Incorporating Biodiversity into Biogeochemistry Models to Improve Prediction of Ecosystem Services in Temperate Grasslands: Review and Roadmap

 ,
,  , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Grasslands as Major Providers of Ecosystem Services
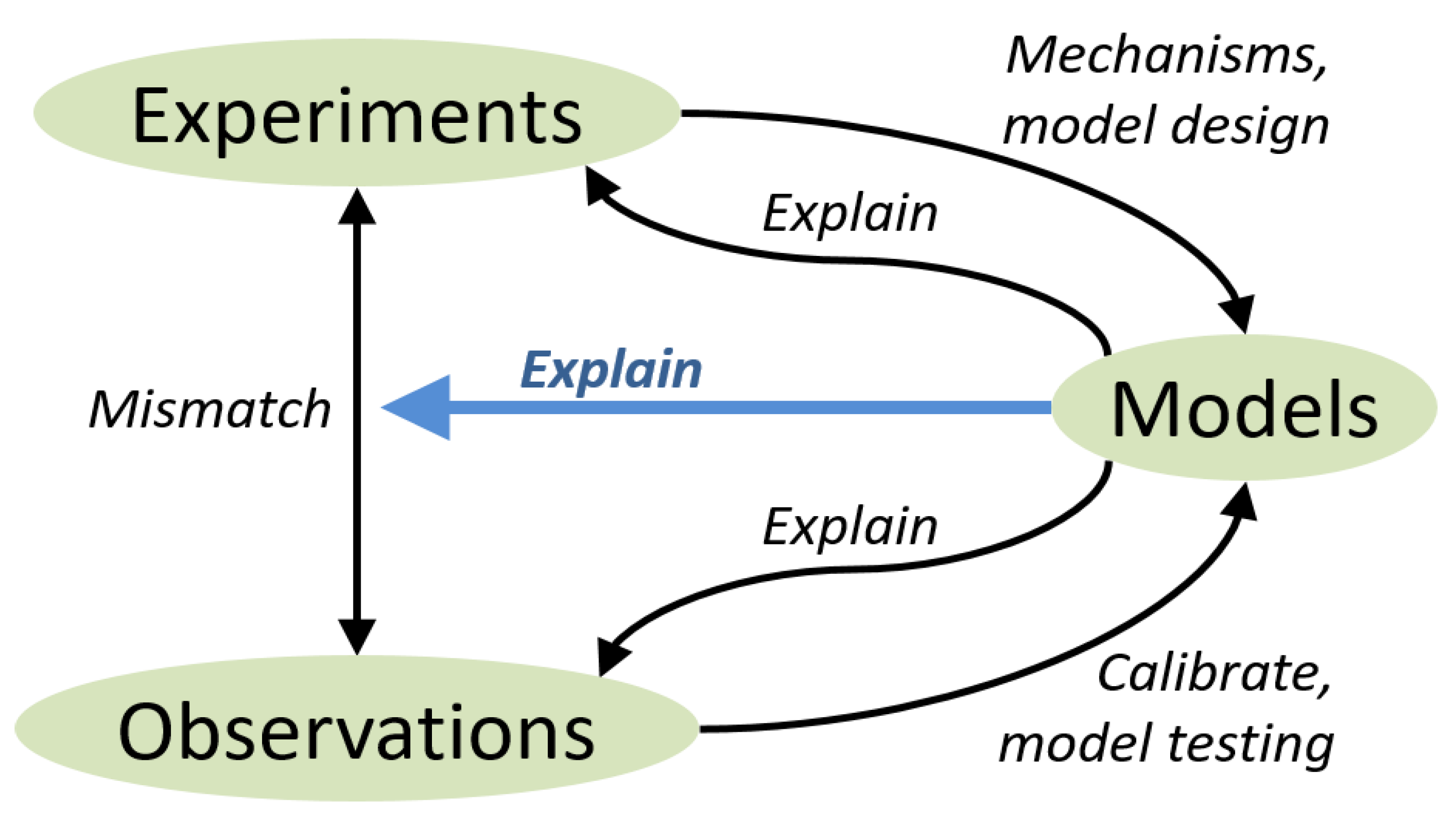
1.2. Using Models to Explain and Predict Grassland ES
1.3. Goals and Outline of This Paper
- We review the current understanding of the impact of grassland biodiversity on ES, and the drivers of biodiversity itself (Section 2).
- We review the state of the art of models for grassland biogeochemistry, and of models for the dynamics of biodiversity (Section 3). We identify the main differences between the two modelling approaches.
- We discuss how BGMs can be modified to simulate both the dynamics of biodiversity itself and the impacts of biodiversity on ES (Section 4).
- We discuss our findings and propose a roadmap for further model development and data use (Section 5).
2. Why Consider Biodiversity in Grassland Models?
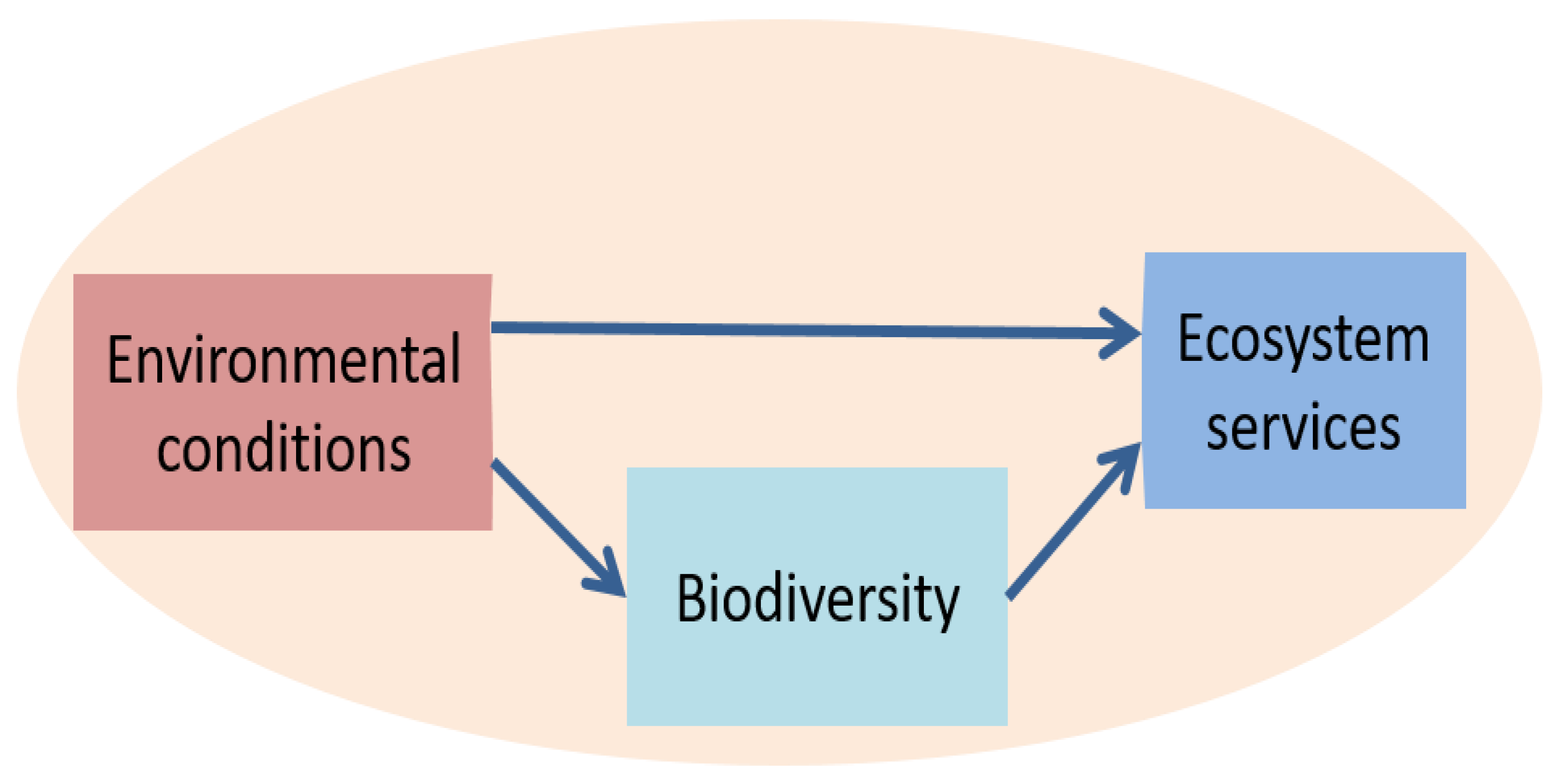
2.1. Impacts of Biodiversity on ES
- Biodiversity (observed species richness) affects the extent to which drought reduces growth and soil respiration [38]. More generally, biodiversity (species number in manipulated grassland communities) increases resistance to various types of climatic event: dry or wet, moderate or extreme, short- or long-term [39]. In marginal grasslands (e.g., on dry soils in the Mediterranean), high biodiversity may extend the growing season. This shortens the period in which soils are bare and therefore protects against soil loss by erosion. Despite the fact that high biodiversity tends to stabilize productivity by reducing climate sensitivity [39], there are examples where biodiversity reduces the stability of ecosystem functioning in response to extreme events [40].
- High biodiversity tends to lead to higher aboveground biomass, especially when species are from different functional types [41]. In grazed grasslands, high biodiversity not only improves productivity, but also grass quality and milk production [42], although the improvement may be absent under already highly productive conditions [40].
- High biodiversity in intensively managed grasslands may suppress weeds [43], but a modest increase in conventional grassland plant diversity with legumes and forbs improves pollination and therefore productivity [44]. In unfertilized grasslands, the presence of legumes increases productivity and soil organic matter, and improves soil texture [45]. The soil improvement also increases resilience against drought.
- Permanent monocultures will eventually decline in productivity due to insect herbivory or pathogen load [46].
- High species richness reduces root decomposition by increasing root C:N ratios, except for legumes [47], with grassland species having widely varying turnover rates [48]. However, high biodiversity (in mixtures that were compared to monocultures) does tend to stimulate soil microbial biomass and soil respiration [49], and it strongly increases carbon sequestration [50,51]. Overall, N-cycling processes are stimulated less by biodiversity than C-cycling [50], except when legumes are added to the mixture [52].
2.2. Impacts of Environmental Change on Biodiversity
- In mixtures subjected to an imposed extreme drought, grass species common to wetter soils (high value of the Ellenberg F index) suffered most senescence and mortality [53]. Legume species suffered more than grasses, irrespective of their Ellenberg value. Recovery was also better in grasses than in the N-fixers.
- Experimental warming tends to decrease species richness, but its impact depends strongly on the specific ecosystem [54].
- Biodiversity reduction in a Californian grassland was due to decreasing winter rain, and not due to changes in grazing, fire, N-deposition, or invasive species [55]. Losses especially concerned native annual forb species with traits indicative of low drought tolerance. In contrast, 13-year long manipulation of temperature and rainfall in infertile grasslands had only a minor effect on biodiversity (species richness and relative abundance of growth forms) and productivity with the exception of reduction from chronic summer drought [56].
- A review of herbaceous systems worldwide showed that nitrogen fertilization tends to reduce plant species richness, whereas irrigation mostly has little effect [59]. Fertilization reduces plant species richness of mountain grasslands the most where summers are cool, where mowing is carried out, and overall where biomass has been increased most by the fertilization [60]. The effects of phosphorus are generally less evident than those of nitrogen [61].
- Grazing tends to be more beneficial for biodiversity than mowing [62]. Johansen et al. [63] observed that abandonment of grazing led to decline in species diversity in Norwegian semi-natural grasslands but the effects varied with climate and soil conditions. However, grazing affects biodiversity nonlinearly: minor grazing increases biodiversity, overgrazing reduces it [64,65,66], while also the timing and the grazing species are important [67].
- In short-rotation (3–4 years) grasslands, the diversity of plant species is initiated by farmers deciding what combination of species is sown and how growth of weeds is suppressed, but weed proportions generally do increase, thereby decreasing sward productivity when they replace high yielding grass, forb and legume varieties [68].
2.3. What Can We Learn from Experiments and Field Observations?
- Grace and colleagues [76] showed that the influence of biodiversity on productivity was very small in mature natural grasslands.
- Veen et al. [77] and Weisser et al. [50] showed that results from weeded experiments may differ from non-weeded experiments. See also Kardol et al. [78] who concluded that their species-removal experiment (non-weeded) allowed for better low-diversity performance than weeded species-addition experiments.
- Gruner et al. [54] found that the experimental set-up affected the response of biodiversity to warming.
3. Modelling the Biogeochemistry and Biodiversity of Grasslands
3.1. Biogeochemistry Models (BGMs)
3.2. Models for Biodiversity
3.3. The Main Differences between BGMs and Models for Biodiversity-ES Relations
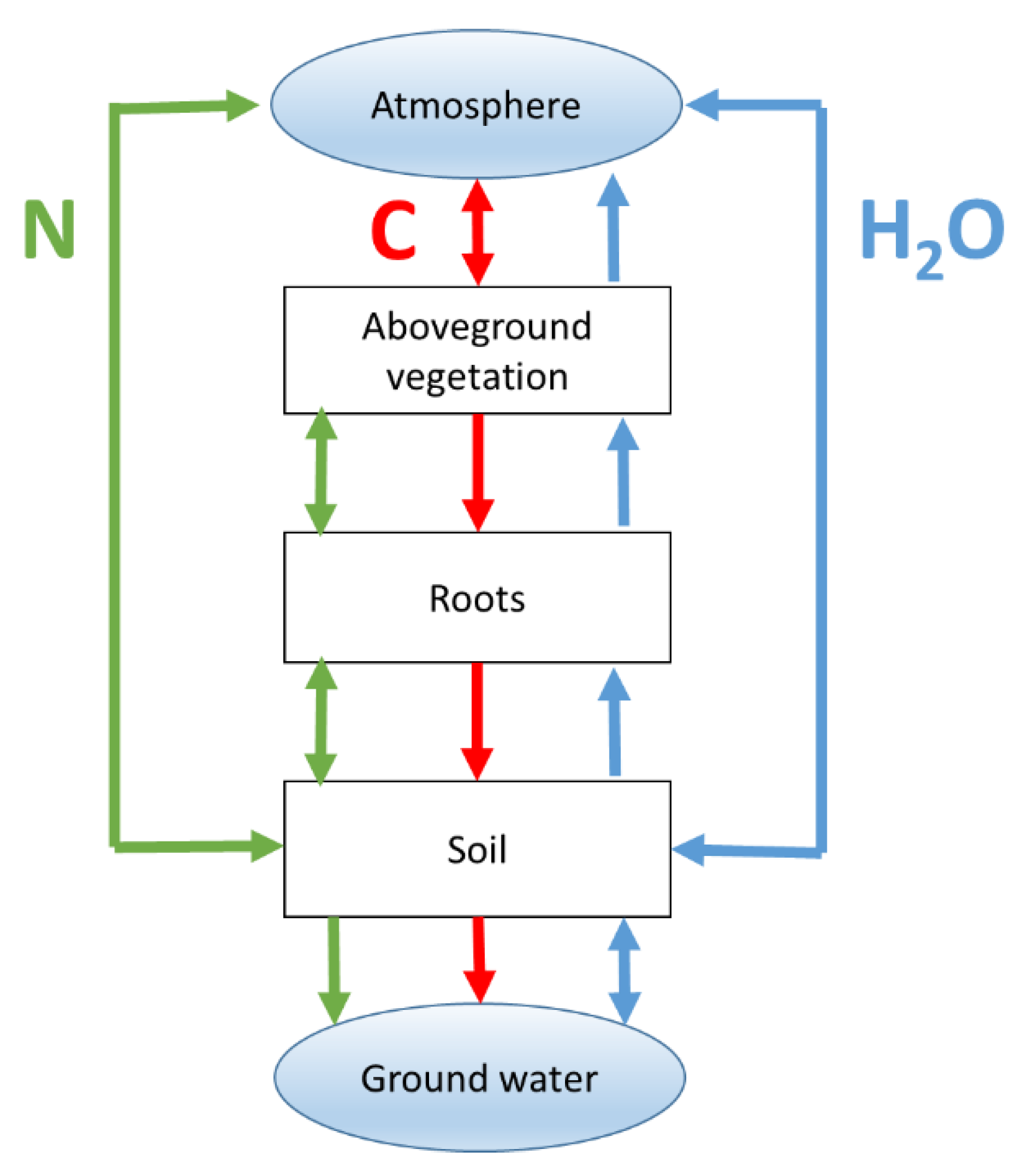
- Model structure. The simulated state variables in BGMs tend to be pools of carbon, nitrogen and water in soils, vegetation and atmosphere, whereas ecological models tend to focus on population dynamical properties such as the abundance of PFTs, species, traits or individuals (and their size-age distribution). The processes that are modelled are biogeochemical/physiological and demographic, respectively. BGMs focus on abiotic environmental drivers whereas biodiversity models usually focus on biotic interactions. For BGMs, traits are static inputs (i.e., model parameters) while for biodiversity models they are dynamic state variables.
- Mathematical formulation. BMGs are never fully analytical models: trajectories over time of state variables must be numerically derived by computer modelling. Biodiversity models can be analytically solvable systems of differential equations, but even when they are computer models, their time step of calculation tends to be much longer than the daily or sub-daily time step of most BGMs. Moreover, biodiversity computer models tend to be discrete-event agent-based (or even individual-based) models in contrast to the ‘big-leaf’ continuum BGMs.
- Data-use. BGMs require detailed information on mainly abiotic conditions (weather, atmospheric [CO2], N-deposition, soil properties) as drivers for the flows of carbon, nitrogen and water, whereas biodiversity models predominantly need biotic information, such as the initial age-size distribution of organisms or the frequency distribution of traits.
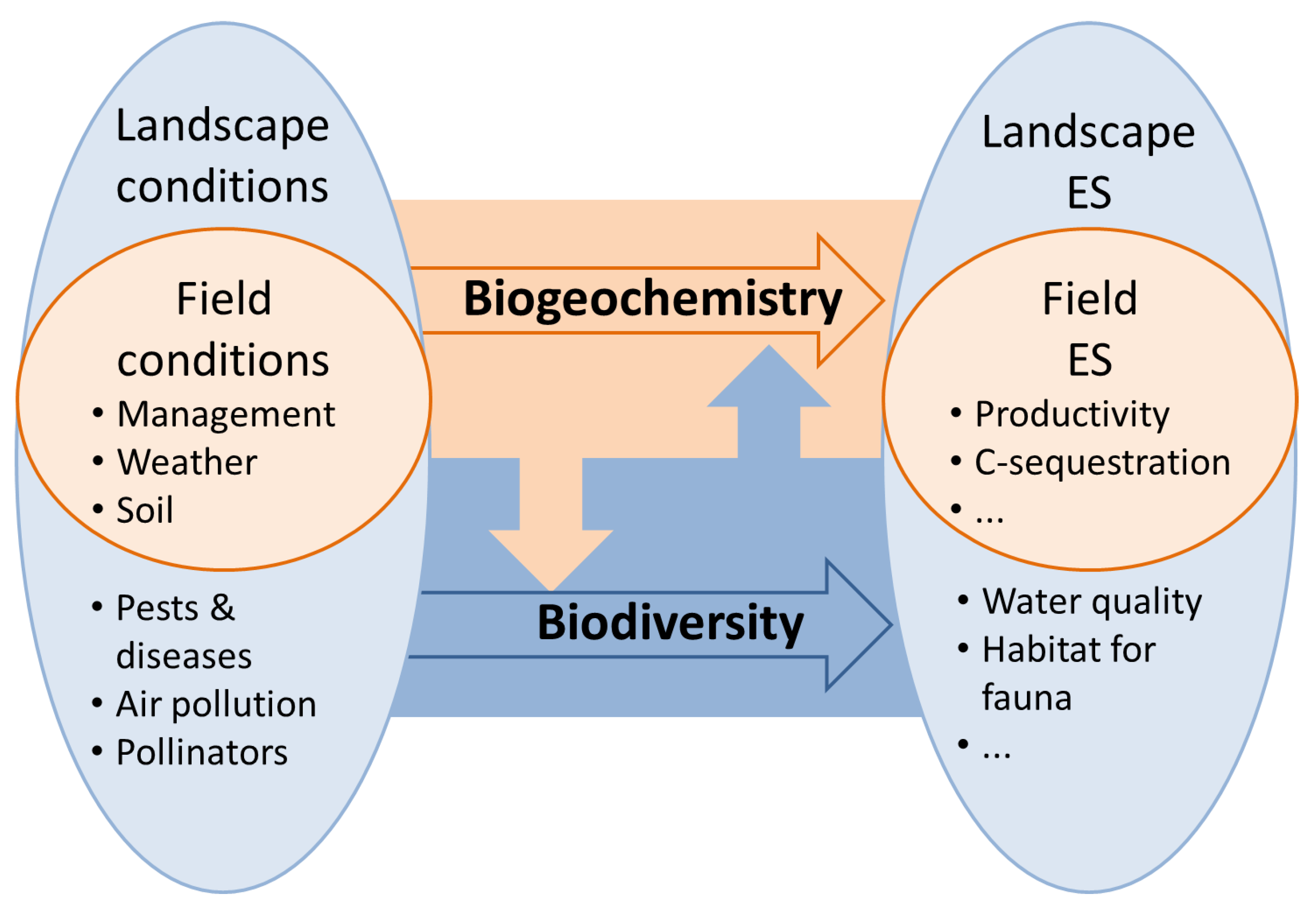
- Spatial scale. BGMs tend to be one-dimensional models that are assumed to be applicable to spatially homogeneous fields, whereas the representation of space in biodiversity models may be poorer in the vertical direction (no leaf or soil layers) but richer horizontally, even extending to simulating heterogeneous landscapes rather than fields (see also Figure 5).
4. Modifying Existing BGMs to Simulate the Impacts and Dynamics of Biodiversity
4.1. Representing Biodiversity as a Constant Metric
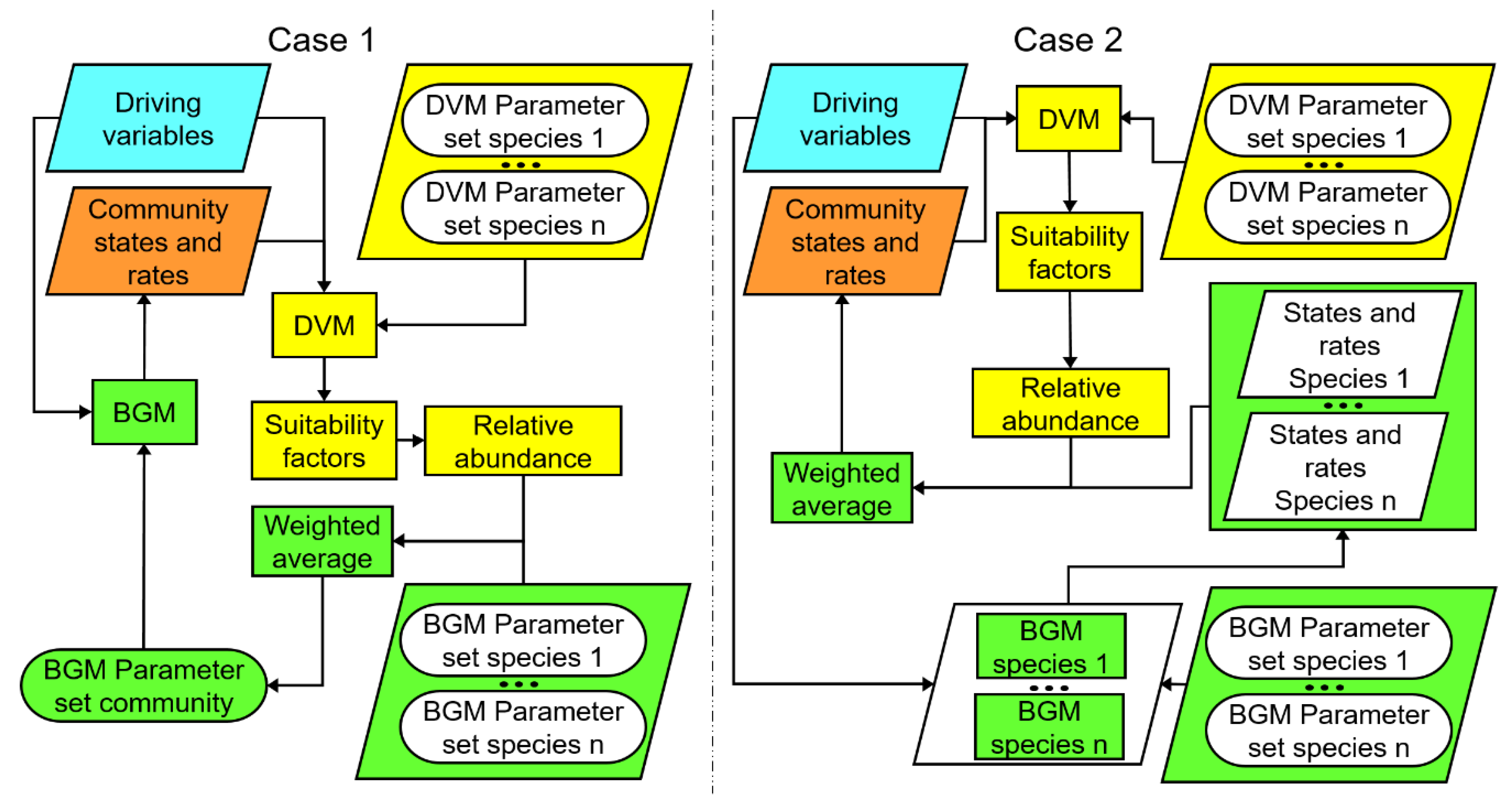
4.2. Representing Multiple Species or PFTs without Simulating Competition
4.3. Representing Multiple Competing Species or PFTs
5. Discussion
5.1. The Need and Scope for Introducing Biodiversity into BGMs
5.2. Reconciling Current BGMs and Models for Biodiversity
5.3. A Roadmap for Future Model Development
- Mechanistic modelling: nearby destinations.
- It will always be useful to have a choice of models, with different levels of complexity, and with different domains of applicability. The three different approaches to joint modelling of biogeochemistry and biodiversity that we distinguished in Section 3 are not mutually exclusive, but can be explored in parallel. Each approach comes with its own set of strengths and weaknesses, which can be assessed by frequent model comparisons against common data.
- It will be worthwhile to keep pursuing the various methods for managing model complexity that we described above (under “Reconciling Current BGMs and Models for Biodiversity”), in particular the identification of further constraints to trait-trait and trait-environment relationships. However, a trade-off between ease of implementation and parameterization on the one hand, and realistic representation of mechanisms on the other, will remain unavoidable.
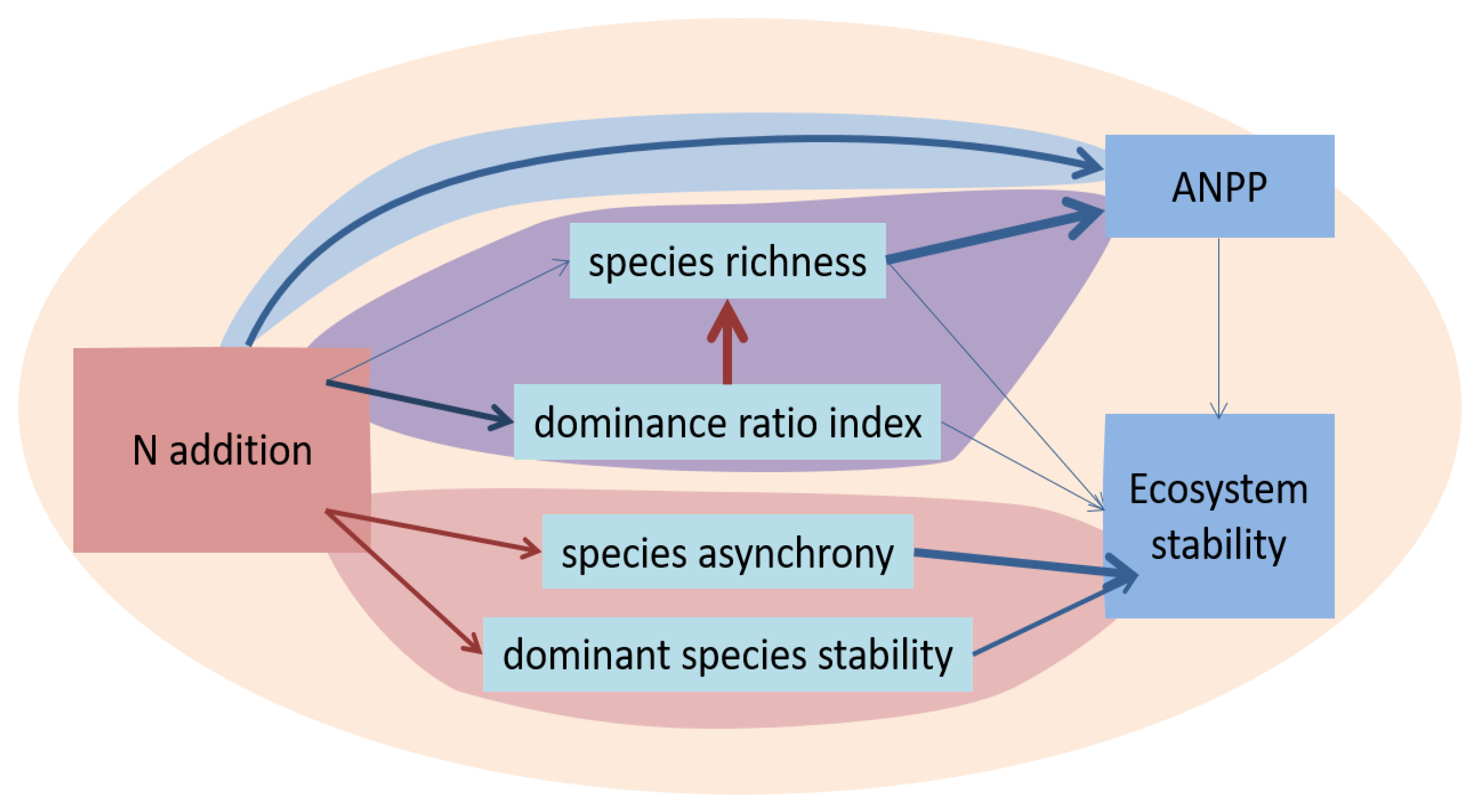
- The biogeochemical modelling will be of immediate benefit to our understanding of biodiversity-ES relationships if it explores ways to reconcile the apparent mismatch between responses of ES to manipulated biodiversity and naturally evolved biodiversity (see Figure 4).
- It will also be important to use biodiversity-representing BGMs to explore why certain responses to environmental change are seen in some grasslands but not in others, as in the examples provided by De Boeck et al. [40] where high biodiversity did not necessarily lead to greater stability of ecosystem functioning in response to extreme events.
- As BGMs are dynamic models, they can, in principle, be employed to explain how the initial response of grasslands to disturbances (resistance phase) differs from the long-term response (recovery phase), given that biodiversity can affect the two phases differently [72].
- Mechanistic modelling: long-range destinations.
- In the long term, future biogeochemistry-biodiversity modelling may aim for more ambitious applications than those documented so far. This includes more comprehensive representation of environmental drivers (e.g., phosphorus, weeds, pests and diseases) and more detail in the representation of biodiversity itself along the different dimensions of PFTs, species and traits.
- Most current BGMs have limited or no representation of spatial processes. They tend to be one-dimensional models that can be run for different locations, with site-specific environmental drivers, but true spatial processes such as species migration and hydrology are generally not represented [138]. Given the fact that the dynamics of biodiversity are only partly determined by within-field processes, future models for biodiversity and biogeochemistry may have to operate at the landscape-scale, as depicted in Figure 5. Applying process-based models to larger spatial scales requires that the interplay of species dynamics and grassland functioning is analyzed at local and regional scales, and assessed in virtual landscapes with heterogeneous soil, climate, management and natural disturbance and stress factors.
- Data collection and benchmarking.
- Model development cannot proceed without supporting data, and data analysis is hampered when reliable models are not available. The future modelling developments outlined above should thus go together with continuing increases in the quantity and diversity of data. We argued above for the parallel development of multiple modelling approaches, but this development should frequently be re-anchored in reality by comparing all models against rich benchmarking data sets, that cover multiple different ES rather than just productivity.
- Benchmarking data should be collected for the many different production situations that grasslands experience: potential growth, water-limitation, nutrient-limitation, weeds, pests, diseases, grazing, mowing. The data should cover extreme events (abiotic, biotic) as well as chronic stress conditions, in particular those that are expected to become more common in the future. Biodiversity may act to stabilize biogeochemical fluxes under extreme conditions, and BGMs need data to test their capacity to account for this.
- Remote sensing and eddy covariance measurements can be collected to assess the prevalence of biodiversity and its impact on GHG over wider areas. Eddy covariance towers have generally been placed in spatially homogeneous landscapes, to facilitate data interpretation despite variable wind directions, but this is no longer a necessity. Levy et al. [139] showed how Bayesian inference can be used to derive a spatial map of vegetation properties around a single measurement tower. A caveat is that eddy covariance measurements on grasslands tend to be unreliable during grazing events because carbon losses from animal respiration are not registered [140].
- The literature shows much evidence for an important role of biodiversity in grasslands, but uncertainties remain because of differences in environmental conditions between studies. Meta-analyses of available data should, for example, not conflate the impacts of experimentally manipulated vs. naturally evolved biodiversity. A Bayesian hierarchical approach to meta-analysis could be used to account for the interactions [141], which would prepare the data for use in model development.
- Overall, we advocate that data keep being collected in both biodiversity-manipulation experiments, and in monitoring studies where biodiversity is not imposed by the researcher (Figure 4). The first study type will provide data that elucidate causal pathways and can thus be used in the design of model structure. The second study type provides data from the actual grasslands for which our models need to be calibrated to issue reliable forecasts.
- Hybrid mechanistic-probabilistic modelling.
- We have focused here on mechanistic modelling of biogeochemistry and its relationship with biodiversity in order to predict ES under new conditions, and to explain observations of ES. However, we may want to predict a more comprehensive suite of grassland ES than is customary or even possible for biogeochemical modelling. For ES that are not outputs of BGMs, modelers could assess whether those ES are predictable from variables that the models do simulate. Aesthetic appeal, for example, may be a function of biodiversity. These added relationships are not mechanistic and may not be robust, but they can be implemented as conditional probability distributions to account for uncertainty.
- More generally, for practical application of our models and for ease of uncertainty quantification, we may want to summarize the input-output relationships of our biodiversity-enhanced BGMs in the form of graphical models (probabilistic networks), which will facilitate uncertainty and risk analysis [138].
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Gilmanov, T.G.; Soussana, J.F.; Aires, L.; Allard, V.; Ammann, C.; Balzarolo, M.; Barcza, Z.; Bernhofer, C.; Campbell, C.L.; Cernusca, A.; et al. Partitioning European grassland net ecosystem CO2 exchange into gross primary productivity and ecosystem respiration using light response function analysis. Agric. Ecosyst. Environ. 2007, 121, 93–120. [Google Scholar] [CrossRef]
- Snow, V.O.; Rotz, C.A.; Moore, A.D.; Martin-Clouaire, R.; Johnson, I.R.; Hutchings, N.J.; Eckard, R.J. The challenges and some solutions to process-based modelling of grazed agricultural systems. Environ. Model. Softw. 2014, 62, 420–436. [Google Scholar] [CrossRef]
- Fischer, M.; Bossdorf, O.; Gockel, S.; Hänsel, F.; Hemp, A.; Hessenmöller, D.; Korte, G.; Nieschulze, J.; Pfeiffer, S.; Prati, D.; et al. Implementing large-scale and long-term functional biodiversity research: The Biodiversity Exploratories. Basic Appl. Ecol. 2010, 11, 473–485. [Google Scholar] [CrossRef]
- Tribot, A.-S.; Deter, J.; Mouquet, N. Integrating the aesthetic value of landscapes and biological diversity. Proc. Biol. Sci. 2018, 285, 20180971. [Google Scholar] [CrossRef] [PubMed]
- Tilman, D.; Hill, J.; Lehman, C. Carbon-negative biofuels from low-input high-diversity grassland biomass. Science 2006, 314, 1598–1600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dass, P.; Houlton, B.Z.; Wang, Y.; Warlind, D. Grasslands may be more reliable carbon sinks than forests in California. Environ. Res. Lett. 2018, 13, 074027. [Google Scholar] [CrossRef]
- Carvalho-Santos, C.; Monteiro, A.T.; Arenas-Castro, S.; Greifeneder, F.; Marcos, B.; Portela, A.P.; Honrado, J.P. Ecosystem services in a protected mountain range of Portugal: Satellite-based products for state and trend analysis. Remote Sens. 2018, 10, 1573. [Google Scholar] [CrossRef] [Green Version]
- Levy, P.E.; Mobbs, D.C.; Jones, S.K.; Milne, R.; Campbell, C.; Sutton, M.A. Simulation of fluxes of greenhouse gases from European grasslands using the DNDC model. Agric. Ecosyst. Environ. 2007, 121, 186–192. [Google Scholar] [CrossRef]
- Chang, J.; Ciais, P.; Viovy, N.; Vuichard, N.; Sultan, B.; Soussana, J.-F. The greenhouse gas balance of European grasslands. Glob. Chang. Biol. 2015, 21, 3748–3761. [Google Scholar] [CrossRef]
- Oertel, C.; Matschullat, J.; Zurba, K.; Zimmermann, F.; Erasmi, S. Greenhouse gas emissions from soils—A review. Geochemistry 2016, 76, 327–352. [Google Scholar] [CrossRef] [Green Version]
- Bastian, O. The role of biodiversity in supporting ecosystem services in Natura 2000 sites. Ecol. Indic. 2013, 24, 12–22. [Google Scholar] [CrossRef]
- Turnbull, L.A.; Isbell, F.; Purves, D.W.; Loreau, M.; Hector, A. Understanding the value of plant diversity for ecosystem functioning through niche theory. Proc. R. Soc. B Biol. Sci. 2016, 283, 20160536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plantureux, S.; Peeters, A.; McCracken, D. Biodiversity in intensive grasslands: Effect of management, improvement and challenges. Agron. Res. 2005, 3, 153–164. [Google Scholar]
- van Klink, R.; van der Plas, F.; van Noordwijk, C.G.E.T.; WallisDeVries, M.F.; Olff, H. Effects of large herbivores on grassland arthropod diversity. Biol. Rev. Camb. Philos. Soc. 2015, 90, 347–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lange, M.; Eisenhauer, N.; Sierra, C.A.; Bessler, H.; Engels, C.; Griffiths, R.I.; Mellado-Vázquez, P.G.; Malik, A.A.; Roy, J.; Scheu, S.; et al. Plant diversity increases soil microbial activity and soil carbon storage. Nat. Commun. 2015, 6, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Yachi, S.; Loreau, M. Biodiversity and ecosystem productivity in a fluctuating environment: The insurance hypothesis. Proc. Natl. Acad. Sci. USA 1999, 96, 1463–1468. [Google Scholar] [CrossRef] [Green Version]
- Norberg, J.; Swaney, D.P.; Dushoff, J.; Lin, J.; Casagrandi, R.; Levin, S.A. Phenotypic diversity and ecosystem functioning in changing environments: A theoretical framework. Proc. Natl. Acad. Sci. USA 2001, 98, 11376–11381. [Google Scholar] [CrossRef] [Green Version]
- Thébault, A.; Mariotte, P.; Lortie, C.J.; MacDougall, A.S. Land management trumps the effects of climate change and elevated CO2 on grassland functioning. J. Ecol. 2014, 102, 896–904. [Google Scholar] [CrossRef]
- Vogel, A.; Scherer-Lorenzen, M.; Weigelt, A. Grassland resistance and resilience after drought depends on management intensity and species richness. PLoS ONE 2012, 7, e36992. [Google Scholar] [CrossRef]
- Craven, D.; Isbell, F.; Manning, P.; Connolly, J.; Bruelheide, H.; Ebeling, A.; Roscher, C.; van Ruijven, J.; Weigelt, A.; Wilsey, B.; et al. Plant diversity effects on grassland productivity are robust to both nutrient enrichment and drought. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150277. [Google Scholar] [CrossRef]
- Cleland, E.E. Biodiversity and ecosystem stability. Nat. Educ. Knowl. 2011, 3, 14. [Google Scholar]
- Craine, J.M.; Ocheltree, T.W.; Nippert, J.B.; Towne, E.G.; Skibbe, A.M.; Kembel, S.W.; Fargione, J.E. Global diversity of drought tolerance and grassland climate-change resilience. Nat. Clim. Chang. 2013, 3, 63–67. [Google Scholar] [CrossRef]
- Volaire, F.; Barkaoui, K.; Norton, M. Designing resilient and sustainable grasslands for a drier future: Adaptive strategies, functional traits and biotic interactions. Eur. J. Agron. 2014, 52, 81–89. [Google Scholar] [CrossRef] [Green Version]
- Cowles, J.M.; Wragg, P.D.; Wright, A.J.; Powers, J.S.; Tilman, D. Shifting grassland plant community structure drives positive interactive effects of warming and diversity on aboveground net primary productivity. Glob. Chang. Biol. 2016, 22, 741–749. [Google Scholar] [CrossRef] [PubMed]
- Wright, A.J.; de Kroon, H.; Visser, E.J.W.; Buchmann, T.; Ebeling, A.; Eisenhauer, N.; Fischer, C.; Hildebrandt, A.; Ravenek, J.; Roscher, C.; et al. Plants are less negatively affected by flooding when growing in species-rich plant communities. New Phytol. 2017, 213, 645–656. [Google Scholar] [CrossRef] [Green Version]
- Zeller, U.; Starik, N.; Göttert, T. Biodiversity, land use and ecosystem services: An organismic and comparative approach to different geographical regions. Glob. Ecol. Conserv. 2017, 10, 114–125. [Google Scholar] [CrossRef]
- Loreau, M.; Mazancourt, C. de Biodiversity and ecosystem stability: A synthesis of underlying mechanisms. Ecol. Lett. 2013, 16, 106–115. [Google Scholar] [CrossRef]
- Spangenberg, J.H.; Görg, C.; Truong, D.T.; Tekken, V.; Bustamante, J.V.; Settele, J. Provision of ecosystem services is determined by human agency, not ecosystem functions. Four case studies. Int. J. Biodivers. Sci. Ecosyst. Serv. Manag. 2014, 10, 40–53. [Google Scholar] [CrossRef] [Green Version]
- da Silveira Pontes, L.; Maire, V.; Schellberg, J.; Louault, F. Grass strategies and grassland community responses to environmental drivers: A review. Agron. Sustain. Dev. 2015, 35, 1297–1318. [Google Scholar] [CrossRef]
- Gaujour, E.; Amiaud, B.; Mignolet, C.; Plantureux, S. Factors and processes affecting plant biodiversity in permanent grasslands. A review. Agron. Sustain. Dev. 2012, 32, 133–160. [Google Scholar] [CrossRef] [Green Version]
- Thuiller, W.; Albert, C.; Araújo, M.B.; Berry, P.M.; Cabeza, M.; Guisan, A.; Hickler, T.; Midgley, G.F.; Paterson, J.; Schurr, F.M.; et al. Predicting global change impacts on plant species’ distributions: Future challenges. Perspect. Plant Ecol. Evol. Syst. 2008, 9, 137–152. [Google Scholar] [CrossRef]
- Balent, G.; Alard, D.; Blanfort, V.; Poudevigne, I. Pratiques de gestion, biodiversité floristique et durabilité des prairies. Fourrages 1999, 160, 385–402. [Google Scholar]
- Van Oijen, M.; Bellocchi, G.; Höglind, M. Effects of climate change on grassland biodiversity and productivity: The need for a diversity of models. Agronomy 2018, 8, 14. [Google Scholar] [CrossRef] [Green Version]
- Porqueddu, C.; Ates, S.; Louhaichi, M.; Kyriazopoulos, A.P.; Moreno, G.; del Pozo, A.; Ovalle, C.; Ewing, M.A.; Nichols, P.G.H. Grasslands in “Old World” and “New World” Mediterranean-climate zones: Past trends, current status and future research priorities. Grass Forage Sci. 2016, 71, 1–35. [Google Scholar] [CrossRef]
- Rapacz, M.; Ergon, Å.; Höglind, M.; Jørgensen, M.; Jurczyk, B.; Østrem, L.; Rognli, O.A.; Tronsmo, A.M. Overwintering of herbaceous plants in a changing climate. Still more questions than answers. Plant Sci. 2014, 225, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Rhazi, L.; Grillas, P.; Saber, E.-R.; Rhazi, M.; Brendonck, L.; Waterkeyn, A. Vegetation of Mediterranean temporary pools: A fading jewel? Hydrobiologia 2012, 689, 23–36. [Google Scholar] [CrossRef]
- Ma, L.; Derner, J.D.; Harmel, R.D.; Tatarko, J.; Moore, A.D.; Rotz, C.A.; Augustine, D.J.; Boone, R.B.; Coughenour, M.B.; Beukes, P.C.; et al. Application of grazing land models in ecosystem management: Current status and next frontiers. In Advances in Agronomy; Elsevier: New York, NY, USA, 2019; p. S0065211319300719. [Google Scholar]
- Burri, S.; Niklaus, P.A.; Grassow, K.; Buchmann, N.; Kahmen, A. Effects of plant productivity and species richness on the drought response of soil respiration in temperate grasslands. PLoS ONE 2018, 13, e0209031. [Google Scholar] [CrossRef]
- Isbell, F.; Craven, D.; Connolly, J.; Loreau, M.; Schmid, B.; Beierkuhnlein, C.; Bezemer, T.M.; Bonin, C.; Bruelheide, H.; de Luca, E.; et al. Biodiversity increases the resistance of ecosystem productivity to climate extremes. Nature 2015, 526, 574–577. [Google Scholar] [CrossRef]
- De Boeck, H.J.; Bloor, J.M.G.; Kreyling, J.; Ransijn, J.C.G.; Nijs, I.; Jentsch, A.; Zeiter, M. Patterns and drivers of biodiversity-stability relationships under climate extremes. J. Ecol. 2018, 106, 890–902. [Google Scholar] [CrossRef]
- Hector, A.; Schmid, B.; Beierkuhnlein, C.; Caldeira, M.C.; Diemer, M.; Dimitrakopoulos, P.G.; Finn, J.A.; Freitas, H.; Giller, P.S.; Good, J.; et al. Plant diversity and productivity experiments in European grasslands. Science 1999, 286, 1123–1127. [Google Scholar] [CrossRef] [Green Version]
- Roca-Fernández, A.I.; Peyraud, J.L.; Delaby, L.; Delagarde, R. Pasture intake and milk production of dairy cows rotationally grazing on multi-species swards. Animal 2016, 10, 1448–1456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Connolly, J.; Sebastià, M.-T.; Kirwan, L.; Finn, J.A.; Llurba, R.; Suter, M.; Collins, R.P.; Porqueddu, C.; Helgadóttir, Á.; Baadshaug, O.H.; et al. Weed suppression greatly increased by plant diversity in intensively managed grasslands: A continental-scale experiment. J. Appl. Ecol. 2018, 55, 852–862. [Google Scholar] [CrossRef] [PubMed]
- Orford, K.A.; Murray, P.J.; Vaughan, I.P.; Memmott, J. Modest enhancements to conventional grassland diversity improve the provision of pollination services. J. Appl. Ecol. 2016, 53, 906–915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lüscher, A.; Mueller-Harvey, I.; Soussana, J.F.; Rees, R.M.; Peyraud, J.L. Potential of legume-based grasslandLivestock systems in Europe: A review. Grass Forage Sci. 2014, 69, 206–228. [Google Scholar] [CrossRef] [PubMed]
- Crews, T.E.; Rumsey, B.E. What agriculture can learn from native ecosystems in building soil organic matter: A review. Sustainability 2017, 9, 578. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Mommer, L.; van Ruijven, J.; de Kroon, H.; Fischer, C.; Gessler, A.; Hildebrandt, A.; Scherer-Lorenzen, M.; Wirth, C.; Weigelt, A. Plant species richness negatively affects root decomposition in grasslands. J. Ecol. 2017, 105, 209–218. [Google Scholar] [CrossRef] [Green Version]
- Kagiya, N.; Reinsch, T.; Taube, F.; Salminen, J.-P.; Kluß, C.; Hasler, M.; Malisch, C.S. Turnover rates of roots vary considerably across temperate forage species. Soil Biol. Biochem. 2019, 139, 107614. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.Y.H.; Chen, X.; Huang, Z. Meta-analysis shows positive effects of plant diversity on microbial biomass and respiration. Nat. Commun. 2019, 10, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Weisser, W.W.; Roscher, C.; Meyer, S.T.; Ebeling, A.; Luo, G.; Allan, E.; Beßler, H.; Barnard, R.L.; Buchmann, N.; Buscot, F.; et al. Biodiversity effects on ecosystem functioning in a 15-year grassland experiment: Patterns, mechanisms, and open questions. Basic Appl. Ecol. 2017, 23, 1–73. [Google Scholar] [CrossRef]
- Yang, Y.; Tilman, D.; Furey, G.; Lehman, C. Soil carbon sequestration accelerated by restoration of grassland biodiversity. Nat. Commun. 2019, 10, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Wei, X.; Reich, P.B.; Hobbie, S.E. Legumes regulate grassland soil N cycling and its response to variation in species diversity and N supply but not CO2. Glob. Chang. Biol. 2019, 25, 2396–2409. [Google Scholar] [PubMed]
- Elst, E.M.; De Boeck, H.J.; Vanmaele, L.; Verlinden, M.; Dhliwayo, P.; Nijs, I. Impact of climate extremes modulated by species characteristics and richness. Perspect. Plant Ecol. Evol. Syst. 2017, 24, 80–92. [Google Scholar] [CrossRef]
- Gruner, D.S.; Bracken, M.E.S.; Berger, S.A.; Eriksson, B.K.; Gamfeldt, L.; Matthiessen, B.; Moorthi, S.; Sommer, U.; Hillebrand, H. Effects of experimental warming on biodiversity depend on ecosystem type and local species composition. Oikos 2016, 126, 8–17. [Google Scholar] [CrossRef] [Green Version]
- Harrison, S.P.; Gornish, E.S.; Copeland, S. Climate-driven diversity loss in a grassland community. Proc. Natl. Acad. Sci. USA 2015, 112, 8672–8677. [Google Scholar] [CrossRef] [Green Version]
- Grime, J.P.; Fridley, J.D.; Askew, A.P.; Thompson, K.; Hodgson, J.G.; Bennett, C.R. Long-term resistance to simulated climate change in an infertile grassland. Proc. Natl. Acad. Sci. USA 2008, 105, 10028–10032. [Google Scholar] [CrossRef] [Green Version]
- Bagella, S.; Salis, L.; Marrosu, G.M.; Rossetti, I.; Fanni, S.; Caria, M.C.; Roggero, P.P. Effects of long-term management practices on grassland plant assemblages in Mediterranean cork oak silvo-pastoral systems. Plant Ecol. 2013, 214, 621–631. [Google Scholar] [CrossRef]
- Seddaiu, G.; Bagella, S.; Pulina, A.; Cappai, C.; Salis, L.; Rossetti, I.; Lai, R.; Roggero, P.P. Mediterranean cork oak wooded grasslands: Synergies and trade-offs between plant diversity, pasture production and soil carbon. Agroforest. Syst. 2018, 92, 893–908. [Google Scholar] [CrossRef]
- DeMalach, N.; Zaady, E.; Kadmon, R. Contrasting effects of water and nutrient additions on grassland communities: A global meta-analysis. Glob. Ecol. Biogeogr. 2017, 26, 983–992. [Google Scholar] [CrossRef]
- Humbert, J.-Y.; Dwyer, J.M.; Andrey, A.; Arlettaz, R. Impacts of nitrogen addition on plant biodiversity in mountain grasslands depend on dose, application duration and climate: A systematic review. Glob. Chang. Biol. 2016, 22, 110–120. [Google Scholar] [CrossRef]
- Soons, M.B.; Hefting, M.M.; Dorland, E.; Lamers, L.P.M.; Versteeg, C.; Bobbink, R. Nitrogen effects on plant species richness in herbaceous communities are more widespread and stronger than those of phosphorus. Biol. Conserv. 2017, 212, 390–397. [Google Scholar] [CrossRef]
- Tälle, M.; Deák, B.; Poschlod, P.; Valkó, O.; Westerberg, L.; Milberg, P. Grazing vs. Mowing: A meta-analysis of biodiversity benefits for grassland management. Agric. Ecosyst. Environ. 2016, 222, 200–212. [Google Scholar]
- Johansen, L.; Taugourdeau, S.; Hovstad, K.A.; Wehn, S. Ceased grazing management changes the ecosystem services of semi-natural grasslands. Ecosyst. People 2019, 15, 192–203. [Google Scholar] [CrossRef] [Green Version]
- Török, P.; Penksza, K.; Tóth, E.; Kelemen, A.; Sonkoly, J.; Tóthmérész, B. Vegetation type and grazing intensity jointly shape grazing effects on grassland biodiversity. Ecol. Evol. 2018, 8, 10326–10335. [Google Scholar] [CrossRef] [PubMed]
- Tilman, D.; Reich, P.B.; Isbell, F. Biodiversity impacts ecosystem productivity as much as resources, disturbance, or herbivory. Proc. Natl. Acad. Sci. USA 2012, 109, 10394–10397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Z.Y.; Jiao, F.; Li, Y.H.; Kallenbach, R.L. Anthropogenic disturbances are key to maintaining the biodiversity of grasslands. Sci. Rep. 2016, 6, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grant, S.A.; Torvell, L.; Sim, E.M.; Small, J.L.; Armstrong, R.H. Controlled grazing studies on Nardus grassland: Effects of between-tussock sward height and species of grazer on Nardus utilization and floristic composition in two fields in Scotland. J. Appl. Ecol. 1996, 33, 1053–1064. [Google Scholar] [CrossRef]
- Lunnan, T.; Todnem, J.; Jørgensen, M. Botanical composition of grassland for silage in mountain districts of Norway. In Proceedings of the Sustainable Meat and Milk Production from Grasslands. In Proceedings of the 27th General Meeting of the European Grassland Federation, Cork, Ireland, 17–21 June 2018; pp. 274–276. [Google Scholar]
- Loreau, M.; Hector, A. Partitioning selection and complementarity in biodiversity experiments. Nature 2001, 412, 72–76. [Google Scholar] [CrossRef]
- Cardinale, B.J.; Matulich, K.L.; Hooper, D.U.; Byrnes, J.E.; Duffy, E.; Gamfeldt, L.; Balvanera, P.; O’Connor, M.I.; Gonzalez, A. The functional role of producer diversity in ecosystems. Am. J. Bot. 2011, 98, 572–592. [Google Scholar] [CrossRef] [Green Version]
- da S. Pontes, L.; Maire, V.; Louault, F.; Soussana, J.-F.; Carrère, P. Impacts of species interactions on grass community productivity under contrasting management regimes. Oecologia 2012, 168, 761–771. [Google Scholar] [CrossRef]
- Ingrisch, J.; Karlowsky, S.; Anadon-Rosell, A.; Hasibeder, R.; König, A.; Augusti, A.; Gleixner, G.; Bahn, M. Land use alters the drought responses of productivity and CO2 fluxes in mountain grassland. Ecosystems 2018, 21, 689–703. [Google Scholar] [CrossRef]
- Rillig, M.C.; Ryo, M.; Lehmann, A.; Aguilar-Trigueros, C.A.; Buchert, S.; Wulf, A.; Iwasaki, A.; Roy, J.; Yang, G. The role of multiple global change factors in driving soil functions and microbial biodiversity. Science 2019, 366, 886–890. [Google Scholar] [CrossRef]
- Liu, J.; Li, X.; Ma, Q.; Zhang, X.; Chen, Y.; Isbell, F.; Wang, D. Nitrogen addition reduced ecosystem stability regardless of its impacts on plant diversity. J. Ecol. 2019, 107, 2427–2435. [Google Scholar] [CrossRef]
- Rosenzweig, M.L. Species Diversity in Space and Time; Cambridge University Press: Cambridge, UK, 1995; ISBN 978-0-521-49952-1. [Google Scholar]
- Grace, J.B.; Michael Anderson, T.; Smith, M.D.; Seabloom, E.; Andelman, S.J.; Meche, G.; Weiher, E.; Allain, L.K.; Jutila, H.; Sankaran, M.; et al. Does species diversity limit productivity in natural grassland communities? Ecol. Lett. 2007, 10, 680–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veen, G.F.; van der Putten, W.H.; Bezemer, T.M. Biodiversity-ecosystem functioning relationships in a long-term non-weeded field experiment. Ecology 2018, 99, 1836–1846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kardol, P.; Fanin, N.; Wardle, D.A. Long-term effects of species loss on community properties across contrasting ecosystems. Nature 2018, 557, 710–713. [Google Scholar] [CrossRef]
- Brilli, L.; Bechini, L.; Bindi, M.; Carozzi, M.; Cavalli, D.; Conant, R.; Dorich, C.D.; Doro, L.; Ehrhardt, F.; Farina, R.; et al. Review and analysis of strengths and weaknesses of agro-ecosystem models for simulating C and N fluxes. Sci. Total Environ. 2017, 598, 445–470. [Google Scholar] [CrossRef] [Green Version]
- Korhonen, P.; Palosuo, T.; Persson, T.; Höglind, M.; Jégo, G.; Van Oijen, M.; Gustavsson, A.-M.; Bélanger, G.; Virkajärvi, P. Modelling grass yields in northern climates a comparison of three growth models for timothy. Field Crops Res. 2018, 224, 37–47. [Google Scholar] [CrossRef]
- Persson, T.; Höglind, M.; Van Oijen, M.; Korhonen, P.; Palosuo, T.; Jégo, G.; Virkajärvi, P.; Bélanger, G.; Gustavsson, A.M. Simulation of timothy nutritive value: A comparison of three process-based models. Field Crops Res. 2019, 231, 81–92. [Google Scholar] [CrossRef]
- Strullu, L.; Beaudoin, N.; Thiébeau, P.; Julier, B.; Mary, B.; Ruget, F.; Ripoche, D.; Rakotovololona, L.; Louarn, G. Simulation using the STICS model of C&N dynamics in alfalfa from sowing to crop destruction. Eur. J. Agron. 2020, 112, 125948. [Google Scholar]
- Calanca, P. Modelling the impacts of seasonal drought on herbage growth under climate change. Adv. Anim. Biosci. 2016, 7, 231–232. [Google Scholar] [CrossRef] [Green Version]
- Jouven, M.; Carrère, P.; Baumont, R. Model predicting dynamics of biomass, structure and digestibility of herbage in managed permanent pastures. 1. Model description. Grass Forage Sci. 2006, 61, 112–124. [Google Scholar] [CrossRef]
- Fitton, N.; Bindi, M.; Brilli, L.; Cichota, R.; Dibari, C.; Fuchs, K.; Huguenin-Elie, O.; Klumpp, K.; Lieffering, M.; Lüscher, A.; et al. Modelling biological N fixation and grass-legume dynamics with process-based biogeochemical models of varying complexity. Eur. J. Agron. 2019, 106, 58–66. [Google Scholar] [CrossRef]
- Gillet, F. Modelling vegetation dynamics in heterogeneous pasture-woodland landscapes. Ecol. Model. 2008, 217, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Tilman, D. Community invasibility, recruitment limitation, and grassland biodiversity. Ecology 1997, 78, 81–92. [Google Scholar] [CrossRef]
- Tilman, D. Resources: A graphical-mechanistic approach to competition and predation. Am. Nat. 1980, 116, 362–393. [Google Scholar] [CrossRef]
- Hardin, G. The competitive exclusion principle. Science 1960, 131, 1292–1297. [Google Scholar] [CrossRef] [Green Version]
- Moulin, T.; Perasso, A.; Gillet, F. Modelling vegetation dynamics in managed grasslands: Responses to drivers depend on species richness. Ecol. Model. 2018, 374, 22–36. [Google Scholar] [CrossRef]
- Schwinning, S.; Parsons, A.J. Interactions between grasses and legumes: Understanding variability in species composition. In Legumes in Sustainable Farming Systems, British Grassland Society Occasional Symposium No. 30, Proceedings of the Joint Conference of BGS and SFS Initiative, SAC, Craibstone, Aberdeen, 2–4 September 1996; Younie, D., Ed.; British Grassland Society (BGS): Hurley, UK, 1996; pp. 153–163. [Google Scholar]
- Loreau, M.; Mouquet, N.; Gonzalez, A. Biodiversity as spatial insurance in heterogeneous landscapes. Proc. Natl. Acad. Sci. USA 2003, 100, 12765–12770. [Google Scholar] [CrossRef] [Green Version]
- de Mazancourt, C.; Isbell, F.; Larocque, A.; Berendse, F.; De Luca, E.; Grace, J.B.; Haegeman, B.; Wayne Polley, H.; Roscher, C.; Schmid, B.; et al. Predicting ecosystem stability from community composition and biodiversity. Ecol. Lett. 2013, 16, 617–625. [Google Scholar] [CrossRef] [Green Version]
- Siehoff, S.; Lennartz, G.; Heilburg, I.C.; Roß-Nickoll, M.; Ratte, H.T.; Preuss, T.G. Process-based modeling of grassland dynamics built on ecological indicator values for land use. Ecol. Model. 2011, 222, 3854–3868. [Google Scholar] [CrossRef]
- Westoby, M.; Walker, B.; Noy-Meir, I. Opportunistic management for rangelands not at equilibrium. J. Range Manag. 1989, 42, 266–274. [Google Scholar] [CrossRef]
- Taubert, F.; Frank, K.; Huth, A. A review of grassland models in the biofuel context. Ecol. Model. 2012, 245, 84–93. [Google Scholar] [CrossRef]
- Weiss, L.; Pfestorf, H.; May, F.; Körner, K.; Boch, S.; Fischer, M.; Müller, J.; Prati, D.; Socher, S.A.; Jeltsch, F. Grazing response patterns indicate isolation of semi-natural European grasslands. Oikos 2014, 123, 599–612. [Google Scholar] [CrossRef]
- May, F.; Grimm, V.; Jeltsch, F. Reversed effects of grazing on plant diversity: The role of below-ground competition and size symmetry. Oikos 2009, 118, 1830–1843. [Google Scholar] [CrossRef]
- Milchunas, D.G.; Sala, O.E.; Lauenroth, W. A generalized model of the effects of grazing by large herbivores on grassland community structure. Am. Nat. 1988, 132, 87–106. [Google Scholar] [CrossRef]
- Fox, J.W. The intermediate disturbance hypothesis should be abandoned. Trends Ecol. Evol. 2013, 28, 86–92. [Google Scholar] [CrossRef]
- Lohmann, D.; Guo, T.; Tietjen, B. Zooming in on coarse plant functional types: Simulated response of savanna vegetation composition in response to aridity and grazing. Theor. Ecol. 2018, 11, 161–173. [Google Scholar] [CrossRef]
- Bittebiere, A.K.; Mony, C.; Clément, B.; Garbey, M. Modeling competition between plants using an Individual Based Model: Methods and effects on the growth of two species with contrasted growth forms. Ecol. Model. 2012, 234, 38–50. [Google Scholar] [CrossRef]
- van Bodegom, P.M.; Douma, J.C.; Verheijen, L.M. A fully traits-based approach to modeling global vegetation distribution. Proc. Natl. Acad. Sci. USA 2014, 111, 13733–13738. [Google Scholar] [CrossRef] [Green Version]
- Laughlin, D.C.; Joshi, C.; van Bodegom, P.M.; Bastow, Z.A.; Fulé, P.Z. A predictive model of community assembly that incorporates intraspecific trait variation. Ecol. Lett. 2012, 15, 1291–1299. [Google Scholar] [CrossRef]
- Laughlin, D.C.; Joshi, C.; Richardson, S.J.; Peltzer, D.A.; Mason, N.W.H.; Wardle, D.A. Quantifying multimodal trait distributions improves trait-based predictions of species abundances and functional diversity. J. Veg. Sci. 2015, 26, 46–57. [Google Scholar] [CrossRef]
- Van Oijen, M.; Höglind, M. Toward a Bayesian procedure for using process-based models in plant breeding, with application to ideotype design. Euphytica 2016, 207, 627–643. [Google Scholar] [CrossRef] [Green Version]
- Doležal, J.; Lanta, V.; Mudrák, O.; Lepš, J. Seasonality promotes grassland diversity: Interactions with mowing, fertilization and removal of dominant species. J. Ecol. 2019, 107, 203–215. [Google Scholar] [CrossRef]
- Xenakis, G.; Ray, D.; Mencuccini, M. Sensitivity and uncertainty analysis from a coupled 3-PG and soil organic matter decomposition model. Ecol. Model. 2008, 219, 1–16. [Google Scholar] [CrossRef]
- Thornley, J. Simulating grass-legume dynamics: A phenomenological submodel. Ann. Bot. 2001, 88, 905–913. [Google Scholar] [CrossRef] [Green Version]
- Confalonieri, R. CoSMo: A simple approach for reproducing plant community dynamics using a single instance of generic crop simulators. Ecol. Model. 2014, 286, 1–10. [Google Scholar] [CrossRef]
- Movedi, E.; Bellocchi, G.; Argenti, G.; Paleari, L.; Vesely, F.; Staglianò, N.; Dibari, C.; Confalonieri, R. Development of generic crop models for simulation of multi-species plant communities in mown grasslands. Ecol. Model. 2019, 401, 111–128. [Google Scholar] [CrossRef]
- Malézieux, E.; Crozat, Y.; DUPRAZ, C.; Laurans, M.; Makowski, D.; Ozier-Lafontaine, H.; Rapidel, B.; De Tourdonnet, S.; Valantin-Morison, M. Mixing plant species in cropping systems: Concepts, tools and models. A review. Agron. Sustain. Dev. 2009, 29, 43–62. [Google Scholar] [CrossRef] [Green Version]
- Sakschewski, B.; von Bloh, W.; Boit, A.; Rammig, A.; Kattge, J.; Poorter, L.; Peñuelas, J.; Thonicke, K. Leaf and stem economics spectra drive diversity of functional plant traits in a dynamic global vegetation model. Glob. Chang. Biol. 2015, 21, 2711–2725. [Google Scholar] [CrossRef]
- Louarn, G.; Faverjon, L. A generic individual-based model to simulate morphogenesis, CN acquisition and population dynamics in contrasting forage legumes. Ann. Bot. 2018, 121, 875–896. [Google Scholar] [CrossRef]
- Colbach, N.; Gardarin, A.; Moreau, D. The response of weed and crop species to shading: Which parameters explain weed impacts on crop production? Field Crops Res. 2019, 238, 45–55. [Google Scholar] [CrossRef]
- Modelling Crop-Weed Interactions; Kropff, M.; van Laar, H.H. (Eds.) CABI Publishing: Wallingford, UK, 1993; ISBN 978-0-85198-745-3. [Google Scholar]
- Van Noordwijk, M.; Lusiana, B. WaNuLCAS, a model of water, nutrient and light capture in agroforestry systems. Agrofor. Syst. 1998, 43, 217–242. [Google Scholar] [CrossRef]
- Van Oijen, M.; Dauzat, J.; Harmand, J.-M.; Lawson, G.; Vaast, P. Coffee agroforestry systems in Central America: II. Development of a simple process-based model and preliminary results. Agrofor. Syst. 2010, 80, 361–378. [Google Scholar] [CrossRef]
- Corre-Hellou, G.; Faure, M.; Launay, M.; Brisson, N.; Crozat, Y. Adaptation of the STICS intercrop model to simulate crop growth and N accumulation in pea-barley intercrops. Field Crops Res. 2009, 113, 72–81. [Google Scholar] [CrossRef]
- Lazzarotto, P.; Calanca, P.; Fuhrer, J. Dynamics of grass clover mixtures: An analysis of the response to management with the PROductive GRASsland Simulator (PROGRASS). Ecol. Model. 2009, 220, 703–724. [Google Scholar] [CrossRef]
- Sharp, J.M.; Edwards, G.R.; Jeger, M.J. A spatially explicit population model of the effect of spatial scale of heterogeneity in grass-clover grazing systems. J. Agric. Sci. 2014, 152, 394–407. [Google Scholar] [CrossRef]
- Biondini, M. A three-dimensional spatial model for plant competition in a heterogeneous soil environment. Ecol. Model. 2001, 142, 189–225. [Google Scholar] [CrossRef]
- Vos, J.; Evers, J.B.; Buck-Sorlin, G.H.; Andrieu, B.; Chelle, M.; de Visser, P.H.B. Functional-structural plant modelling: A new versatile tool in crop science. J. Exp. Bot. 2010, 61, 2101–2115. [Google Scholar] [CrossRef]
- Faverjon, L.; Escobar-Gutiérrez, A.; Litrico, I.; Julier, B.; Louarn, G. A generic individual-based model can predict yield, nitrogen content, and species abundance in experimental grassland communities. J. Exp. Bot. 2019, 70, 2491–2504. [Google Scholar] [CrossRef]
- Soussana, J.-F.; Maire, V.; Gross, N.; Bachelet, B.; Pagès, L.; Martin, R.; Hill, D.; Wirth, C. Gemini: A grassland model simulating the role of plant traits for community dynamics and ecosystem functioning. Parameterization and evaluation. Ecol. Model. 2012, 231, 134–145. [Google Scholar] [CrossRef]
- Maire, V.; Soussana, J.F.; Gross, N.; Bachelet, B.; Pagès, L.; Martin, R.; Reinhold, T.; Wirth, C.; Hill, D. Plasticity of plant form and function sustains productivity and dominance along environment and competition gradients. A modeling experiment with Gemini. Ecol. Model. 2013, 254, 80–91. [Google Scholar] [CrossRef]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.C.; Diemer, M.; et al. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef] [PubMed]
- Maire, V.; Gross, N.; Hill, D.; Martin, R.; Wirth, C.; Wright, I.J.; Soussana, J.-F. Disentangling coordination among functional traits using an individual-centred model: Impact on plant performance at intra- and inter-specific levels. PLoS ONE 2013, 8, e77372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kipling, R.P.; Virkajärvi, P.; Breitsameter, L.; Curnel, Y.; De Swaef, T.; Gustavsson, A.-M.; Hennart, S.; Höglind, M.; Järvenranta, K.; Minet, J.; et al. Key challenges and priorities for modelling European grasslands under climate change. Sci. Total Environ. 2016, 566, 851–864. [Google Scholar] [CrossRef] [Green Version]
- Kattge, J.; DíAz, S.; Lavorel, S.; Prentice, I.C.; Leadley, P.; BöNisch, G.; Garnier, E.; Westoby, M.; Reich, P.B.; Wright, I.J.; et al. TRY—A global database of plant traits. Glob. Chang. Biol. 2011, 17, 2905–2935. [Google Scholar] [CrossRef]
- Verheijen, L.M.; Aerts, R.; Brovkin, V.; Cavender-Bares, J.; Cornelissen, J.H.C.; Kattge, J.; van Bodegom, P.M. Inclusion of ecologically based trait variation in plant functional types reduces the projected land carbon sink in an earth system model. Glob. Chang. Biol. 2015, 21, 3074–3086. [Google Scholar] [CrossRef]
- Peaucelle, M.; Bacour, C.; Ciais, P.; Vuichard, N.; Kuppel, S.; Peñuelas, J.; Marchesini, L.B.; Blanken, P.D.; Buchmann, N.; Chen, J.; et al. Covariations between plant functional traits emerge from constraining parameterization of a terrestrial biosphere model. Glob. Ecol. Biogeogr. 2019, 28, 1351–1365. [Google Scholar] [CrossRef] [Green Version]
- Van Oijen, M. Bayesian methods for quantifying and reducing uncertainty and error in forest models. Curr. For. Rep. 2017, 3, 269–280. [Google Scholar] [CrossRef] [Green Version]
- Cruz, P.; Theau, J.-P.; Lecloux, E.; Jouany, C.; Duru, M. Typologie fonctionnelle de graminées fourragères pérennes: Une classification multitraits. Fourrages 2010, 201, 11–17. [Google Scholar]
- Maire, V.; Martre, P.; Kattge, J.; Gastal, F.; Esser, G.; Fontaine, S.; Soussana, J.-F. The coordination of leaf photosynthesis links C and N Fluxes in C3 plant species. PLoS ONE 2012, 7, e38345. [Google Scholar] [CrossRef]
- Prentice, I.C.; Dong, N.; Gleason, S.M.; Maire, V.; Wright, I.J. Balancing the costs of carbon gain and water transport: Testing a new theoretical framework for plant functional ecology. Ecol. Lett. 2014, 17, 82–91. [Google Scholar] [CrossRef]
- Chen, T.-W.; Cabrera-Bosquet, L.; Alvarez Prado, S.; Perez, R.; Artzet, S.; Pradal, C.; Coupel-Ledru, A.; Fournier, C.; Tardieu, F. Genetic and environmental dissection of biomass accumulation in multi-genotype maize canopies. J. Exp. Bot. 2019, 70, 2523–2534. [Google Scholar] [CrossRef] [PubMed]
- Van Oijen, M. Tools for landscape science: Theory, models and data. In Current Trends in Landscape Research; Müller, L., Eulenstein, F., Eds.; Springer: Berlin, Germany, 2019; pp. 219–231. ISBN 978-3-030-30069-2. [Google Scholar]
- Levy, P.; Drewer, J.; Jammet, M.; Leeson, S.; Friborg, T.; Skiba, U.; van Oijen, M. Inference of spatial heterogeneity in surface fluxes from eddy covariance data: A case study from a subarctic mire ecosystem. Agric. For. Meteorol. 2020, 280, 107783. [Google Scholar] [CrossRef]
- Puche, N.; Senapati, N.; Flechard, C.R.; Klumpp, K.; Kirschbaum, M.U.F.; Chabbi, A. Modeling carbon and water fluxes of managed grasslands: Comparing flux variability and net carbon budgets between grazed and mowed systems. Agronomy 2019, 9, 183. [Google Scholar] [CrossRef] [Green Version]
- Gurevitch, J.; Koricheva, J.; Nakagawa, S.; Stewart, G. Meta-analysis and the science of research synthesis. Nature 2018, 555, 175–182. [Google Scholar] [CrossRef] [PubMed]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Van Oijen, M.; Barcza, Z.; Confalonieri, R.; Korhonen, P.; Kröel-Dulay, G.; Lellei-Kovács, E.; Louarn, G.; Louault, F.; Martin, R.; Moulin, T.; et al. Incorporating Biodiversity into Biogeochemistry Models to Improve Prediction of Ecosystem Services in Temperate Grasslands: Review and Roadmap. Agronomy 2020, 10, 259. https://doi.org/10.3390/agronomy10020259
Van Oijen M, Barcza Z, Confalonieri R, Korhonen P, Kröel-Dulay G, Lellei-Kovács E, Louarn G, Louault F, Martin R, Moulin T, et al. Incorporating Biodiversity into Biogeochemistry Models to Improve Prediction of Ecosystem Services in Temperate Grasslands: Review and Roadmap. Agronomy. 2020; 10(2):259. https://doi.org/10.3390/agronomy10020259
Chicago/Turabian StyleVan Oijen, Marcel, Zoltán Barcza, Roberto Confalonieri, Panu Korhonen, György Kröel-Dulay, Eszter Lellei-Kovács, Gaëtan Louarn, Frédérique Louault, Raphaël Martin, Thibault Moulin, and et al. 2020. "Incorporating Biodiversity into Biogeochemistry Models to Improve Prediction of Ecosystem Services in Temperate Grasslands: Review and Roadmap" Agronomy 10, no. 2: 259. https://doi.org/10.3390/agronomy10020259