Identification of Quantitative Trait Loci for Spikelet Fertility at the Booting Stage in Rice (Oryza sativa L.) under Different Low-Temperature Conditions



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Mapping Population
2.2. Genotyping Using SSR and SNP Markers
2.3. Phenotyping of Cold Tolerance
2.4. Linkage Map Construction and QTL Analysis
3. Results
3.1. Phenotypic Variation in Parental Cultivars and RILs
3.2. Linkage Map Construction
3.3. Identification of QTL for Spikelet Fertility under Field and Greenhouse Conditions
3.4. QTL Identification for Agronomic Traits Related to Cold Tolerance
4. Discussion
4.1. Potential Use of the Cold Tolerance QTL in Inter-Subspecies Rice Breeding
4.2. Reliable Cold Tolerance QTL at the Booting Stage Identified by the Dual Screening System
4.3. Comparison with Previously Identified Cold Tolerance QTL at the Booting Stage
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Yoshida, S. Growth and development of the rice plant. In Fundamentals of Rice Crop Science; International Rice Research Institute: Los Baños, Philippines, 1981; pp. 1–61. [Google Scholar]
- Xie, G.; Kato, H.; Imai, R. Biochemical identification of the OsMKK6-OsMPK3 signalling pathway for chilling stress tolerance in rice. Biochem. J. 2012, 443, 95–102. [Google Scholar]
- Lee, M. Low temperature tolerance in rice: The Korean experience. In Proceedings of the Conference: Increased Lowland Rice Production in the Mekong Region, Vientiane, Laos, 30 October–2 November 2000. [Google Scholar]
- Farrell, T.; Fox, K.M.; Williams, R.L.; Fukai, S.; Reinke, R.F.; Lewin, L.G. Temperature constraints to rice production in Australia and Laos: A shared problem. In Proceedings of the Conference: Increased Lowland Rice Production in the Mekong Region, Vientiane, Laos, 30 October–2 November 2000. [Google Scholar]
- Xu, L.M.; Zhou, L.; Zeng, Y.W.; Wang, F.; Zhang, H.; Shen, S.; Li, Z. Identification and mapping of quantitative trait loci for cold tolerance at the booting stage in a japonica rice near-isogenic line. Plant. Sci. 2008, 174, 340–347. [Google Scholar]
- Satake, T.; Hayase, H. Male sterility caused by cooling treatment at the young microspore stage in rice plants: V. Estimations of pollen developmental stage and the most sensitive stage to coolness. Jpn. J. Crop. Sci. 1970, 39, 468–473. [Google Scholar]
- Mackill, D.J.; Coffman, W.R.; Garrity, D.P. Rainfed Lowland Rice Improvement; International Rice Research Institute: Los Baños, Philippines, 1996. [Google Scholar]
- Saito, K.; Miura, K.; Nagano, K.; Hayano-Saito, Y.; Saito, A.; Araki, H.; Kato, A. Chromosomal location of quantitative trait loci for cool tolerance at booting stage in rice variety ‘Norin-PL8’. Breed. Sci. 1995, 45, 337–340. [Google Scholar]
- Takeuchi, Y.; Hayasaka, H.; Chiba, B.; Tanaka, I.; Shimano, T.; Yamagishi, M.; Nagano, K.; Sasaki, T.; Yano, M. Mapping quantitative trait loci controlling cool-temperature tolerance at booting stage in temperate japonica rice. Breed. Sci. 2001, 51, 191–197. [Google Scholar]
- Shinada, H.; Iwata, N.; Sato, T.; Fujino, K. Genetical and morphological characterization of cold tolerance at fertilization stage in rice. Breed. Sci. 2013, 63, 197–204. [Google Scholar]
- Saito, K.; Miura, K.; Nagano, K.; Hayano-Saito, Y.; Araki, H.; Kato, A. Identification of two closely linked quantitative trait loci for cold tolerance on chromosome 4 of rice and their association with anther length. Theor. Appl. Genet. 2001, 103, 862–868. [Google Scholar]
- Saito, K.; Hayano-Saito, Y.; Maruyama-Funatsuki, W.; Sato, Y.; Kato, A. Physical mapping and putative candidate gene identification of a quantitative trait locus Ctb1 for cold tolerance at the booting stage of rice. Theor. Appl. Genet. 2004, 109, 515–522. [Google Scholar]
- Saito, K.; Hayano-Saito, Y.; Kuroki, M.; Sato, Y. Map-based cloning of the rice cold tolerance gene Ctb1. Plant. Sci. 2010, 179, 97–102. [Google Scholar]
- Andaya, V.C.; Mackill, D.J. QTLs conferring cold tolerance at the booting stage of rice using recombinant inbred lines from a japonica × indica cross. Theor. Appl Genet. 2003, 106, 1084–1090. [Google Scholar]
- Kuroki, M.; Saito, K.; Matsuba, S.; Yokogami, N.; Shimizu, H.; Ando, I.; Sato, Y. A quantitative trait locus for cold tolerance at the booting stage on rice chromosome 8. Theor. Appl Genet. 2007, 115, 593–600. [Google Scholar]
- Zhang, Z.; Li, J.; Pan, Y.; Li, J.; Zhou, L.; Shi, H.; Zeng, Y.; Guo, H.; Yang, S.; Zheng, W.; et al. Natural variation in CTB4a enhances rice adaptation to cold habitats. Nat. Commun. 2017, 8, 14788. [Google Scholar]
- Suh, J.P.; Jeung, J.U.; Lee, J.I.; Choi, Y.H.; Yea, J.D.; Virk, P.S.; Mackill, D.J.; Jena, K.K. Identification and analysis of QTLs controlling cold tolerance at the reproductive stage and validation of effective QTLs in cold-tolerant genotypes of rice (Oryza sativa L.). Theor. Appl. Genet. 2010, 120, 985–995. [Google Scholar]
- Yang, L.M.; Liu, H.L.; Zhao, H.W.; Wang, J.G.; Sun, J.; Zheng, H.L.; Lei, L.; Zou, D.T. Mapping quantitative trait loci and meta-analysis for cold tolerance in rice at booting stage. Euphytica 2019, 215, 89. [Google Scholar]
- Cho, Y.G.; Kang, M.R.; Kim, Y.W.; Eun, M.Y.; Chung, T.Y.; Lee, Y.T. Development of RFLP framework map of rice (Oryza sativa L.) using recombinant inbreed population derived from Milyang23/Gihobyeo cross. Korean J. Breed. 1998, 30, 289–297. [Google Scholar]
- Ji, H.; Kim, H.; Lee, G.; Yoon, U.; Kim, T.; Seol, Y.; Yun, D.; Koh, H.; Eun, M. Molecular genetic and physical map using PCR-based DNA markers with the recombinant inbred population derived from Milyang23/Gihobyeo cross. Korean J. Breed. 2012, 44, 273–281. [Google Scholar]
- Murray, M.G.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980, 8, 4321–4325. [Google Scholar]
- Thomson, M.J.; Zhao, K.; Wright, M.; McNally, K.L.; Rey, J.; Tung, C.; Reynolds, A.; Scheffler, B.; Eizenga, G.; McClung, A.; et al. High-throughput single nucleotide polymorphism genotyping for breeding applications in rice using the BeadXpress platform. Mol. Breed. 2012, 29, 875–886. [Google Scholar]
- Wright, M.H.; Tung, C.W.; Zhao, K.; Reynolds, A.; McCouch, S.R.; Bustamante, C.D. Alchemy: A reliable method for automated SNP genotype calling for small batch sizes and highly homozygous populations. Bioinformatics 2010, 26, 2952–2960. [Google Scholar]
- Tanno, H.; Kinoshita, M.; Kiuchi, H.; Hirayama, Y.; Kikuchi, H. Estimation of cool-weather tolerance at the flowering stage and its relationship with that of the booting stage in Hokkaido rice varieties. Jpn. J. Crop. Sci. 2000, 69, 493–499. [Google Scholar]
- Lincoln, S.E.; Daly, M.J.; Lander, E.S. Constructing Genetic Linkage Maps With MAPMAKER/EXP Version 3.0 A Tutorial and Reference Manual; A Whitehead Institute for Biomedical Research Technical Report; Whitehead Institute for Biomedical Research: Cambridge, MA, USA, 1993. [Google Scholar]
- Wang, J.; Li, H.; Zhang, L.; Li, C.; Meng, L. Users’ Manual of QTL IciMapping v3.1; Institute of Crop Science, CAAS: Beijing, China; Crop Research Informatics Lab: Mexico City, Mexico, 2011. [Google Scholar]
- Septiningsih, E.M.; Sanchez, D.L.; Singh, N.; Sendon, P.M.; Pamplona, A.M.; Heuer, S.; Mackill, D.J. Identifying novel QTLs for submergence tolerance in rice cultivars IR72 and Madabaru. Theor. Appl. Genet. 2012, 124, 867–874. [Google Scholar]
- Kawahara, Y.; Bastide, M.; Hamilton, J.P.; Kanamori, H.; McCombie, W.R.; Ouyang, S.; Schwartz, D.C.; Tanaka, T.; Wu, J.; Zhou, S.; et al. Improvement of the Oryza sativa Nipponbare reference genome using next generation sequence and optical map data. Rice 2013, 6, 4. [Google Scholar]
- Mackill, D.J.; Lei, X. Genetic variation for traits related to temperate adaptation of rice cultivars. Crop. Sci. 1997, 37, 1340–1346. [Google Scholar]
- Pan, Y.; Zhang, H.; Zhang, D.; Li, J.; Xiong, H.; Yu, J.; Li, J.; Rashid, M.A.R.; Li, G.; Ma, X.; et al. Genetic analysis of cold tolerance at the germination and booting stages in rice by association mapping. PLoS ONE 2015, 10, e0120590. [Google Scholar]
- Kim, B.; Kim, D.G.; Lee, G.; Seo, J.; Choi, I.Y.; Choi, B.S.; Yang, T.J.; Kim, K.S.; Lee, J.; Chin, J.H.; et al. Defining the genome structure of ‘Tongil’ rice, an important cultivar in the Korean “Green Revolution”. Rice 2014, 7, 22. [Google Scholar]
- Won, Y.J.; Ahn, E.K.; Lee, J.H.; Suh, J.P.; Lee, J.S.; Chang, J.K.; Choi, Y.H.; Hwang, T.S.; Lim, Y.T.; Ha, W.G.; et al. Regional yield variation of some Korean high-yielding rice varieties. Korean J. Int. Agric. 2014, 26, 414–418. [Google Scholar]
- Lee, C.M.; Park, H.S.; Baek, M.K.; Suh, J.P.; Kim, C.S.; Lee, K.M.; Park, S.G.; Cho, Y.C. Characterization of grain-related traits of 300 Korean rice varieties. In Proceedings of the Conference: 2019 Korean Society of Breeding Science (KSBS) & Society for the Advancement of Breeding Research in Asia and Oceania (SABRAO) international conference on plant breeding for sustainable development, Gwangju, Korea, 2–5 July 2019. [Google Scholar]
- Satake, T. Male sterility caused by cooling treatment at the young microspore stage in rice plants. XXIX. The mechanism of enhancement in cool tolerance by raising water temperature before the critical stage. Jpn. J. Crop. Sci. 1989, 58, 240–245. [Google Scholar]
- Dai, L.; Lin, X.; Ye, C.; Ise, K.; Saito, K.; Kato, A.; Xu, F.; Yu, T.; Zhang, D. Identification of quantitative trait loci controlling cold tolerance at the reproductive stage in Yunnan landrace of rice, Kunmingxiaobaigu. Breed. Sci. 2004, 54, 253–258. [Google Scholar]
- Oh, C.S.; Choi, Y.H.; Lee, S.J.; Yoon, D.; Moon, H.; Ahn, S. Mapping of quantitative trait loci for cold tolerance in weedy rice. Breed. Sci. 2004, 54, 373–380. [Google Scholar]
- Lan, T.; Liang, K.J.; Chen, Z.W.; Duan, Y.L.; Wang, J.L.; Ye, N.; Wu, W.R. Genetic analysis and gene mapping of cold-indued seedling chlorosis in rice. Hereditas 2007, 29, 1121–1125. [Google Scholar]
- Lan, T.; Wang, B.; Ling, Q.; Xu, C.; Tong, Z.; Liang, K.; Duan, Y.; Jin, J.; Wu, W. Fine mapping of cisc(t), a gene for cold-induced seedling chlorosis and identification of its candidate in rice. Chin. Sci. Bull. 2010, 55, 3149–3153. [Google Scholar]




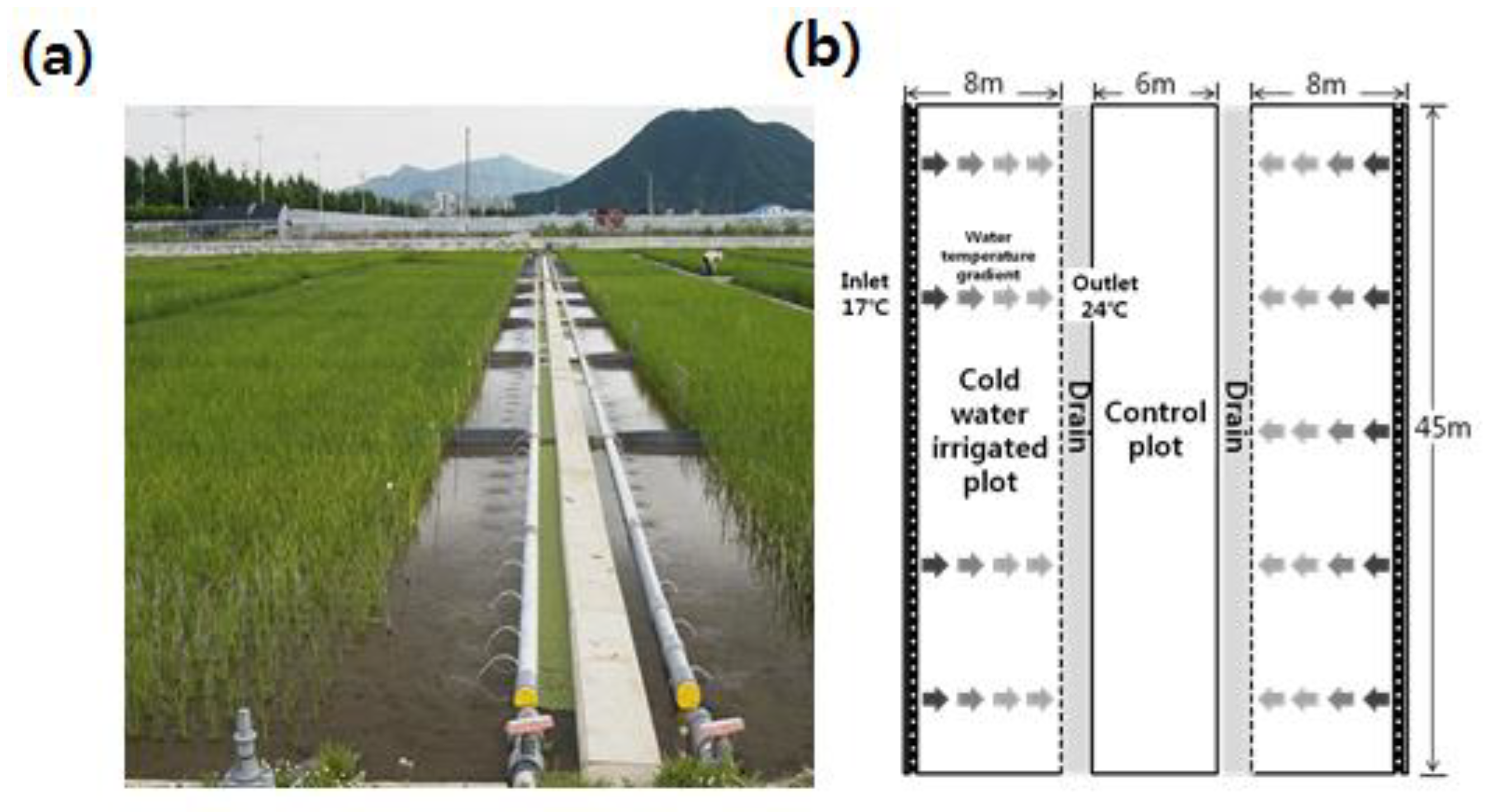{kind=link}
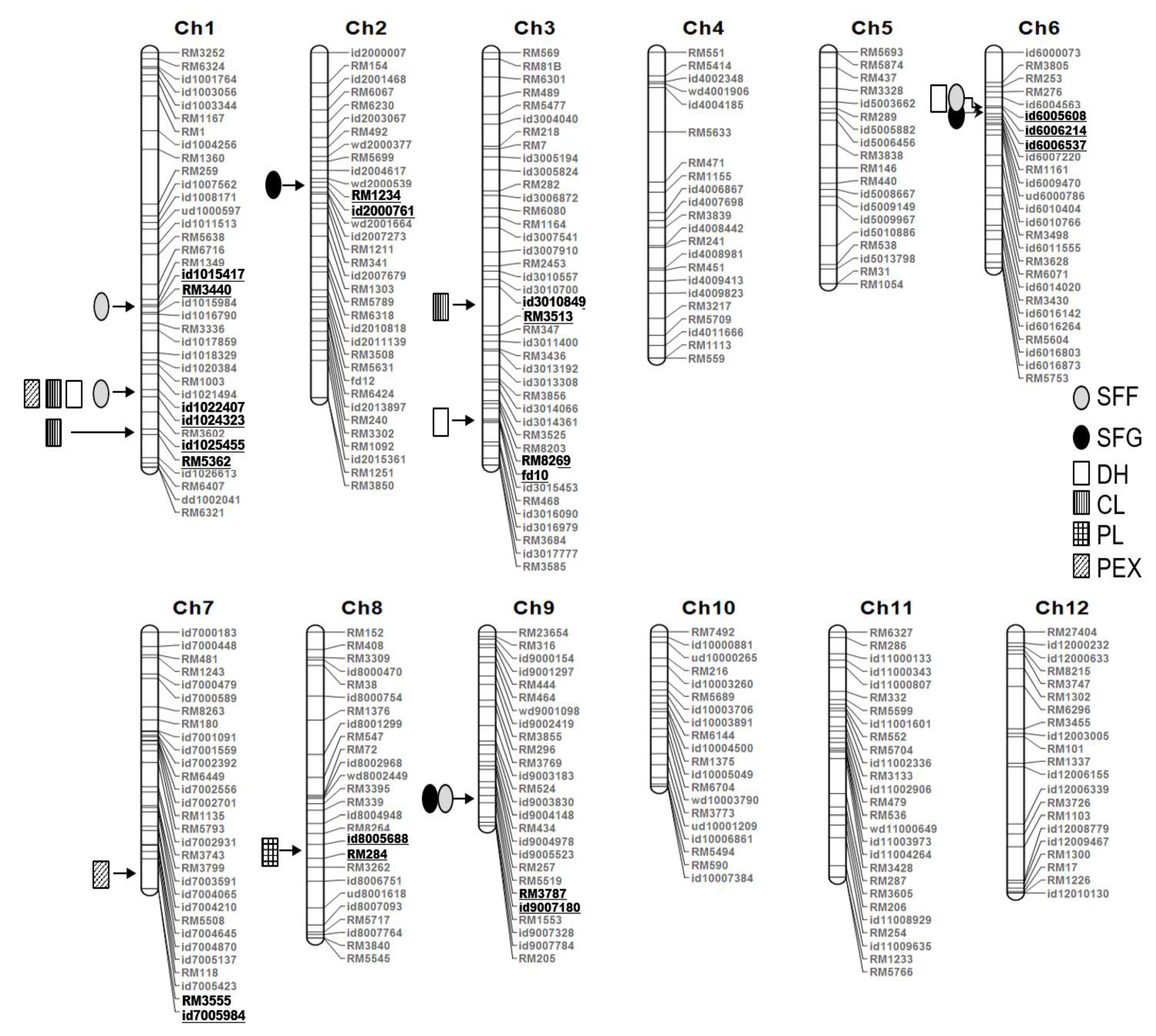{kind=link}
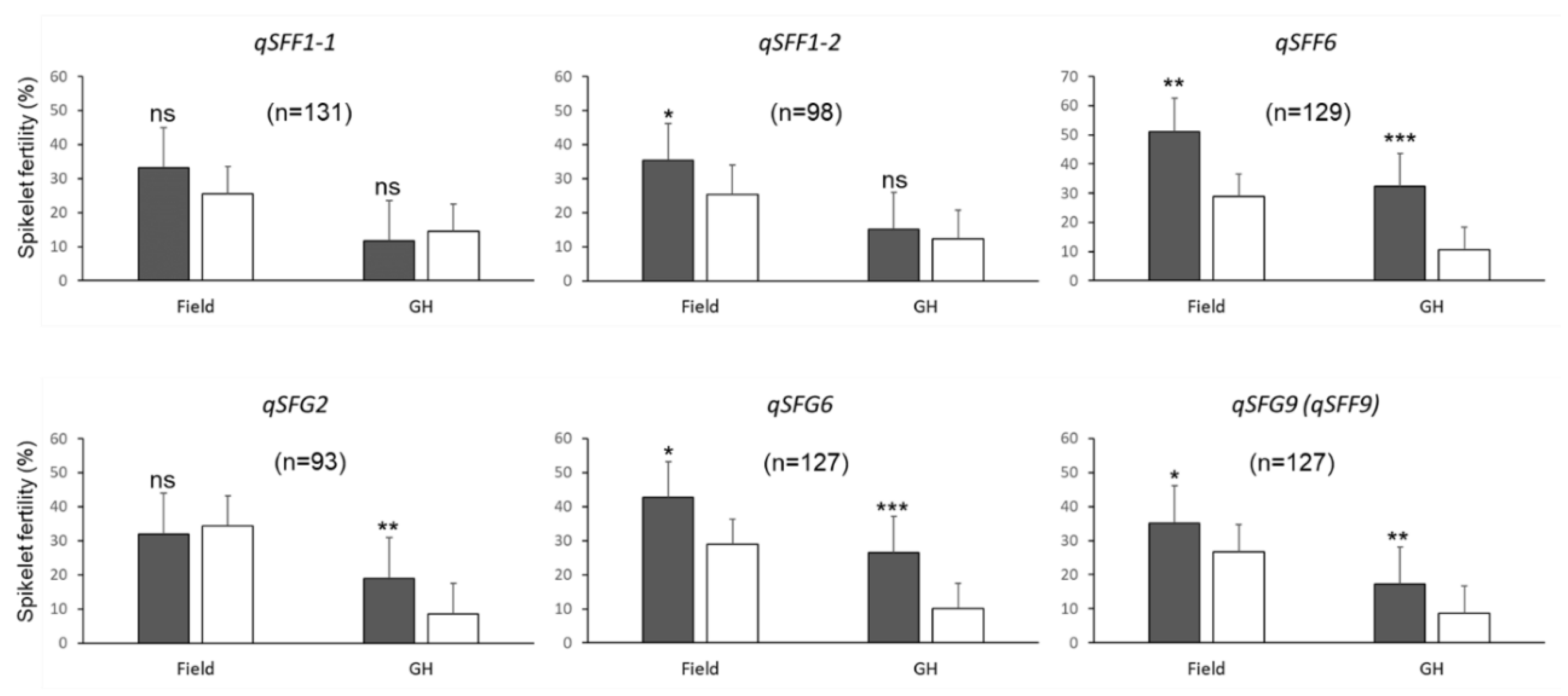{kind=link}
| Evaluation Method a | Treatment | Parent (Mean ± SD) | RIL c | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Milyang23 | Giho b | Mean | Var | CV | Min | Max | Skewness | Kurtosis | |||
| Field | Control | 90.3 ± 1.73 | 90.0 ± 1.73 | ns | 73.1 | 155.1 | 0.17 | 29.4 | 95.3 | −0.919 | 0.982 |
| Cold | 51.5 ± 2.08 | 76.0 ± 5.13 | * | 30.6 | 481.9 | 0.72 | 0.0 | 86.3 | 0.567 | −0.481 | |
| Greenhouse | Control | 87.5 ± 4.03 | 89.9 ± 3.66 | ns | 73.6 | 180.5 | 0.18 | 8.4 | 97.7 | −1.457 | 4.198 |
| Cold | 12.4 ± 2.33 | 55.7 ± 4.18 | ** | 13.2 | 264.2 | 1.24 | 0.0 | 82.7 | 1.841 | 3.405 | |
| Trait a | Treatment | Parent (Mean ± SD) | RIL c | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Milyang23 | Giho b | Mean | Var | CV | Min | Max | Skewness | Kurtosis | |||
| DH | Control | 101.7 ± 1.15 | 100.7 ± 1.15 | ns | 98.7 | 30.2 | 0.06 | 84.0 | 130.0 | 2.124 | 9.583 |
| Cold | 120.3 ± 2.52 | 111.3 ± 0.58 | * | 109.3 | 43.4 | 0.06 | 96.0 | 141.0 | 1.500 | 4.687 | |
| CL | Control | 65.3 ± 1.53 | 77.0 ± 2.00 | * | 74.3 | 186.5 | 0.18 | 40.0 | 103.2 | −0.085 | −0.493 |
| Cold | 37.0 ± 7.00 | 66.0 ± 1.73 | * | 50.9 | 168.1 | 0.26 | 24.6 | 81.6 | 0.080 | −0.753 | |
| PL | Control | 20.3 ± 1.15 | 22.5 ± 1.29 | ns | 20.7 | 4.8 | 0.11 | 15.4 | 25.4 | 0.100 | −0.321 |
| Cold | 18.3 ± 1.55 | 17.2 ± 0.35 | ns | 17.8 | 4.7 | 0.12 | 12.8 | 23.4 | −0.073 | −0.263 | |
| PEX | Control | 1.4 ± 0.51 | 8.3 ± 0.42 | ** | 5.1 | 11.2 | 0.66 | −4.4 | 13.2 | −0.036 | 0.105 |
| Cold | −5.5 ± 1.00 | 3.9 ± 0.32 | ** | -1.6 | 20.3 | −2.67 | −13.8 | 7.8 | 0.081 | −0.420 | |
| Treatment | Trait a | DH | CL | PL | PEX | SFF | SFG |
|---|---|---|---|---|---|---|---|
| Control | DH | 1 | ns | ns | −0.177 ** | ns | −0.213 ** |
| CL | 1 | 0.211 ** | 0.455 ** | 0.149 ** | ns | ||
| PL | 1 | 0.143 ** | 0.120 * | ns | |||
| PEX | 1 | 0.206 ** | ns | ||||
| SFF | 1 | 0.363 ** | |||||
| SFG | 1 | ||||||
| Cold | DH | 1 | −0.259 ** | −0.111 * | −0.241 ** | 0.122 * | ns |
| CL | 1 | 0.409 ** | 0.615 ** | 0.430 ** | ns | ||
| PL | 1 | 0.399 ** | 0.195 ** | ns | |||
| PEX | 0.518 ** | 0.124 * | |||||
| SFF | 1 | 0.116 * | |||||
| SFG | 1 |
| Chr. | No. of Tested Markers | No. of Polymorphic Markers a | % Polymorphism | No. of Markers Used for Map Construction b | Chromosome Length (cM) | Average Distance c (cM) | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| SSR | SNP | SSR | SNP | SSR | SNP | SSR | SNP | Sum | |||
| 1 | 83 | 44 | 63 | 34 | 75.9 | 77.3 | 16 | 20 | 36 | 190.0 | 5.3 |
| 2 | 80 | 37 | 32 | 32 | 40.0 | 86.5 | 19 | 15 | 34 | 159.0 | 4.7 |
| 3 | 68 | 41 | 43 | 23 | 63.2 | 56.1 | 21 | 19 | 40 | 182.5 | 4.6 |
| 4 | 50 | 35 | 28 | 20 | 56.0 | 57.1 | 12 | 10 | 22 | 138.4 | 6.3 |
| 5 | 72 | 28 | 28 | 13 | 38.9 | 46.4 | 11 | 8 | 19 | 137.3 | 7.2 |
| 6 | 36 | 35 | 27 | 20 | 75.0 | 57.1 | 10 | 16 | 26 | 110.3 | 4.2 |
| 7 | 44 | 30 | 37 | 26 | 84.1 | 86.7 | 12 | 18 | 30 | 120.9 | 4.0 |
| 8 | 54 | 29 | 41 | 21 | 75.9 | 72.4 | 15 | 11 | 26 | 137.8 | 5.3 |
| 9 | 40 | 23 | 36 | 18 | 90.0 | 78.3 | 14 | 12 | 26 | 85.4 | 3.3 |
| 10 | 38 | 24 | 26 | 14 | 68.4 | 58.3 | 9 | 11 | 20 | 75.1 | 3.8 |
| 11 | 53 | 30 | 34 | 18 | 64.2 | 60.0 | 16 | 11 | 27 | 108.3 | 4.0 |
| 12 | 53 | 28 | 29 | 15 | 54.7 | 53.6 | 13 | 8 | 21 | 119.0 | 5.7 |
| Sum(Mean) | 671 | 384 | 424 | 254 | (65.5) | (65.8) | 168 | 159 | 327 | 1564.0 | (4.9) |
| Evaluation Method | Trait a | QTLs | Chr. | Marker Interval | Position (Mb) b | Position (cM) | LOD c | R2 d | Additive Effect e | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Field | SFF | qSFF1-1 | 1 | id1015417 | ~ | RM3440 | 26.3 | ~ | 27.2 | 120 | 3.5 | 6.6 | 6.7 |
| qSFF1-2 | 1 | id1024323 | ~ | RM3602 | 38.3 | ~ | 39.0 | 160 | 5.6 | 10.6 | −8.5 | ||
| qSFF6 | 6 | id6004563 | ~ | id6005608 | 7.1 | ~ | 8.7 | 39 | 3.4 | 6.5 | −12.9 | ||
| qSFF9 | 9 | RM3787 | ~ | id9007180 | 20.0 | ~ | 20.8 | 75 | 3.3 | 6.3 | −6.5 | ||
| Greenhouse | SFG | qSFG2 | 2 | RM1234 | ~ | ud2000761 | 11.3 | ~ | 14.2 | 67 | 2.5 | 5.7 | −3.7 |
| qSFG6 | 6 | id6006537 | ~ | id6007220 | 10.6 | ~ | 11.4 | 43 | 5.2 | 15.1 | −8.5 | ||
| qSFG9 | 9 | id9007180 | ~ | RM1553 | 20.8 | ~ | 21.0 | 76 | 2.6 | 6.7 | −4.0 | ||
| Trait a | QTLs | Chr. | Marker Interval | Position (Mb) b | Position (cM) | LOD c | R2 d | Additive Effect e | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DH | qDH1 | 1 | id1022407 | ~ | id1024323 | 35.5 | ~ | 38.3 | 159 | 9.0 | 17.3 | 2.8 |
| qDH3 | 3 | RM8269 | ~ | fd10 | 31.4 | ~ | 31.5 | 159 | 5.0 | 9.2 | 2.1 | |
| qDH6 | 6 | id6004563 | ~ | id6005608 | 7.1 | ~ | 8.7 | 39 | 11.2 | 24.8 | −6.7 | |
| CL | qCL1-1 | 1 | id1022407 | ~ | id1024323 | 35.5 | ~ | 38.3 | 159 | 17.4 | 37.5 | −6.8 |
| qCL1-2 | 1 | id1025455 | ~ | RM5362 | 40.0 | ~ | 41.1 | 174 | 5.8 | 9.9 | −3.5 | |
| qCL3 | 3 | id3010849 | ~ | RM3513 | 24.4 | ~ | 25.1 | 116 | 6.7 | 13.6 | 4.2 | |
| qCL10 | 10 | id10004500 | ~ | RM1375 | 16.1 | ~ | 16.7 | 42 | 2.9 | 4.8 | −2.5 | |
| PL | qPL1-1 | 1 | id1016790 | ~ | RM3336 | 28.6 | ~ | 28.6 | 124 | 5.6 | 6.5 | 0.7 |
| qPL1-2 | 1 | RM3602 | ~ | id1025455 | 39.0 | ~ | 40.0 | 173 | 8.5 | 10.8 | −0.9 | |
| qPL3-1 | 3 | id3005194 | ~ | id3005824 | 10.1 | ~ | 11.1 | 50 | 2.9 | 3.3 | 0.5 | |
| qPL3-2 | 3 | id3013192 | ~ | id3013308 | 28.3 | ~ | 28.5 | 134 | 8.9 | 10.9 | 0.9 | |
| qPL8-1 | 8 | wd8002449 | ~ | RM3395 | 10.3 | ~ | 13.7 | 77 | 5.2 | 6.0 | −0.7 | |
| qPL8-2 | 8 | id8007764 | ~ | RM3840 | 27.8 | ~ | 27.9 | 137 | 6.7 | 8.1 | −0.8 | |
| qPL10 | 10 | id10003260 | ~ | RM5689 | 12.1 | ~ | 13.6 | 23 | 6.0 | 7.3 | −0.7 | |
| PEX | qPEX1 | 1 | id1024323 | ~ | RM3602 | 38.3 | ~ | 39.0 | 160 | 11.5 | 25.1 | −2.3 |
| qPEX7 | 7 | id7005423 | ~ | RM3555 | 27.6 | ~ | 27.9 | 120 | 4.9 | 9.8 | 1.4 | |
| qPEX8 | 8 | RM8264 | ~ | id8005688 | 19.8 | ~ | 20.8 | 94 | 3.3 | 7.1 | −1.2 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, J.-M.; Mo, Y.; Hyun, U.-J.; Jeung, J.-U. Identification of Quantitative Trait Loci for Spikelet Fertility at the Booting Stage in Rice (Oryza sativa L.) under Different Low-Temperature Conditions. Agronomy 2020, 10, 1225. https://doi.org/10.3390/agronomy10091225
Jeong J-M, Mo Y, Hyun U-J, Jeung J-U. Identification of Quantitative Trait Loci for Spikelet Fertility at the Booting Stage in Rice (Oryza sativa L.) under Different Low-Temperature Conditions. Agronomy. 2020; 10(9):1225. https://doi.org/10.3390/agronomy10091225
Chicago/Turabian StyleJeong, Jong-Min, Youngjun Mo, Ung-Jo Hyun, and Ji-Ung Jeung. 2020. "Identification of Quantitative Trait Loci for Spikelet Fertility at the Booting Stage in Rice (Oryza sativa L.) under Different Low-Temperature Conditions" Agronomy 10, no. 9: 1225. https://doi.org/10.3390/agronomy10091225