Response to the Cold Stress Signaling of the Tea Plant (Camellia sinensis) Elicited by Chitosan Oligosaccharide


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Cold Treatments
2.2. Physiological Response Assay
2.3. cDNA Library Construction and Sequencing
2.4. RNA-Seq Data Analysis
2.5. Identification of Differentially Expressed Genes
2.6. Gene Ontology and KEGG Pathway Analysis
2.7. Quantitative RT-PCR (qRT-PCR) Analysis
2.8. Statistical Analysis
3. Results
3.1. Physiological Parameter Response to a Low Temperature
3.2. Transcriptome Sequencing and Assembly
3.3. Differentially Expressed Genes Analysis
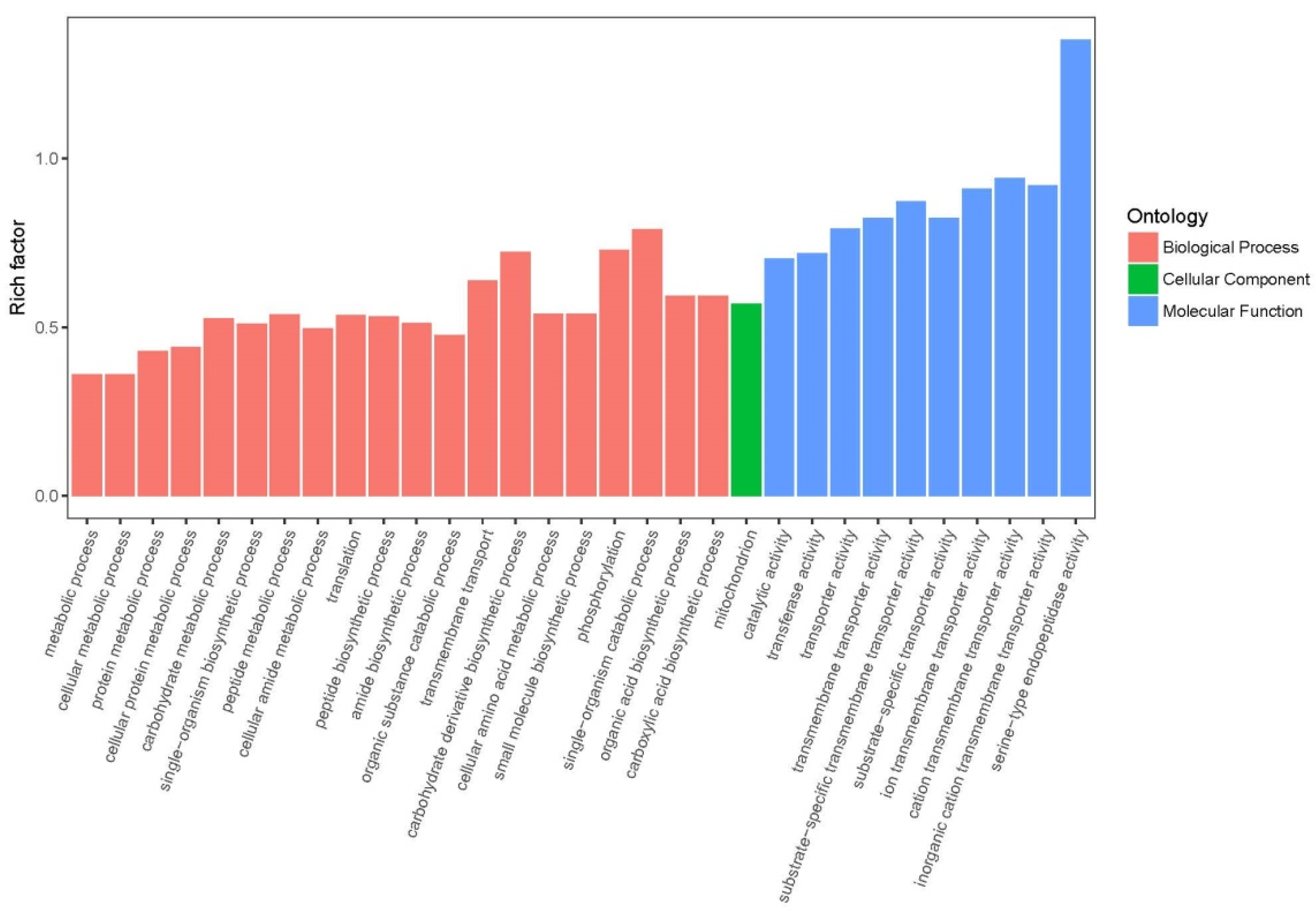
3.4. Gene Ontology (GO) Annotation
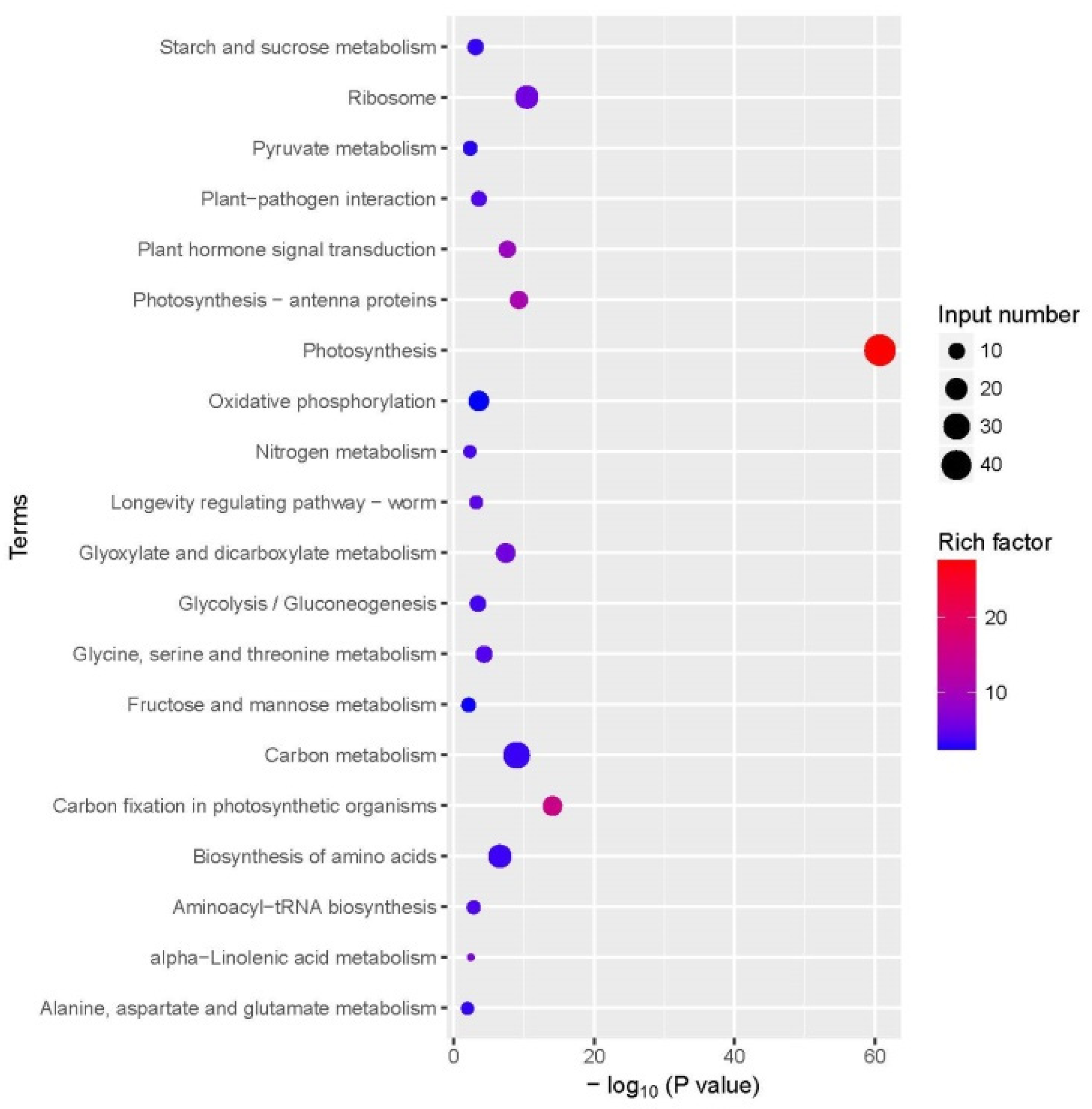
3.5. Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathway Annotation
3.6. qRT-PCR Validation of Differentially Expressed Transcripts from RNA-Seq
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Singh, H.R.; Hazarika, P. Biotechnological Approaches for Tea Improvement. In Biotechnological Progress and Beverage Consumption; Academic Press: Cambridge, MA, USA, 2020; pp. 111–148. [Google Scholar]
- FAOSTAT-Food and Agriculture Organization of the United Nations Statistics Division. Available online: http://faostat3.fao.org/home/E (accessed on 31 December 2018).
- Ma, C.L.; Chen, L.; Wang, X.; Jin, J.Q.; Ma, J.Q.; Yao, M.Z. Differential expression analysis of different albescent stages of ‘Anji Baicha’ (Camellia sinensis (L.) O. Kuntze) using cDNA microarray. Sci. Hortic. 2012, 148, 246–254. [Google Scholar] [CrossRef]
- Du, Y.Y.; Liang, Y.R.; Wang, H.; Wang, K.R.; Lu, J.L.; Zhang, G.H.; Lin, W.P.; Li, M.; Fang, Q.Y. A study on the chemical composition of albino tea cultivars. J. Hort. Sci. Biotechnol. 2006, 81, 809–812. [Google Scholar] [CrossRef]
- Cheng, H.; Li, S.F.; Chen, M.; Yu, F.L.; Yan, J.; Liu, Y.M.; Chen, L.A. Physiological and biochemical essence of the extraordinary characters of Anji Baicha. J. Tea Sci. 1999, 19, 87–92. [Google Scholar]
- Du, Y.Y.; Chen, H.; Zhong, W.L.; Wu, L.Y.; Ye, J.H.; Lin, C.; Zheng, X.Q.; Lu, J.L.; Liang, Y.R. Effect of temperature on accumulation of chlorophylls and leaf ultrastructure of low temperature induced albino tea plant. Afr. J. Biotechnol. 2008, 7, 1881–1885. [Google Scholar] [CrossRef]
- Feng, L.; Gao, M.J.; Hou, R.Y.; Hu, X.Y.; Zhang, L.; Wan, X.C.; Wei, S. Determination of quality constituents in the young leaves of albino tea cultivars. Food Chem. 2014, 155, 98–104. [Google Scholar] [CrossRef]
- Wei, K.; Wang, L.Y.; Zhou, J.; He, W.; Zeng, J.M.; Jiang, Y.W.; Cheng, H. Comparison of catechins and purine alkaloids in albino and normal green tea cultivars (Camellia sinensis L.) by HPLC. Food Chem. 2012, 130, 720–724. [Google Scholar] [CrossRef]
- Shen, J.; Wang, Y.; Chen, C.; Ding, Z.; Hu, J.; Zheng, C.; Li, Y. Metabolite profiling of tea (Camellia sinensis L.) leaves in winter. Sci. Hortic. 2015, 192, 1–9. [Google Scholar] [CrossRef]
- Wang, L.; Cao, H.; Qian, W.; Yao, L.; Hao, X.; Li, N.; Yang, Y.; Wang, X. Identification of a novel bZIP transcription factor in Camellia sinensis as a negative regulator of freezing tolerance in transgenic Arabidopsis. Ann. Bot. 2017, 119, 1195–1209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.W.; Li, T.Y.; Wang, Q.S.; LeCompte, J.; Harkess, R.L.; Bi, G.H. Screening tea cultivars for novel climates: Plant growth and leaf quality of Camellia sinensis cultivars grown in Mississippi, United States. Front. Plant Sci. 2020, 11, 280. [Google Scholar] [CrossRef]
- Li, X.; Ahammed, G.; Li, Z.; Zhang, L.; Wei, J.; Yan, P.; Zhang, L.; Han, W. Freezing stress deteriorates tea quality of new flush by inducing photosynthetic inhibition and oxidative stress in mature leaves. Sci. Hortic. 2018, 230, 155–160. [Google Scholar] [CrossRef]
- Wang, X.C.; Zhao, Q.Y.; Ma, C.L.; Zhang, Z.H.; Cao, H.L.; Kong, Y.M.; Yue, C.; Hao, X.Y.; Chen, L.; Ma, J.Q.; et al. Global transcriptome profiles of Camellia sinensis during cold acclimation. BMC Genom. 2013, 14, 415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, Y.; Ma, Q.; Zhu, Z.; Cui, Q.; Chen, C.; Chen, X.; Fang, W.; Li, X. Functional analysis of CsCBF3 transcription factor in tea plant (Camellia sinensis) under cold stress. Plant Growth Regul. 2016, 80, 335–343. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhu, X.; Chen, X.; Song, C.; Zou, Z.; Wang, Z.; Wang, M.; Fang, W.; Li, X. Identification and characterization of cold-responsive microRNAs in tea plant (Camellia sinensis) and their targets using high-throughput sequencing and degradome analysis. BMC Plant Biol. 2014, 14, 271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.J.; Zheng, L.Y.; Wang, X.C. Effect of cold acclimation and ABA on cold hardiness contents of proline in tea plants. J. Tea Sci. 2004, 24, 177–182. [Google Scholar]
- Li, Y.Y.; Wang, X.W.; Ban, Q.Y.; Zhu, X.X.; Jiang, C.J.; Wei, C.L.; Bennetzen, J.L. Comparative transcriptomic analysis reveals gene expression associated with cold adaptation in the tea plant Camellia sinensis. BMC Genom. 2019, 20, 624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ban, Q.Y.; Wang, X.W.; Pan, C.; Wang, Y.W.; Kong, L.; Jiang, H.G.; Xu, Y.Q.; Wang, W.Z.; Pan, Y.T.; Li, Y.Y.; et al. Comparative analysis of the response and gene regulation in cold resistant and susceptible tea plants. PLoS ONE 2017, 12, e0188514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.H.; Yang, Y.Q.; Sun, K.; Chen, Y.; Chen, X.; Li, X.H. Exogenous Melatonin Enhances Cold, Salt and Drought Stress Tolerance by Improving Antioxidant Defense in Tea Plant (Camellia sinensis (L.) O. Kuntze). Molecules 2019, 24, 1826. [Google Scholar] [CrossRef] [Green Version]
- Cabrera, J.; Wégria, G.; Onderwater, R.; González, G.; Nápoles, M.; Falcón-Rodríguez, A.; Costales, D.; Rogers, H.; Diosdado, E.; González, S.; et al. Practical use of oligosaccharins in agriculture. Acta. Hortic. 2013, 1009, 195–212. [Google Scholar] [CrossRef]
- Kim, S.; Rajapakse, N. Enzymatic production and biological activities of chitosan oligosaccharides (COS): A review. Carbohydr. Polym. 2005, 62, 357–368. [Google Scholar] [CrossRef]
- Yin, H.; Du, Y.G.; Zhang, J.Z. Low molecular weight and oligomeric chitosans and their bioactivities. Curr. Top. Med. Chem. 2009, 9, 1546–1559. [Google Scholar] [CrossRef]
- Wang, M.Y.; Chen, Y.C.; Zhang, R.; Wang, W.X.; Zhao, X.M.; Du, Y.G.; Yin, H. Effects of chitosan oligosaccharides on the yield components and production quality of different wheat cultivars (Triticum aestivum L.) in Northwest China. Field Crop. Res. 2015, 172, 11–20. [Google Scholar] [CrossRef]
- Chatelain, P.G.; Pintado, M.E.; Vasconcelos, M.W. Evaluation of chitooligosaccharide application on mineral accumulation and plant growth in Phaseolus vulgaris. Plant Sci. 2014, 215, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Zou, P.; Tian, X.Y.; Dong, B.; Zhang, C.S. Size effects of chitooligomers with certain degrees of polymerization on the chilling tolerance of wheat seedlings. Carbohydr. Polym. 2017, 160, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.J.; Li, Y.Y.; Yu, C.M.; Wang, Y.; Li, X.M.; Li, N.; Chen, Q.; Bu, N. Alleviation of exogenous oligochitosan on wheat seedlings growth under salt stress. Protoplasma 2012, 249, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Zou, P.; Li, K.C.; Liu, S.; Xing, R.; Qin, Y.K.; Yu, H.K.; Zhao, M.M.; Li, P.C. Effect of chitooligosaccharides with different degrees of acetylation on wheat seedlings under salt stress. Carbohydr. Polym. 2015, 126, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Yang, A.M.; Yu, L.; Chen, Z.; Zhang, S.X.; Shi, J.; Zhao, X.Z.; Yang, Y.Y.; Hu, D.Y.; Song, B.A. Label-free quantitative proteomic analysis of chitosan oligosaccharide-treated rice infected with southern rice black-streaked dwarf virus. Viruses 2017, 9, 115. [Google Scholar] [CrossRef]
- Jia, X.C.; Meng, Q.S.; Zeng, H.H.; Wang, W.X.; Yin, H. Chitosan oligosaccharide induces resistance to tobacco mosaic virus in Arabidopsis via the salicylic acid-mediated signalling pathway. Sci. Rep. 2016, 6, 26144–26155. [Google Scholar] [CrossRef]
- Ahmed, K.; Khan, M.; Siddiqui, H.; Jahan, A. Chitosan and its oligosaccharides, a promising option for sustainable crop production-a review. Carbohydr. Polym. 2020, 227, 115331. [Google Scholar] [CrossRef]
- Cheplick, S.; Sarkar, D.; Bhowmik, P.C.; Shetty, K. Improved resilience and metabolic response of transplanted blackberry plugs using chitosan oligosaccharide elicitor treatment. Can. J. Plant Sci. 2017, 98, 717–731. [Google Scholar] [CrossRef] [Green Version]
- Zeng, D.F.; Luo, X.R. Physiological effects of chitosan coating on wheat growth and activities of protective enzyme with drought tolerance. Open J. Soil Sci. 2012, 2, 282–288. [Google Scholar] [CrossRef] [Green Version]
- Hu, Z.H.; Tang, B.; Wu, Q.; Zheng, J.; Leng, P.S.; Zhang, K.Z. Transcriptome sequencing analysis reveals a difference in monoterpene biosynthesis between scented Lilium ‘Siberia’ and unscented Lilium ‘Novano’. Front. Plant Sci. 2017, 8, 1351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.D.; Xin, H.H.; Wang, M.L.; Ma, Q.P.; Wang, L.; Kaleri, N.A.; Wang, Y.H.; Li, X.H. Transcriptomic analysis reveals the molecular mechanisms of drought-stress-induced decreases in Camellia sinensis leaf quality. Front. Plant Sci. 2016, 7, 385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, X.Y.; Tang, H.; Wang, B.; Yue, C.; Wang, L.; Zeng, J.M.; Yang, Y.J.; Wang, X.C. Integrative transcriptional and metabolic analyses provide insights into cold spell response mechanisms in young shoots of the tea plant. Tree Physiol. 2018, 38, 1655–1671. [Google Scholar] [CrossRef] [PubMed]
- Paul, A.; Jha, A.; Bhardwaj, S.; Singh, S.; Shankar, R.; Kumar, S. RNA-seq-mediated transcriptome analysis of actively growing and winter dormant shoots identifies non-deciduous habit of evergreen tree tea during winters. Sci. Rep. 2014, 4, 5932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.F.; Xu, Y.X.; Ma, J.Q.; Jin, J.Q.; Huang, D.J.; Yao, M.Z.; Ma, C.L.; Chen, L. Biochemical and transcriptomic analyses reveal different metabolite biosynthesis profiles among three color and developmental stages in ‘Anji Baicha’ (Camellia sinensis). BMC Plant Biol. 2016, 16, 195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, C.L.; Yang, H.; Wang, S.B.; Zhao, J.; Liu, C.; Gao, L.P.; Xia, E.H.; Lu, Y.; Tai, Y.L.; She, G.B.; et al. Draft genome sequence of Camellia sinensis var. sinensis provides insights into the evolution of the tea genome and tea quality. Proc. Natl. Acad. Sci. USA 2018, 115, 201719622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, Y.; Smyth, G.; Shi, W. featureCounts: An efficient general-purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.; McCarthy, D.; Smyth, G. edgeR: A bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Xie, C.; Mao, X.; Huang, J.; Ding, Y.; Wu, J.; Dong, S.; Kong, L.; Gao, G.; Li, C.; Wei, L. KOBAS 2.0: A web server for annotation and identification of enriched pathways and diseases. Nucleic Acids Res. 2011, 39, 316–322. [Google Scholar] [CrossRef] [Green Version]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [Green Version]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.N.; Huang, M.; Cao, F.B.; Pardha-Saradhi, P.; Zou, Y.B. Urea application promotes amino acid metabolism and membrane lipid peroxidation in Azolla. PLoS ONE 2017, 12, e0185230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, C.Z.; Zhu, C.; Fu, H.F.; Li, X.Z.; Chen, L.; Lin, Y.L.; Lai, Z.X.; Guo, Y.Q. Genome-wide investigation of superoxide dismutase (SOD) gene family and their regulatory miRNAs reveal the involvement in abiotic stress and hormone response in tea plant (Camellia sinensis). PLoS ONE 2019, 14, e0223609. [Google Scholar] [CrossRef] [PubMed]
- Zou, P.; Li, K.C.; Liu, S.; He, X.F.; Xing, R.; Zhang, X.Q.; Li, P.C. Effect of sulfated chitooligosaccharides on wheat seedlings (Triticum aestivum L.) under saltstress. J. Agric. Food Chem. 2016, 64, 2815–2821. [Google Scholar] [CrossRef] [PubMed]
- Anwar, A.; Yan, Y.; Liu, Y.; Li, Y.; Yu, X. 5-aminolevulinic acid improves nutrient uptake and endogenous hormone accumulation, enhancing low-temperature stress tolerance in cucumbers. Int. J. Mol. Sci. 2018, 19, 3379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suorsa, M.; Sirpio, S.; Allahverdiyeva, Y.; Paakkarinen, V.; Mamedov, F.; Styring, S.; Aro, E.M. PsbR, a missing link in the assembly of the oxygen-evolving complex of plant photosystem II. J. Biol. Chem. 2006, 281, 145–150. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.Q.; Li, K.C.; Xing, R.; Liu, S.; Chen, X.L.; Yang, H.Y.; Li, P.C. MiRNA and mRNA expression profiles reveal insight into the chitosan-mediated regulation of plant growth. J. Agric. Food Chem. 2018, 66, 3810–3822. [Google Scholar] [CrossRef] [PubMed]
- Li, X.W.; Zhu, Y.X.; Chen, C.Y.; Geng, Z.J.; Li, X.Y.; Ye, T.T.; Mao, X.N.; Du, F. Cloning and characterization of two chlorophyll A/B binding protein genes and analysis of their gene family in Camellia sinensis. Sci. Rep. 2020, 10, 4602. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.F.; Zhao, H.; Guo, F.; Shi, X.P.; Ye, C.; Y, P.X.; Liu, B.Y.; Ni, D.J. Transcriptomic analysis reveals mechanism of light-sensitive albinism in tea plant Camellia sinensis ‘Huangjinju’. BMC Plant Biol. 2020, 20, 216. [Google Scholar] [CrossRef]
- Sharma, A.; Kumar, V.; Shahzad, B.; Ramakrishnan, M.; Sidhu, G.P.S.; Bali, A.S.; Handa, N.; Kapoor, D.; Yadav, P.; Khanna, K.; et al. Photosynthetic response of plants under different abiotic stresses: A review. J. Plant Growth Regul. 2020, 39, 509–531. [Google Scholar] [CrossRef]
- Chamnanmanoontham, N.; Pongprayoon, W.; Pichayangkura, R.; Roytrakul, S.; Chadchawan, S. Chitosan enhances rice seedling growth via gene expression network between nucleus and chloroplast. Plant Growth Regul. 2015, 75, 101–114. [Google Scholar] [CrossRef]
- Abel, S.; Nguyen, M.D.; Theologis, A. The PS-IAA4/5-like family of early inducible mRNAs in Arabidopsis thaliana. J. Mol. Biol. 1995, 251, 533–549. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Yue, R.; Tao, S.; Yang, Y.; Zhang, L.; Xu, M.; Wang, H.; Shen, C. Genome-wide identification, expression analysis of auxin-responsive GH3 family genes in maize (Zea mays L.) under abiotic stresses. J. Integr. Plant Biol. 2015, 57, 783–795. [Google Scholar] [CrossRef] [PubMed]
- Solano, R.; Ecker, J.R. Ethylene gas: Perception, signaling and response. Curr. Opin. Plant Biol. 1998, 1, 393–398. [Google Scholar] [CrossRef]
- Chomczynski, P.; Sacchi, N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef]
- La Camera, S.; Gouzerh, G.; Dhondt, S.; Hoffmann, L.; Fritig, B.; Legrand, M.; Heitz, T. Metabolic reprogramming in plant innate immunity: The contributions of phenylpropanoid and oxylipin pathways. Immunol. Rev. 2004, 198, 267–284. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | ConT3_1 | ConT3_2 | ConT3_3 | TreT3_1 | TreT3_2 | TreT3_3 |
|---|---|---|---|---|---|---|
| Raw reads | 55,965,032 | 56,476,808 | 66,044,722 | 57,864,054 | 67,743,104 | 65,453,870 |
| Clean reads | 52,619,470 | 53,061,678 | 62,155,236 | 54,555,936 | 63,422,124 | 61,416,118 |
| Q30 (%) | 98.45 | 98.45 | 98.70 | 98.65 | 98.55 | 98.45 |
| GC content (%) | 46.60 | 46.46 | 46.63 | 46.82 | 46.83 | 47.21 |
| Total reads | 44,163,580 | 43,980,650 | 52,344,630 | 45,455,332 | 52,920,720 | 51,188,834 |
| Total mapped | 41,828,592 (94.71%) | 41,644,005 (94.69%) | 49,676,907 (94.90%) | 43,274,546 (95.20%) | 50,292,573 (95.03%) | 48,551,418 (94.85%) |
| Unique mapped | 38,522,223 (92.10%) | 38,095,551 (91.48%) | 45,663,412 (91.92%) | 39,232,023 (90.66%) | 45,576,183 (90.62%) | 42,734,470 (88.02%) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Zhang, Q.; Ou, L.; Ji, D.; Liu, T.; Lan, R.; Li, X.; Jin, L. Response to the Cold Stress Signaling of the Tea Plant (Camellia sinensis) Elicited by Chitosan Oligosaccharide. Agronomy 2020, 10, 915. https://doi.org/10.3390/agronomy10060915
Li Y, Zhang Q, Ou L, Ji D, Liu T, Lan R, Li X, Jin L. Response to the Cold Stress Signaling of the Tea Plant (Camellia sinensis) Elicited by Chitosan Oligosaccharide. Agronomy. 2020; 10(6):915. https://doi.org/10.3390/agronomy10060915
Chicago/Turabian StyleLi, Yingying, Qiuqiu Zhang, Lina Ou, Dezhong Ji, Tao Liu, Rongmeng Lan, Xiangyang Li, and Linhong Jin. 2020. "Response to the Cold Stress Signaling of the Tea Plant (Camellia sinensis) Elicited by Chitosan Oligosaccharide" Agronomy 10, no. 6: 915. https://doi.org/10.3390/agronomy10060915