95 ± 0.04 c | 1120.00 ± 0.06 b | 605.71 ± 0.02 a | 774.76 ± 0.04 c | 917.62 ± 0.03 b | 917.62 ± 0.03 b | 917.62 ± 0.03 b |
| 8.0–5.0 | 896.19 ± 0.04 b | 912.86 ± 0.04 a | 743.81 ± 0.04 c | 703.33 ± 0.03 b | 955.71 ± 0.05 c | 955.71 ± 0.05 c | 955.71 ± 0.02 c |
| 8.0–6.0 | 755.71 ± 0.06 a | 1110.48 ± 0.07 b | 691.43 ± 0.06 b | 674.76 ± 0.06 a | 824.76 ± 0.04 a | 824.76 ± 0.04 a | 824.76 ± 0.08 a |
| 10.0–4.0 | 1298.57 ± 0.04 e | 1424.76 ± 0.09 d | 1077.14 ± 0.03 d | 850.95 ± 0.03 d | 974.76 ± 0.02 c | 974.76 ± 0.02 c | 974.76 ± 0.01 c |
| 10.0–5.0 | 1236.67 ± 0.04 d | 1177.14 ± 0.01 c | 1079.52 ± 0.02 d | 953.33 ± 0.06 e | 1279.52 ± 0.05 e | 1279.52 ± 0.05 e | 1279.52 ± 0.01 e |
| 10.0–6.0 | 1327.14 ± 0.00 f | 1053.33 ± 0.03 c | 1070.00 ± 0.03 d | 924.76 ± 0.05 e | 1117.62 ± 0.03 d | 1117.62 ± 0.03 d | 1117.62 ± 0.03 d |
| ABTS (µmol TE/g PCPC, DW) | ||||||
|---|---|---|---|---|---|---|
| Germinated PCPC-pH 10/pH4.0 | ||||||
| Time (h) | 15 °C | 20 °C | 25 °C | 30 °C | 35 °C | 40 °C |
| 24 | 365.20 ± 0.01 d | 323.90 ± 0.01 c | 240.45 ± 0.01 b | 497.00 ± 0.05 d | 564.44 ± 0.09 a | 619.18 ± 0.05 f |
| 48 | 364.20 ± 0.01 d | 348.62 ± 0.01 c | 242.08 ± 0.09 b | 246.02 ± 0.05 a | 407.71 ± 0.10 d | 546.10 ± 0.02 e |
| 72 | 618.48 ± 0.04 c | 531.55 ± 0.02 d | 496.20 ± 0.03 c | 453.34 ± 0.03 c | 506.91 ± 0.06 e | 215.52 ± 0.06 a |
| 96 | 603.87 ± 0.03 c | 234.06 ± 0.01 b | 246.06 ± 0.03 b | 417.63 ± 0.03 b | 196.42 ± 0.03 a | 498.61 ± 0.05 d |
| 120 | 490.21 ± 0.09 b | 197.87 ± 0.06 a | 570.97 ± 0.07 d | 493.02 ± 0.04 d | 402.75 ± 0.09 c | 404.77 ± 0.07 b |
| 144 | 407.21 ± 0.09 a | 562.68 ± 0.02 e | 175.37 ± 0.01 a | 547.58 ± 0.02 e | 804.35 ± 0.08 e | 488.44 ± 0.05 d |
| 168 | 490.45 ± 0.09 b | 571.49 ± 0.09 e | 603.60 ± 0.03 e | 278.57 ± 0.06 a | 235.10 ± 0.04 b | 470.05 ± 0.01 c |
| FRAP (µmol TE/g PCPC, DW) | ||||||
|---|---|---|---|---|---|---|
| Germinated PCPC pH 10-pH 4.0 | ||||||
| Time (h) | 15 °C | 20 °C | 25 °C | 30 °C | 35 °C | 40 °C |
| 24 | 257.98 ± 0.01 c | 208.58 ± 0.01 a | 154.86 ± 0.01 a | 155.52 ± 0.93 a | 168.25 ± 3.33 a | 173.61 ± 0.01 a |
| 48 | 243.44 ± 0.01 c | 243.40 ± 0.01 b | 165.11 ± 0.02 a | 454.86 ± 0.01 a | 771.68 ± 0.67 c | 611.19 ± 0.09 c |
| 72 | 741.49 ± 0.09 d | 770.21 ± 0.21 d | 678.18 ± 0.10 d | 815.04 ± 0.20 c | 922.25 ± 0.52 d | 628.94 ± 0.70 c |
| 96 | 807.29 ± 0.09 e | 339.15 ± 0.36 b | 335.36 ± 0.91 c | 750.69 ± 0.44 b | 349.39 ± 0.02 a | 552.42 ± 0.59 b |
| 120 | 100.46 ± 0.21 a | 286.71 ± 0.69 c | 772.85 ± 0.29 c | 877.92 ± 0.36 d | 747.02 ± 0.09 c | 452.69 ± 0.06 a |
| 144 | 121.00 ± 0.21 b | 751.10 ± 0.49 d | 236.10 ± 1.52 b | 984.83 ± 0.38 e | 363.58 ± 0.51 a | 699.23 ± 3.17 d |
| 168 | 120.01 ± 0.21 b | 800.83 ± 0.30 e | 817.12 ± 0.55 e | 762.63 ± 0.13 b | 435.30 ± 0.38 b | 701.44 ± 0.35 e |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vilcacundo, E.; García, A.; Vilcacundo, M.; Morán, R.; Samaniego, I.; Carrillo, W. Antioxidant Purple Corn Protein Concentrate from Germinated Andean Purple Corn Seeds. Agronomy 2020, 10, 1282. https://doi.org/10.3390/agronomy10091282
Vilcacundo E, García A, Vilcacundo M, Morán R, Samaniego I, Carrillo W. Antioxidant Purple Corn Protein Concentrate from Germinated Andean Purple Corn Seeds. Agronomy. 2020; 10(9):1282. https://doi.org/10.3390/agronomy10091282
Chicago/Turabian StyleVilcacundo, Edgar, Antón García, Mario Vilcacundo, Roberto Morán, Iván Samaniego, and Wilman Carrillo. 2020. "Antioxidant Purple Corn Protein Concentrate from Germinated Andean Purple Corn Seeds" Agronomy 10, no. 9: 1282. https://doi.org/10.3390/agronomy10091282