Metabolomic Analysis for Compositional Differences of Bush Tea (Athrixia phylicoides DC.) Subjected to Seasonal Dynamics


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site
2.2. Experimental Design and Treatments
2.3. Sample Preparation
2.4. LC-MS/MS Triple Quadrupole Mass Spectrometer
2.5. 1H NMR Data Acquisition
2.6. Statistical Analysis
3. Results
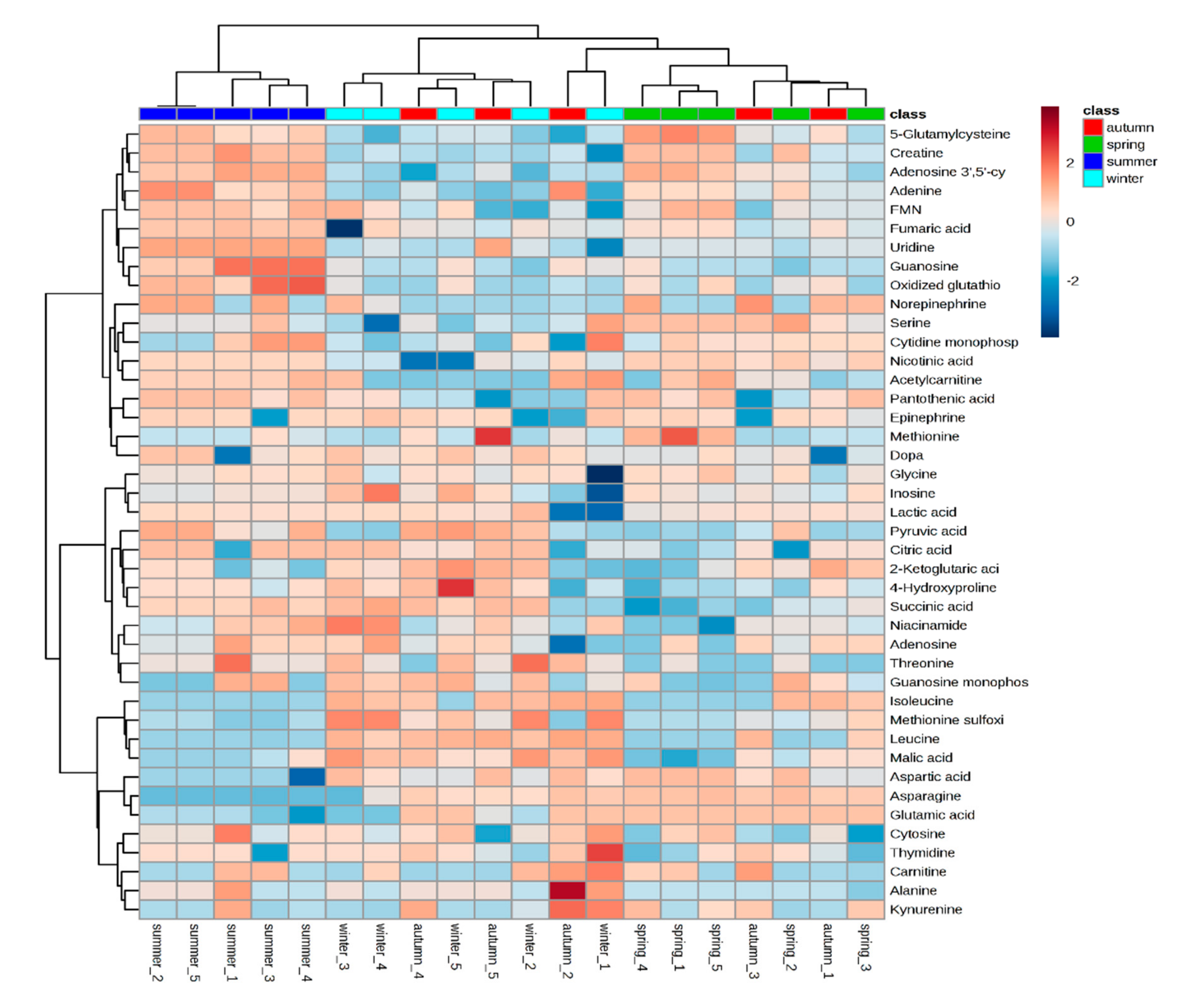
3.1. Seasonal Variation on Bush Tea Primary Metabolites Using LC-MS/MS
3.2. The Effect of Harvest Season on Bush Tea Metabolome Using 1H NMR
3.3. Annotation of Bush Tea Metabolites from 1H NMR Spectrum
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nchabeleng, L.; Mariga, I.K.; Ngezimana, W.; Mudau, F.N. Bush tea (Athrixia phylicoides DC.) success stories in South Africa: A review. ESci. J. Crop Prod. 2013, 2, 37–43. [Google Scholar]
- McGaw, L.J.; Bagla, V.; Steenkamp, P.A.; Fouche, G.; Olivier, J.; Eloff, J.N.; Myer, M.S. Antifungal and antibacterial activity and chemical composition of polar and non-polar extracts of Athrixia phylicoides determined using bioautography and HPLC. BMC Complement. Altern. Med. 2013, 13, 356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malongane, F.; McGaw, L.J.; Nyoni, H.; Mudau, F. Metabolic profiling of four South African herbal teas using high resolution liquid chromatography-mass spectrometry and nuclear magnetic resonance. Food Chem. 2018, 257, 90–100. [Google Scholar] [CrossRef]
- Lerotholi, L.; Chaudhary, S.; Combrinck, S.; Viljoen, A.M. Bush tea (Athrixia phylicoides): A review of the traditional uses, bioactivity and phytochemistry. South Afr. J. Bot. 2017, 110, 4–17. [Google Scholar] [CrossRef]
- Tshivhandekano, I.; Ntushelo, K.; Ngezimana, W.; Tshikalange, T.E.; Mudau, F. Chemical compositions and antimicrobial activities of Athrixia phylicoides DC. (bush tea), Monsonia burkeana (special tea) and synergistic effects of both combined herbal teas. Asian Pac. J. Trop. Med. 2014, 7, 448–453. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Gao, L.; Shan, Y.; Liu, Y.; Tian, Y.; Xia, T. Influence of shade on flavonoid biosynthesis in tea (Camellia sinensis (L.) O. Kuntze). Sci. Hortic. 2012, 141, 7–16. [Google Scholar] [CrossRef]
- Mudau, F.N.; Ngele, A.; Mashela, P.W.; Soundy, P. Seasonal variation of tannin content in wild bush tea. Med. Aromat. Plant Sci. Biotechnol. 2007, 1, 74–76. [Google Scholar]
- Wei, K.; Wang, L.; Zhou, J.; He, W.; Zeng, J.-M.; Jiang, Y.-W.; Cheng, H. Comparison of catechins and purine alkaloids in albino and normal green tea cultivars (Camellia sinensis L.) by HPLC. Food Chem. 2012, 130, 720–724. [Google Scholar] [CrossRef]
- Yao, L.; Caffin, N.; D’Arcy, B.; Jiang, Y.; Shi, J.; Singanusong, R.; Liu, X.; Datta, N.; Kakuda, Y.; Xu, Y. Seasonal Variations of Phenolic Compounds in Australia-Grown Tea (Camellia sinensis). J. Agric. Food Chem. 2005, 53, 6477–6483. [Google Scholar] [CrossRef]
- Selmar, D.; Kleinwächter, M. Stress Enhances the Synthesis of Secondary Plant Products: The Impact of Stress-Related Over-Reduction on the Accumulation of Natural Products. Plant Cell Physiol. 2013, 54, 817–826. [Google Scholar] [CrossRef]
- Mudau, F.; Mogotlane, I.; Mashela, P.; Soundy, P. Seasonal Variation of Total Antioxidant Contents of Wild Bush Tea. Acta Hortic. 2008, 802, 273–276. [Google Scholar] [CrossRef]
- Morison, J.I.L.; Lawlor, D.W. Interactions between increasing CO2concentration and temperature on plant growth. Plant Cell Environ. 1999, 22, 659–682. [Google Scholar] [CrossRef] [Green Version]
- Grace, S.C.; Logan, B.A. Energy dissipation and radical scavenging by the plant phenylpropanoid pathway. Philos. Trans. R. Soc. B Boil. Sci. 2000, 355, 1499–1510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kranner, I.; Beckett, R.; Wornik, S.; Zorn, M.; Pfeifhofer, H.W. Revival of a resurrection plant correlates with its antioxidant status. Plant J. 2002, 31, 13–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kale, V.S. Variable rates of primary and secondary metabolites during different seasons and physiological stages in Convolvulus, Withania and Datura. Asian J. Exp. Biol. Sci. 2010, 10, 50–53. [Google Scholar]
- Szakiel, A.; Pączkowski, C.; Henry, M. Influence of environmental abiotic factors on the content of saponins in plants. Phytochem. Rev. 2010, 10, 471–491. [Google Scholar] [CrossRef]
- Lee, J.-E.; Lee, B.-J.; Hwang, J.-A.; Ko, K.-S.; Chung, J.-O.; Kim, E.-H.; Lee, S.-J.; Hong, Y.-S. Metabolic Dependence of Green Tea on Plucking Positions Revisited: A Metabolomic Study. J. Agric. Food Chem. 2011, 59, 10579–10585. [Google Scholar] [CrossRef]
- Kellogg, J.J.; Graf, T.; Paine, M.F.; McCune, J.S.; Kvalheim, O.M.; Oberlies, N.H.; Cech, N.B. Comparison of Metabolomics Approaches for Evaluating the Variability of Complex Botanical Preparations: Green Tea (Camellia sinensis) as a Case Study. J. Nat. Prod. 2017, 80, 1457–1466. [Google Scholar] [CrossRef] [Green Version]
- Vuwani, M.K.; Nkomo, M.; Ngezimana, W.; Makunga, N.P.; Mudau, F. Response of Phytochemicals in Bush Tea (Athrixia phylicoides DC.) as Influenced by Selected Micronutrients. HortScience 2017, 52, 965–971. [Google Scholar] [CrossRef]
- Zhang, Q.; Shi, Y.; Ma, L.; Yi, X.; Ruan, J. Metabolomic Analysis Using Ultra-Performance Liquid Chromatography-Quadrupole-Time of Flight Mass Spectrometry (UPLC-Q-TOF MS) Uncovers the Effects of Light Intensity and Temperature under Shading Treatments on the Metabolites in Tea. PLoS ONE 2014, 9, e112572. [Google Scholar] [CrossRef]
- Jing, J.; Shi, Y.-Z.; Zhang, Q.; Wang, J.; Ruan, J. Prediction of Chinese green tea ranking by metabolite profiling using ultra-performance liquid chromatography–quadrupole time-of-flight mass spectrometry (UPLC–Q-TOF/MS). Food Chem. 2017, 221, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Jiang, Y.; Duan, J.; Shi, J.; Xue, S.; Kakuda, Y. Variation in catechin contents in relation to quality of ‘Huang Zhi Xiang’ Oolong tea (Camellia sinensis) at various growing altitudes and seasons. Food Chem. 2010, 119, 648–652. [Google Scholar] [CrossRef]
- Lee, J.-E.; Lee, B.-J.; Chung, J.-O.; Hwang, J.-A.; Lee, S.-J.; Lee, C.-H.; Hong, Y.-S. Geographical and Climatic Dependencies of Green Tea (Camellia sinensis) Metabolites: A1H NMR-Based Metabolomics Study. J. Agric. Food Chem. 2010, 58, 10582–10589. [Google Scholar] [CrossRef]
- Lee, J.-E.; Lee, B.-J.; Chung, J.-O.; Kim, H.-N.; Kim, E.; Jung, S.; Lee, H.; Lee, S.-J.; Hong, Y.-S. Metabolomic unveiling of a diverse range of green tea (Camellia sinensis) metabolites dependent on geography. Food Chem. 2015, 174, 452–459. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Dai, W.; Lu, M.; Lv, H.; Guo, L.; Zhang, Y.; Zhu, Y.; Peng, Q.; Lin, Z. Study of the dynamic changes in the non-volatile chemical constituents of black tea during fermentation processing by a non-targeted metabolomics approach. Food Res. Int. 2016, 79, 106–113. [Google Scholar] [CrossRef]
- Mudau, F.N.; Ngezimana, W. Effect of different drying methods on chemical composition and antimicrobial activity of bush tea (Athrixia phylicoides). Int. J. Agric. Biol. 2014, 16, 1011–1014. [Google Scholar]
- Maree, J.; Viljoen, A.M. Phytochemical distinction between Pelargonium sidoides and Pelargonium reniforme—A quality control perspective. South Afr. J. Bot. 2012, 82, 83–91. [Google Scholar] [CrossRef] [Green Version]
- Selvaraj, K.; Chowdhury, R.; Bhattacharjee, C. Isolation and structural elucidation of flavonoids from aquatic fern Azolla microphylla and evaluation of free radical scavenging activity. Int. J. Pharm. Pharm. Sci. 2013, 5, 743–749. [Google Scholar]
- Georgeta SPana, P.I.; Tunde, H.; Sanda, B. The isolation and identification of rutin from pharmaceutical products. Analele Univ. Din. Oradea Fasc. Ecotoxicol. Zooteh Si Tehnol. Ind. Aliment. 2016, 16, 109–113. [Google Scholar]
- Padayachee, K. The Phytochemistry and Biological Activities of Athrixia Phylicoides. M.Sc. Thesis, University of Witwatersrand, Johannesburg, South Africa, 2011. Available online: http://wiredspace.wits.ac.za/handle/10539/10667 (accessed on 11 August 2017).
- Exarchou, V.; Troganis, A.; Gerothanassis, I.P.; Tsimidou, M.Z.; Boskou, D. Identification and quantification of caffeic and rosmarinic acid in complex plant extracts by the use of variable-temperature two-dimensional nuclear magnetic resonance spectroscopy. J. Agric. Food Chem. 2001, 49, 2–8. [Google Scholar] [CrossRef]
- Hayashi, T.; Uchida, K.; Hayashi, K.; Niwayama, S.; Morita, N. A cytotoxic flavone from Scoparia dulcis L. Chem. Pharm. Bull. 1988, 36, 4849–4851. [Google Scholar] [CrossRef] [Green Version]
- Sambandam, B.; Thiyagarajan, D.; Ayyaswamy, A.; Raman, P.; Kulasekaran, J.; Venkatasamy, H. Extraction and isolation of flavonoid quercetin from the leaves of Trigonella foenum-graecum and their anti-oxidant activity. Int. J. Pharm. Pharm. Sci. 2016, 8, 120–124. [Google Scholar]
- Lin, L.-J.; Huang, X.-B.; Lv, Z.-C. Isolation and identification of flavonoids components from Pteris vittata L. SpringerPlus 2016, 5, 1649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caligiani, A.; Palla, G.; Maietti, A.; Cirlini, M.; Brandolini, V. 1H NMR Fingerprinting of Soybean Extracts, with Emphasis on Identification and Quantification of Isoflavones. Nutrients 2010, 2, 280–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gogna, N.; Hamid, N.; Dorai, K. Metabolomic profiling of the phytomedicinal constituents of Carica papaya L. leaves and seeds by 1H NMR spectroscopy and multivariate statistical analysis. J. Pharm. Biomed. Anal. 2015, 115, 74–85. [Google Scholar] [CrossRef]
- Crawford, R.M. Seasonal differences in plant responses to flooding and anoxia. Can. J. Bot. 2003, 81, 1224–1246. [Google Scholar] [CrossRef]
- Hare, P.; Cress, W. Metabolic implications of stress-induced proline accumulation in plants. Plant Growth Regul. 1997, 21, 79–102. [Google Scholar] [CrossRef]
- Harbowy, M.E.; Balentine, D.A. Tea chemistry. Crit. Rev. Plant Sci. 1997, 16, 415–480. [Google Scholar] [CrossRef]
- Krol, A.; Amarowicz, R.; Weidner, S. The effects of cold stress on the phenolic compounds and antioxidant capacity of grapevine (Vitis vinifera L.) leaves. J. Plant Physiol. 2015, 189, 97–104. [Google Scholar] [CrossRef]
- Erturk, Y.; Ercisli, S.; Sengul, M.; Eser, Z.; Haznedar, A.; Turan, M. Seasonal variation of total phenolic, antioxidant activity and minerals in fresh tea shoots (Camellia sinensis var. sinensis). Pak. J. Pharm. Sci. 2010, 23, 69–74. [Google Scholar]
- Gololo, S.S.; Shai, L.J.; Agyei, N.M.; Mogale, M.A. Effect of seasonal changes on the quantity of phytochemicals in the leaves of three medicinal plants from Limpopo province, South Africa. J. Pharmacogn. Phytother. 2016, 8, 168–172. [Google Scholar]
- Mudau, F.; Soundy, P.; Du Toit, E.; Olivier, J. Variation in polyphenolic content of Athrixia phylicoides (L.) (bush tea) leaves with season and nitrogen application. South Afr. J. Bot. 2006, 72, 398–402. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Season | Autumn | Winter | Spring | Summer |
|---|---|---|---|---|
| (Peak Intensities) | ||||
| Essential amino acids | ||||
| Threonine | 3.29 × 105 ± 0.42 × 105 b | 3.83 × 105 ± 0.75 × 105 b | 8.73 × 105 ± 1.34 × 105 a | 8.65 × 105 ± 0.41 × 105 a |
| Methionine sulfoxide | 1.45 × 105 ± 0.28 × 105 c | 2.80 × 105 ± 0.47 × 105 a | 2.20 × 105 ± 0.13 × 105 b | 1.54 × 105 ± 0.07 × 105 c |
| Isoleucine | 3.24 × 104 ± 1.13 × 104 a | 3.58 ×104 ± 0.93 ×104 a | 2.01 ×104 ± 0.13 × 104 b | 0.0 ± 0.0 c |
| Leucine | 3.49 × 104 ± 1.28 × 104 b | 3.85 ×104 ± 0.39 × 104 a | 0.59 ×104 ± 0.04 × 104 c | 0.0 ± 0.0 d |
| Conditional and non-essential amino acids | ||||
| Asparagine | 0.37 × 106 ± 0.06 × 106 b | 0.11 × 106 ± 0.02 × 106 c | 1.51 × 106 ± 0.22 × 106 a | 0.0 ± 0.0 d |
| Aspartic acid | 1.46 × 105 ± 0.19 × 105 bc | 1.27 × 105 ± 012 × 105 c | 3.23 × 105 ± 0.43 × 105 a | 1.65 × 105 ± 0.09 × 105 b |
| Serine | 1.46 × 105 ± 0.21 × 105 c | 0.99 × 105 ± 0.09 × 105 d | 3.82 × 105 ± 0.41 × 105 a | 2.06 × 105 ± 0.12 × 105 b |
| Alanine | 5.16 × 105 ± 0.77 × 105 c | 4.42 × 105 ± 0.61 × 105 c | 9.91 × 105 ± 1.34 × 105 b | 11.93 × 105 ± 2.47 × 105 a |
| Glutamic acid | 1.42 × 106 ± 0.21 × 106 c | 1.32 × 106 ± 0.16 × 106 c | 2.46 × 106 ± 0.27 × 106 a | 1.69 × 106 ± 0.14 × 106 b |
| Glycine | 0.99 × 104 ± 0.09 × 104 c | 1.16 × 104 ± 0.34 × 104 c | 2.57 × 104 ± 0.29 × 104 a | 1.63 × 104 ± 0.58 × 104 b |
| Proline | 3.48 × 107 ± 0.07 × 107 b | 2.75 × 107 ± 0.47 × 107 c | 10.99 × 107 ± 1.69 × 107 a | 9.87 × 107 ± 0.56 × 107 a |
| Organic acids | ||||
| Malic acid | 7.71 × 105 ± 1.71 × 105 c | 13.95 × 105 ± 2.93 x105 a | 10.25 ×105 ± 0.79 × 105 b | 11.28 × 105 ± 0.85 × 105 b |
| Pyruvic acid | 1.06 × 103 ± 0.47 × 103 c | 1.30 × 103 ± 0.82 x103 b | 1.01 ×103 ± 0.66 × 103 c | 1.78 × 103 ± 0.41 × 103 a |
| Lactic acid | 4.83 × 104 ± 1.41 × 104 c | 7.99 × 104.4 ± 3.1 x104 b | 7.03 × 104 ± 1.34 × 104 b | 12.28 × 104 ± 1.05 × 104 a |
| Citric acid | 14.22 × 104 ± 4.85 × 105 c | 22.34 × 105 ± 6.45 x105 b | 15.41 × 105 ± 2.52 × 105 c | 36.27 × 105 ± 7.59 × 105 a |
| Succinic acid | 2.78 × 104 ± 0.83 × 104 c | 4.04 × 104 ± 1.28 x104 b | 2.69 × 104 ± 0.23 × 104 c | 5.62 × 104 ± 0.35 × 104 a |
| Fumaric acid | 4.20 × 104 ± 1.33 × 104 c | 4.65 × 104 ± 2.19 x104 c | 10.03 × 104 ± 2.38 ×104 b | 20.97 × 104 ± 1.32 × 104 a |
| Vitamins | ||||
| Nicotinic acid | 1.98 × 104 ± 0.56 × 104 b | 1.21 × 104 ± 0.46 x104 c | 9.25 × 104 ± 1.51 × 104 a | 8.60 × 104 ± 0.65 × 104 a |
| Niacin amide | 6.84 × 104 ± 1.49 × 104 c | 9.54 × 104 ± 1.86 x104 b | 9.49 × 104 ± 0.84 × 104 b | 15.00 × 104 ± 1.65 × 104 a |
| Pantothenic acid | 2.19 × 104 ± 0.41 × 104 c | 2.91 × 104 ± 0.69 x104 b | 6.92 × 104 ± 1.21 × 104 a | 6.74 × 104 ± 0.57 × 104 a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohale, K.C.; Bodede, O.; Araya, H.T.; Mudau, F.N. Metabolomic Analysis for Compositional Differences of Bush Tea (Athrixia phylicoides DC.) Subjected to Seasonal Dynamics. Agronomy 2020, 10, 892. https://doi.org/10.3390/agronomy10060892
Mohale KC, Bodede O, Araya HT, Mudau FN. Metabolomic Analysis for Compositional Differences of Bush Tea (Athrixia phylicoides DC.) Subjected to Seasonal Dynamics. Agronomy. 2020; 10(6):892. https://doi.org/10.3390/agronomy10060892
Chicago/Turabian StyleMohale, Keletso C., Olusola Bodede, Hintsa T. Araya, and Fhatuwani N. Mudau. 2020. "Metabolomic Analysis for Compositional Differences of Bush Tea (Athrixia phylicoides DC.) Subjected to Seasonal Dynamics" Agronomy 10, no. 6: 892. https://doi.org/10.3390/agronomy10060892