Tipburn Incidence and Ca Acquisition and Distribution in Lisianthus (Eustoma grandiflorum (Raf.) Shinn.) Cultivars under Different Ca Concentrations in Nutrient Solution

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plants
2.2. Experimental Design
2.3. Tipburn Severity and Incidence
2.4. Measurement of Dry Weight and Ca Concentration
2.5. Ca acquisition Competence
2.6. Statistical Analysis
3. Results and Discussion
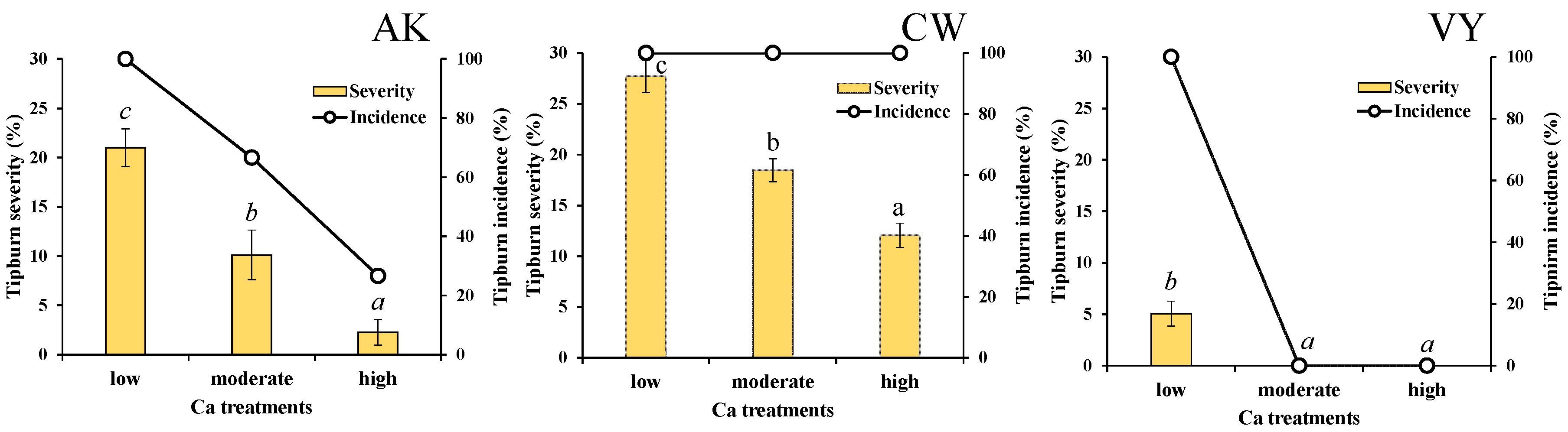
3.1. Tipburn Severity and Incidence
3.2. Plant Growth
3.3. Ca Acquisition Competence
3.4. Ca Distribution
3.4.1. Ca Concentrations of Whole Leaves, Stems, and Roots
3.4.2. Ca Concentrations of Upper (1st), Middle (4th), and Lower (7th) Leaves
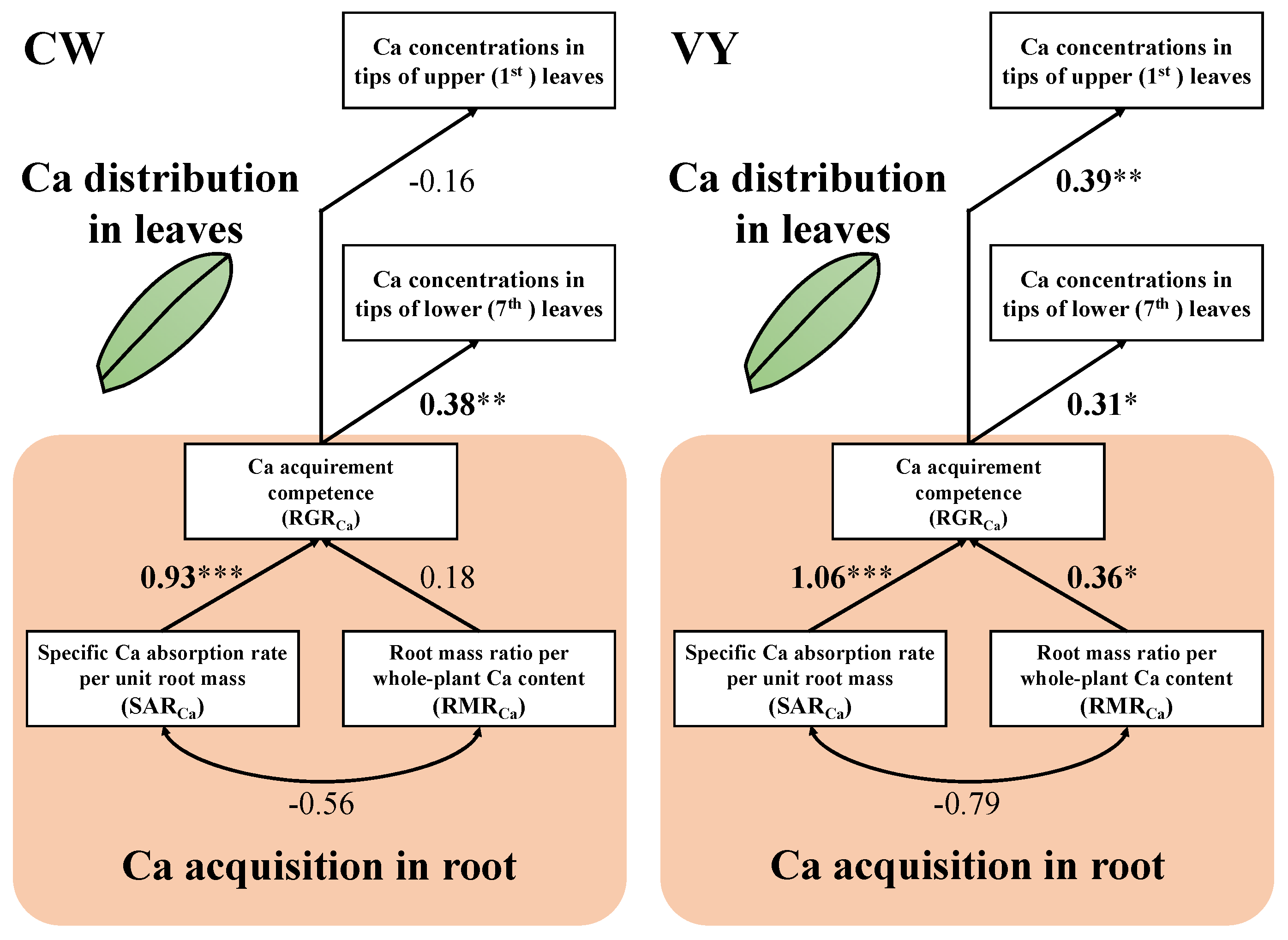
3.5. Contributions of Ca Acquisition and Ca Distribution under Different Ca Concentrations
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Barta, D.J.; Tibbitts, T.W. Calcium localization in lettuce leaves with and without tipburn: Comparison of controlled-environment and field-grown plants. J. Am. Soc. Hortic. Sci. 1991, 116, 870–875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wissemeier, A.H.; Zühlke, G. Relation between climatic variables, growth and the incidence of tipburn in field-grown lettuce as evaluated by simple, partial and multiple regression analysis. Sci. Hortic. 2002, 93, 193–204. [Google Scholar] [CrossRef]
- Sago, Y. Effects of light intensity and growth rate on tipburn development and leaf calcium concentration in butterhead lettuce. HortScience 2016, 51, 1087–1091. [Google Scholar] [CrossRef]
- Uno, Y.; Okubo, H.; Itoh, H.; Koyama, R. Reduction of leaf lettuce tipburn using an indicator cultivar. Sci. Hortic. 2016, 210, 14–18. [Google Scholar] [CrossRef]
- Holmes, S.C.; Wells, D.E.; Pickens, J.M.; Kemble, J.M. Selection of Heat Tolerant Lettuce (Lactuca sativa L.) Cultivars Grown in Deep Water Culture and Their Marketability. Horticulturae 2019, 5, 50. [Google Scholar] [CrossRef] [Green Version]
- Kuo, C.G.; Tsay, J.S.; Tsai, C.L.; Chen, R.J. Tipburn of Chinese cabbage in relation to calcium nutrition and distribution. Sci. Hortic. 1981, 14, 131–138. [Google Scholar] [CrossRef]
- Aloni, B.; Pashkar, T.; Libel, R. The possible involvement of gibberellins and calcium in tipburn of Chinese cabbage: Study of intact plants and detached leaves. Plant Growth Regul. 1986, 4, 3–11. [Google Scholar] [CrossRef]
- Pressman, E.; Shaked, R.; Arcan, L. The effect of flower-inducing factors on leaf tipburn formation in Chinese cabbage. J. Plant Physiol. 1993, 141, 210–214. [Google Scholar] [CrossRef]
- Magnusson, M. Mineral fertilizers and green mulch in Chinese cabbage [Brassica pekinensis (Lour.) Rupr.]: Effect on nutrient uptake, yield and internal tipburn. Acta Agric. Scand. Sect. B Plant Soil Sci. 2002, 52, 25–35. [Google Scholar] [CrossRef]
- Mason, G.F.; Guttridge, C.G. The influence of relative humidity and nutrition on leaf tipburn of strawberry. Sci. Hortic. 1975, 3, 339–349. [Google Scholar] [CrossRef]
- Guttridge, C.G.; Bradfield, E.G.; Holder, R. Dependence of calcium transport into strawberry leaves on positive pressure in the xylem. Ann. Bot. 1981, 48, 473–480. [Google Scholar] [CrossRef]
- Bautista, A.S.; López-Galarza, S.; Martínez, A.; Pascual, B.; Maroto, J.V. Influence of cation proportions of the nutrient solution on tipburn incidence in strawberry plants. J. Plant Nutr. 2009, 32, 1527–1539. [Google Scholar] [CrossRef]
- Palencia, P.; Martinez, F.; Ribeiro, E.; Pestana, M.; Gama, F.; Saavedra, T.; de Varennes, A.; Correia, P.J. Relationship between tipburn and leaf mineral composition in strawberry. Sci. Hortic. 2010, 126, 242–246. [Google Scholar] [CrossRef]
- Chang, Y.C.; Miller, W.B. Growth and calcium partitioning in Lilium ‘Star Gazer’ in relation to leaf calcium deficiency. J. Am. Soc. Hortic. Sci. 2003, 128, 788–796. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.C.; Miller, W.B. The relationship between leaf enclosure, transpiration, and upper leaf necrosis on Lilium ‘Star Gazer’. J. Am. Soc. Hortic. Sci. 2004, 129, 128–133. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.C.; Miller, W.B. The development of upper leaf necrosis in Lilium ‘Star Gazer’. J. Am. Soc. Hortic. Sci. 2005, 130, 759–766. [Google Scholar] [CrossRef]
- Kuronuma, T.; Watanabe, Y.; Ando, M.; Watanabe, H. Relevance of tipburn incidence to the competence for Ca acquirement and Ca distributivity in lisianthus [Eustoma grandiflorum (Raf.) Shinn.] cultivars. Sci. Hortic. 2019, 246, 805–811. [Google Scholar] [CrossRef]
- Kuronuma, T.; Kinoshita, N.; Ando, M.; Watanabe, H. Difference of Ca distribution before and after the onset of tipburn in lisianthus [Eustoma grandiflorum (Raf.) Shinn.] cultivars. Sci. Hortic. 2020, 261. [Google Scholar] [CrossRef]
- Garnier, E. Resource capture, biomass allocation and growth in herbaceous plants. Trends Ecol. Evol. 1991, 6, 126–131. [Google Scholar] [CrossRef]
- Kuronuma, T.; Watanabe, H. Physiological and morphological traits and competence for carbon sequestration of several green roof plants under a controlled environmental system. J. Am. Soc. Hortic. Sci. 2016, 141, 583–590. [Google Scholar] [CrossRef] [Green Version]
- Kuronuma, T.; Watanabe, H. Relevance of carbon sequestration to the physiological and morphological traits of several green roof plants during the first year after construction. Am. J. Plant Sci. 2016, 8, 14–27. [Google Scholar] [CrossRef] [Green Version]
- Poorter, H.; Remkes, C. Leaf area ratio and net assimilation rate of 24 wild species differing in relative growth rate. Oecologia 1990, 83, 553–559. [Google Scholar] [CrossRef] [PubMed]
- Osone, Y.; Ishida, A.; Tateno, M. Correlation between relative growth rate and specific leaf area requires associations of specific leaf area with nitrogen absorption rate of roots. New Phytol. 2008, 179, 417–427. [Google Scholar] [CrossRef] [PubMed]
- Imada, S.; Yamanaka, N.; Tami, S. Contribution of root growth responses to leaf traits and relative growth rate of Populus alba under different water-table conditions. Trees 2010, 24, 1163–1172. [Google Scholar] [CrossRef]
- Oguchi, R.; Ozaki, H.; Hanada, K.; Hikosaka, K. Which plant trait explains the variations in relative growth rate and its response to elevated carbon dioxide concentration among Arabidopsis thaliana ecotypes derived from a variety of habitats? Oecologia 2016, 180, 865–876. [Google Scholar] [CrossRef]
- Kuronuma, T.; Watanabe, Y.; Ando, M.; Watanabe, H. Tipburn severity and calcium distribution in lisianthus (Eustoma Grandiflorum (Raf.) Shinn.) cultivars under different relative air humidity conditions. Agronomy 2018, 8, 218. [Google Scholar] [CrossRef] [Green Version]
- Hunt, R. Plant Growth Curves: The Functional Approach to Plant Growth Analysis; Edward Arnold Ltd.: London, UK, 1982. [Google Scholar]
- Lee, J.; Park, I.; Lee, Z.W.; Kim, S.W.; Baek, N.; Park, H.S.; Park, S.U.; Kwon, S.Y.; Kim, H. Regulation of the major vacuolar Ca2+ transporter genes, by intercellular Ca2+ concentration and abiotic stresses, in tip-burn resistant Brassica oleracea. Mol. Boil. Rep. 2013, 40, 177–188. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Cultivars | Treatments | Dry Weight (g) | LMR (g/g) | SMR (g/g) | RMR (g/g) | ||||
|---|---|---|---|---|---|---|---|---|---|
| AK | low | 3.7 | a | 0.56 | n.s. | 0.20 | c | 0.24 | a |
| moderate | 5.4 | b | 0.55 | 0.18 | b | 0.27 | ab | ||
| high | 5.0 | b | 0.55 | 0.15 | a | 0.30 | b | ||
| CW | low | 4.2 | a | 0.63 | n.s. | 0.18 | b | 0.19 | a |
| moderate | 5.2 | ab | 0.61 | 0.16 | a | 0.23 | ab | ||
| high | 5.9 | b | 0.59 | 0.16 | a | 0.24 | b | ||
| VY | low | 4.6 | a | 0.57 | n.s. | 0.14 | b | 0.29 | n.s. |
| moderate | 5.5 | b | 0.57 | 0.11 | a | 0.31 | |||
| high | 5.4 | b | 0.57 | 0.12 | ab | 0.31 | |||
| Cultivars | Treatments | Total Ca Concentrations | RGRCa | SARCa | RMRCa | ||||
|---|---|---|---|---|---|---|---|---|---|
| (mg-Ca/kg-DW) | (mg-Ca mg-Ca−1 week−1) | (mg-Ca g-DW−1 week−1) | (mg-Ca/g-DW) | ||||||
| AK | low | 2.7 | a | 0.59 | a | 6.1 | a | 0.11 | n.s. |
| moderate | 3.9 | b | 0.68 | b | 8.8 | b | 0.10 | ||
| high | 4.4 | b | 0.69 | b | 9.1 | b | 0.10 | ||
| CW | low | 2.7 | a | 0.60 | a | 7.4 | a | 0.10 | b |
| moderate | 3.8 | b | 0.67 | b | 9.7 | b | 0.09 | a | |
| high | 4.5 | c’ | 0.71 | b | 11.2 | c | 0.09 | a | |
| VY | low | 2.7 | a | 0.66 | a | 6.4 | a | 0.10 | b |
| moderate | 3.8 | b | 0.73 | b | 8.7 | b | 0.09 | ab | |
| high | 4.6 | c | 0.75 | b | 10.8 | c | 0.08 | a | |
| Cultivars | Treatments | Ca Concentrations (mg-Ca/kg-DW) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Whole Leaves | Upper Stems | Lower Stems | Roots | ||||||
| AK | low | 1.6 | a | 0.52 | a | 0.60 | a | 6.79 | a |
| moderate | 2.0 | b | 0.87 | b | 0.93 | b | 9.44 | b | |
| high | 2.2 | c | 0.96 | b | 1.11 | b | 9.81 | b | |
| CW | low | 1.6 | a | 0.80 | a | 0.95 | a | 7.69 | a |
| moderate | 2.1 | b | 1.58 | b | 1.10 | b | 9.89 | b | |
| high | 2.6 | c | 2.24 | c | 1.45 | b | 11.09 | c | |
| VY | low | 1.0 | a | 0.75 | a | 0.65 | a | 7.02 | a |
| moderate | 2.2 | b | 1.19 | b | 1.00 | b | 7.51 | a | |
| high | 2.7 | c | 1.70 | c | 1.13 | b | 9.35 | b | |
| Cultivars | Treatments | Ca Concentrations (mg-Ca/kg-DW) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Upper (1st) Leaves | Middle (4th) Leaves | Lower (7th) Leaves | |||||||||||
| Tips | Bases | Tips | Bases | Tips | Bases | ||||||||
| AK | low | 1.77 | a | 1.29 | a | 1.33 | a | 1.09 | a | 1.92 | a | 1.08 | a |
| moderate | 2.50 | b | 1.73 | b | 2.01 | b | 1.88 | b | 2.28 | b | 1.57 | b | |
| high | 3.28 | b | 1.88 | b | 2.39 | c | 2.13 | c | 2.42 | b | 1.74 | b | |
| CW | low | 2.15 | n.s. | 1.55 | a | 1.79 | n.s. | 1.77 | a | 1.63 | a | 1.43 | a |
| moderate | 1.85 | 2.31 | b | 1.78 | 2.68 | b | 1.77 | ab | 2.01 | b | |||
| high | 1.85 | 3.09 | c | 1.86 | 3.44 | c | 2.12 | b | 2.71 | c | |||
| VY | low | 1.52 | a | 1.36 | a | 1.75 | a | 1.51 | a | 1.81 | a | 1.37 | a |
| moderate | 3.52 | b | 1.90 | ab | 2.27 | b | 2.23 | b | 2.23 | b | 2.16 | b | |
| high | 3.36 | b | 2.72 | b | 2.45 | b | 2.69 | c | 2.06 | b | 2.39 | b | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuronuma, T.; Ando, M.; Watanabe, H. Tipburn Incidence and Ca Acquisition and Distribution in Lisianthus (Eustoma grandiflorum (Raf.) Shinn.) Cultivars under Different Ca Concentrations in Nutrient Solution. Agronomy 2020, 10, 216. https://doi.org/10.3390/agronomy10020216
Kuronuma T, Ando M, Watanabe H. Tipburn Incidence and Ca Acquisition and Distribution in Lisianthus (Eustoma grandiflorum (Raf.) Shinn.) Cultivars under Different Ca Concentrations in Nutrient Solution. Agronomy. 2020; 10(2):216. https://doi.org/10.3390/agronomy10020216
Chicago/Turabian StyleKuronuma, Takanori, Masaya Ando, and Hitoshi Watanabe. 2020. "Tipburn Incidence and Ca Acquisition and Distribution in Lisianthus (Eustoma grandiflorum (Raf.) Shinn.) Cultivars under Different Ca Concentrations in Nutrient Solution" Agronomy 10, no. 2: 216. https://doi.org/10.3390/agronomy10020216